
Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage


 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Cold Stress Treatment
2.2. Determination of Photosynthetic Pigment Contents
2.3. Determination of Osmolyte Contents
2.4. Determination of Oxidative Damage and Enzymatic Antioxidant Activities
2.5. Determination of Non-Enzymatic Antioxidant Compounds
3. Statistical Analysis
4. Results
4.1. Effect of Cold Stress on Photosynthetic Pigment Contents
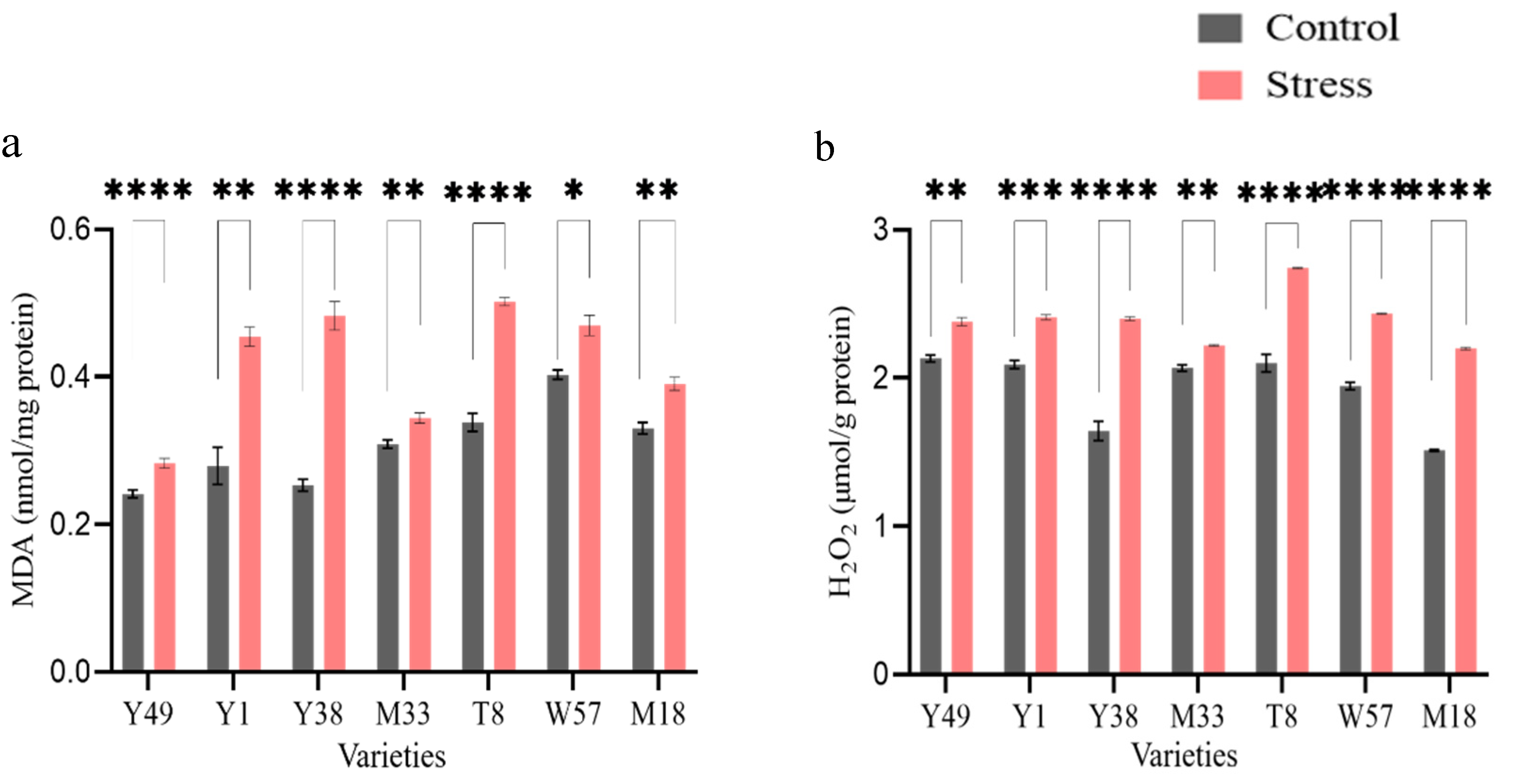
4.2. Oxidative Stress Evaluation
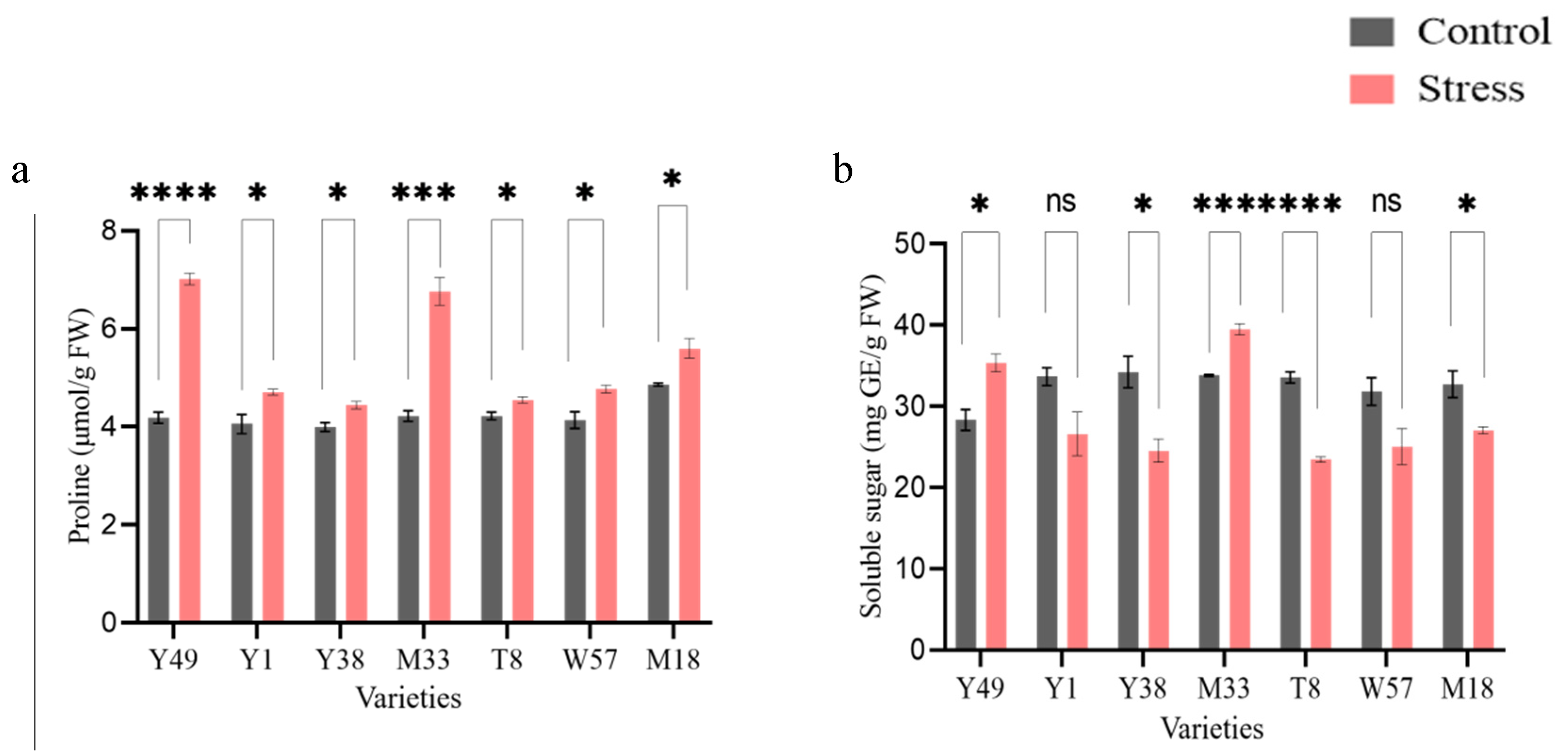
4.3. Effect of Cold Stress on Osmolytes Contents
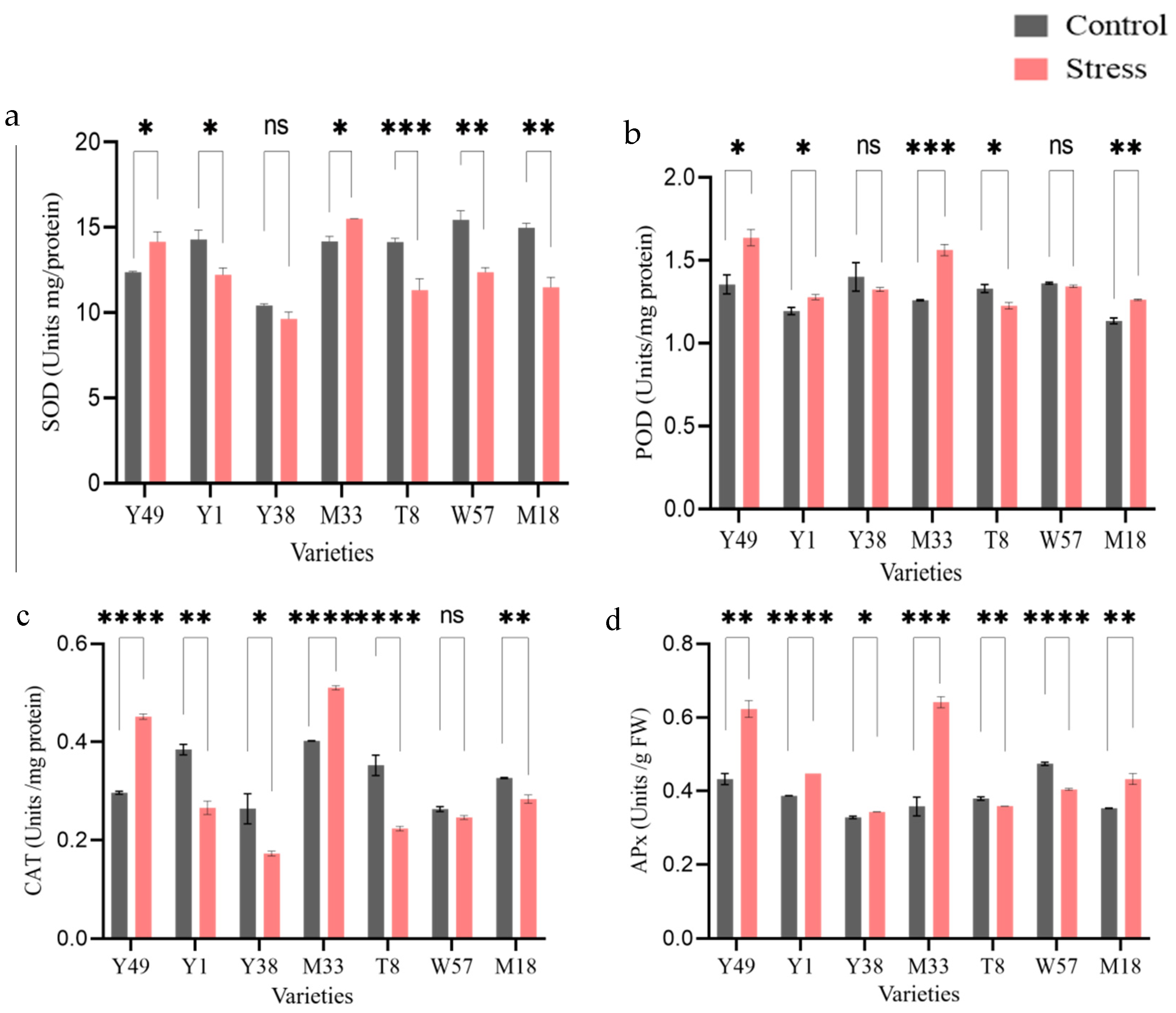
4.4. Enzymatic Antioxidant Activities (SOD, POD, CAT, and APx)
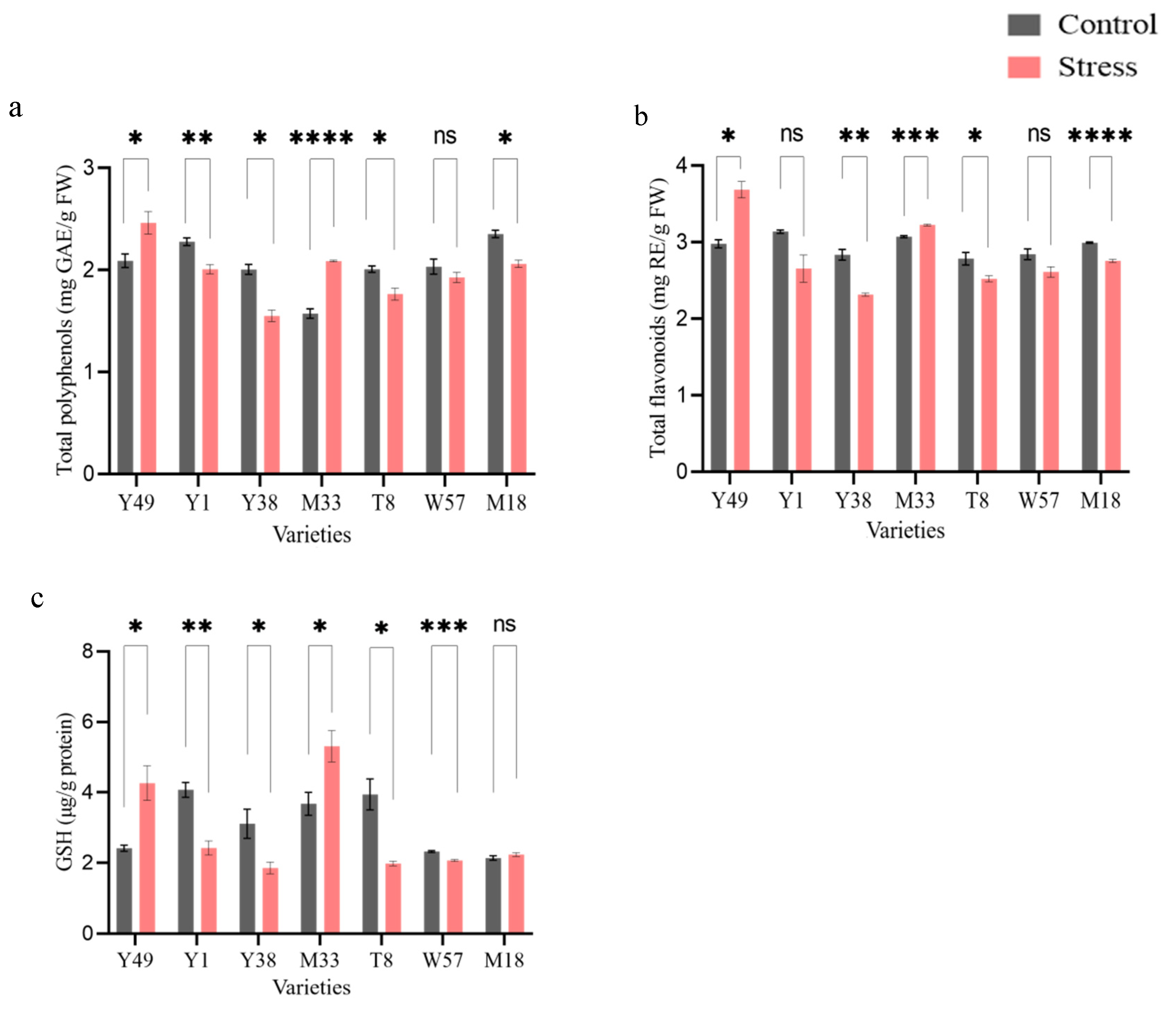
4.5. Non-Enzymatic Antioxidant Activities (TFC, TPC, and GSH)
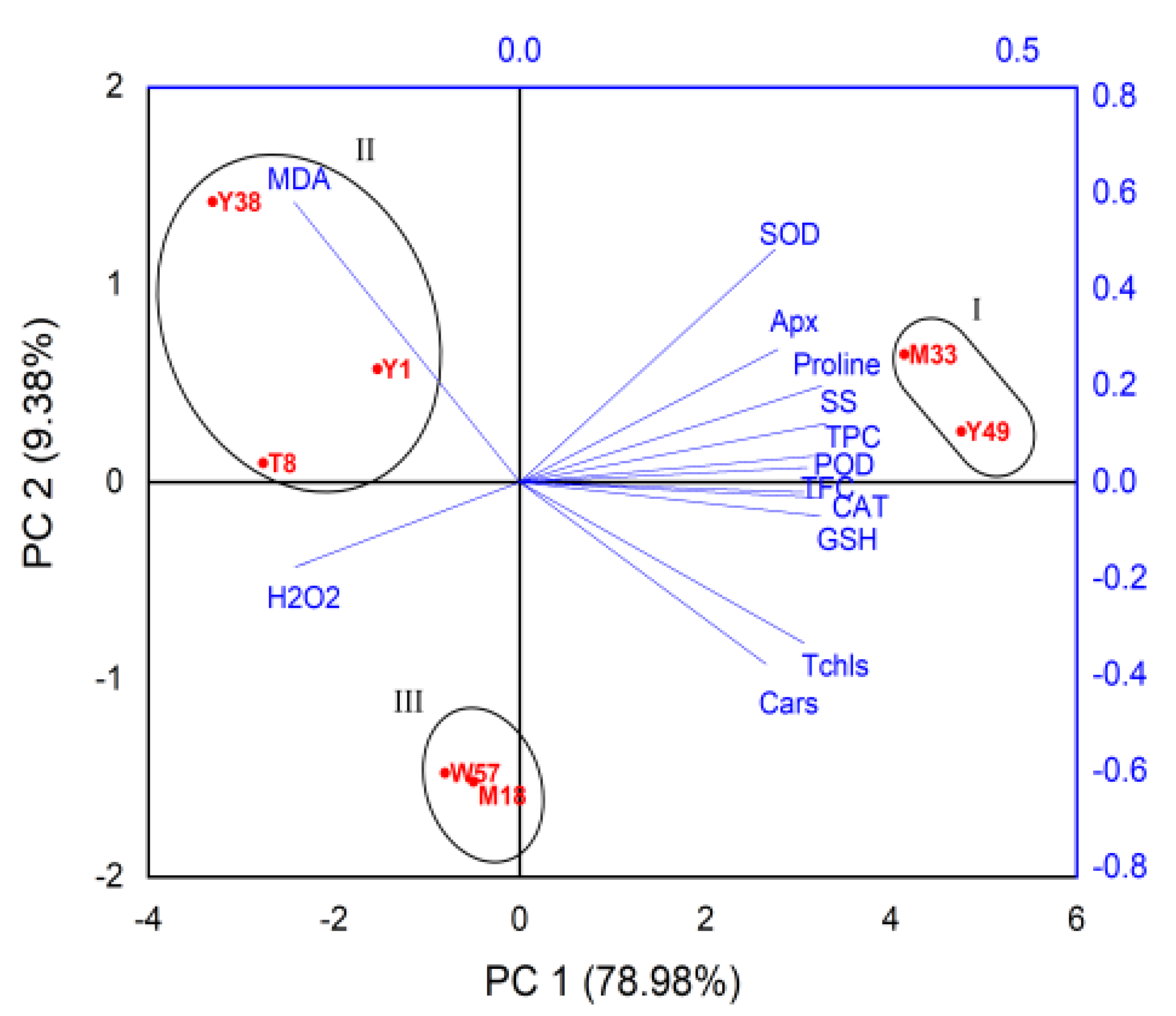
4.6. Principle Component Analysis (PCA)
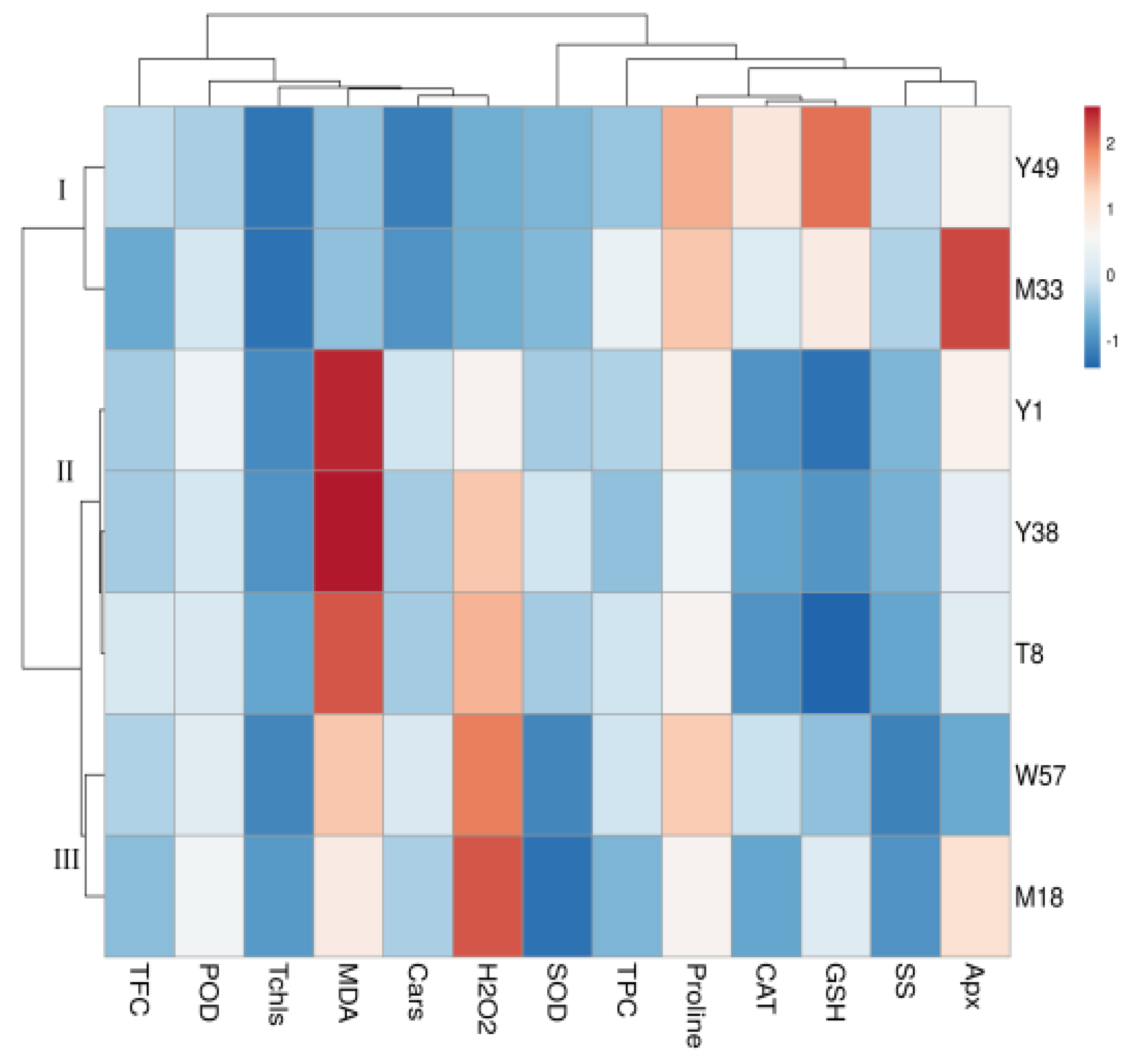
4.7. Cluster Heat Map Analysis
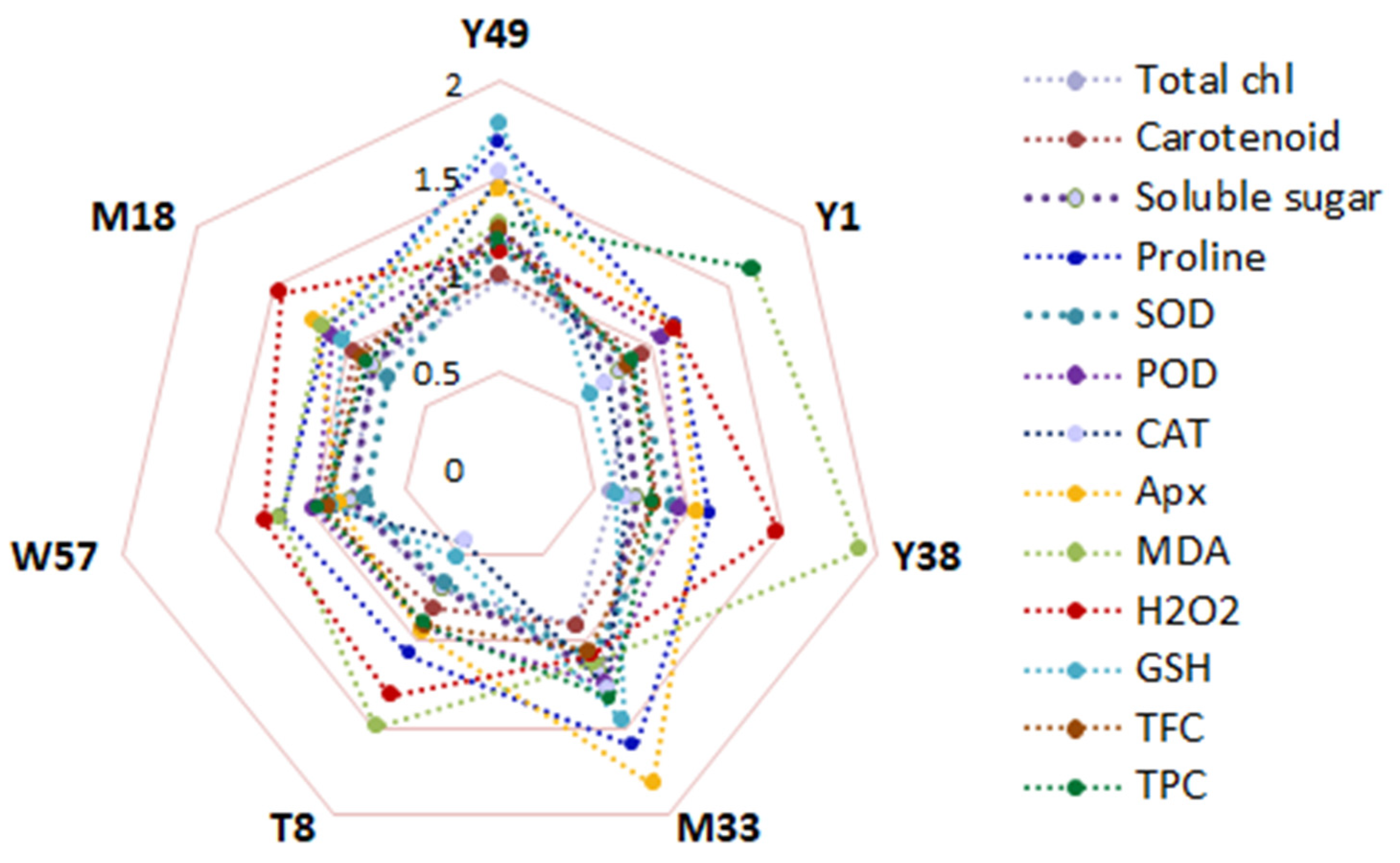
4.8. Genotypic Variation under Cold Stress
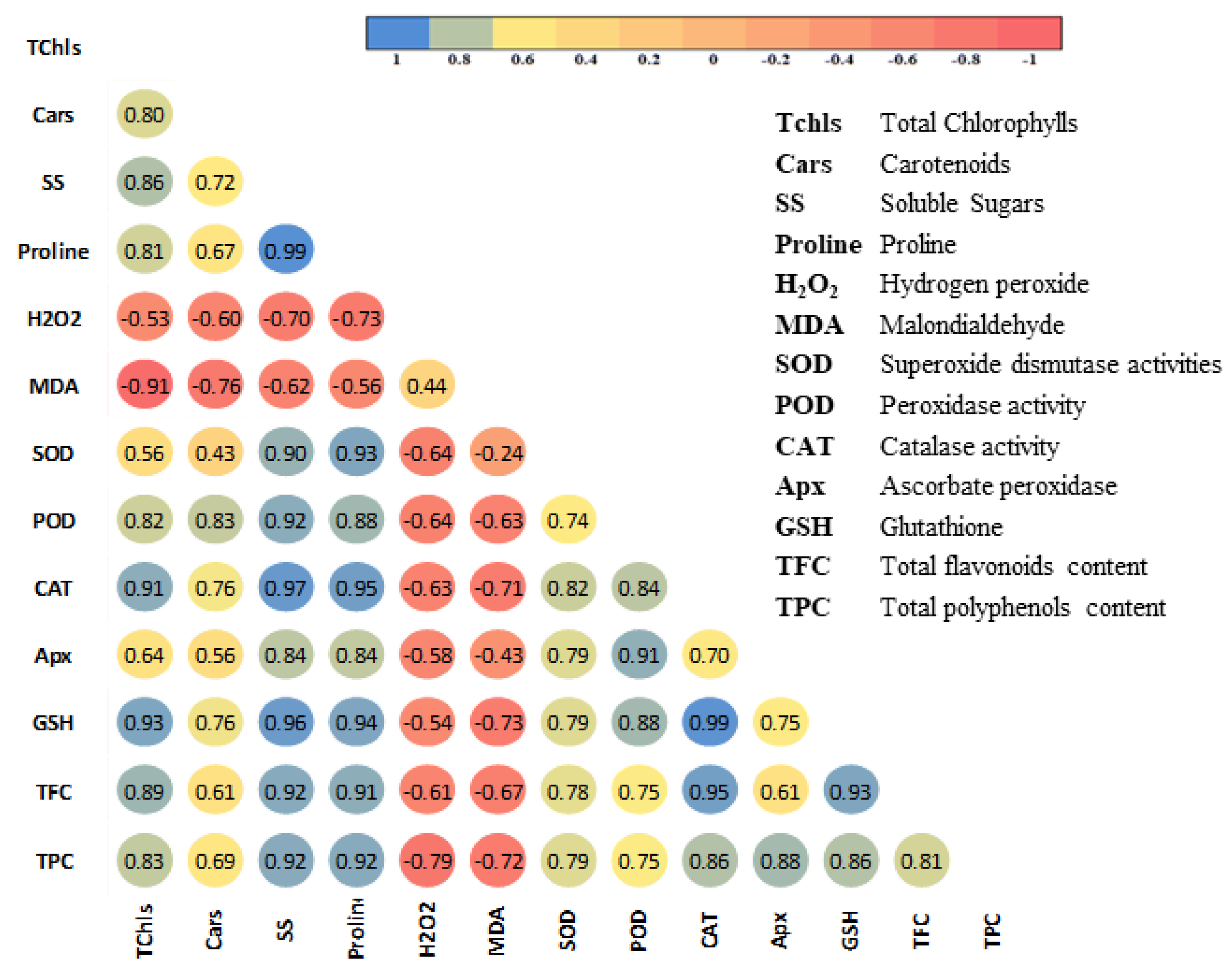
4.9. Correlation of Various Biochemical Parameters
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raza, A.; Su, W.; Hussain, M.A.; Mehmood, S.S.; Zhang, X.; Cheng, Y.; Zou, X.; Lv, Y. Integrated Analysis of Metabolome and Transcriptome Reveals Insights for Cold Tolerance in Rapeseed (Brassica napus L.). Front. Plant Sci. 2021, 12, 1796. [Google Scholar] [CrossRef]
- Szczerba, A.; Płażek, A.; Pastuszak, J.; Kopeć, P.; Hornyák, M.; Dubert, F. Effect of low temperature on germination, growth, and seed yield of four soybean (Glycine max L.) cultivars. Agronomy 2021, 11, 800. [Google Scholar] [CrossRef]
- Yang, Q.S.; Gao, J.; He, W.D.; Dou, T.X.; Ding, L.J.; Wu, J.H.; Li, C.Y.; Peng, X.X.; Zhang, S.; Yi, G.J. Comparative transcriptomics analysis reveals difference of key gene expression between banana and plantain in response to cold stress. BMC Genom. 2015, 16, 446. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, Q.; Hussain, S.; Mei, J.; Dong, H.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Pre-sowing Seed Treatments in Direct-seeded Early Rice: Consequences for Emergence, Seedling Growth and Associated Metabolic Events under Chilling Stress. Sci. Rep. 2016, 6, 19637. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R. A Review Report: Low Temperature Stress for Crop Production. Int. J. Pure Appl. Biosci. 2018, 6, 575–598. [Google Scholar] [CrossRef]
- Saleem, M.H.; Rehman, M.; Zahid, M.; Imran, M.; Xiang, W.; Liu, L. Morphological changes and antioxidative capacity of jute (Corchorus capsularis, Malvaceae) under different color light-emitting diodes. Rev. Bras. Bot. 2019, 42, 581–590. [Google Scholar] [CrossRef]
- Kumari, K.; Singh, P.K.; Kumari, S.; Singh, K.M. Dynamics of Jute Export in India. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 3769–3774. [Google Scholar] [CrossRef]
- Islam, M.M. Biochemistry, Medicinal and Food values of Jute (Corchorus capsularis L. and C. olitorius L.) leaf: A Review. Int. J. Enhanc. Res. Sci. Technol. Eng. 2013, 2, 35–44. [Google Scholar]
- Dansi, A.; Adjatin, A.; Adoukonou-Sagbadja, H.; Faladé, V.; Yedomonhan, H.; Odou, D.; Dossou, B. Traditional leafy vegetables and their use in the Benin Republic. Genet. Resour. Crop Evol. 2008, 55, 1239–1256. [Google Scholar] [CrossRef]
- Zeghichi, S.; Kallithraka, S.; Simopoulos, A.P. Nutritional composition of molokhia (Corchorus olitorius) and stamnagathi (Cichorium spinosum). World Rev. Nutr. Diet. 2003, 91, 1–21. [Google Scholar] [CrossRef]
- Yang, Z.; Lu, R.; Dai, Z.; Yan, A.; Tang, Q.; Cheng, C.; Xu, Y.; Yang, W.; Su, J. Salt-stress response mechanisms using de novo transcriptome sequencing of salt-tolerant and sensitive Corchorus spp. Genotypes. Genes 2017, 8, 226. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.; Ali, S. Agronomic Research Advances in Jute Crops of Bangladesh. AASCIT J. Biol. 2018, 3, 34–46. [Google Scholar]
- Hossain, M.B.; Awal, A.; Rahman, M.A.; Haque, S.; Khan, H. Distinction between Cold-sensitive and -tolerant Jute by DNA Polymorphisms. J. Biochem. Mol. Biol. 2003, 36, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Cui, J.M.; Li, G.X.; Yuan, M.; Zhang, Z.W.; Yuan, S.; Zhang, H.Y. Effect of salicylic acid on the antioxidant system and photosystem II in wheat seedlings. Biol. Plant. 2016, 60, 139–147. [Google Scholar] [CrossRef]
- Rentel, M.C.; Knight, M.R.; Kingdom, U. Oxidative Stress-Induced Calcium Signaling. Plant Physiol. 2004, 135, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Erdal, S.; Genisel, M.; Turk, H.; Dumlupinar, R.; Demir, Y. Modulation of alternative oxidase to enhance tolerance against cold stress of chickpea by chemical treatments. J. Plant Physiol. 2015, 175, 95–101. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. Water-Culture Method Growing Plants without Soil; California Agricultural Experiment Station: Berkely, CA, USA, 1950; Volume 347, pp. 29–31. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 207, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A. The estimaion of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 197. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Biswas, A.; Dey, S.; Li, D.; Liu, Y.; Zhang, J.; Huang, S.; Pan, G.; Deng, Y. Comparison of Phytochemical Profile, Mineral Content, and in Vitro Antioxidant Activities of Corchorus capsularis and Corchorus olitorius Leaf Extracts from Different Populations. J. Food Qual. 2020, 2020, 2931097. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Chang, S.K.C. Effect of soaking, boiling, and steaming on total phenolic contentand antioxidant activities of cool season food legumes. Food Chem. 2008, 110, 1–13. [Google Scholar] [CrossRef]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Physiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Zainol, M.K.; Abd-Hamid, A.; Yusof, S.; Muse, R. Antioxidative activity and total phenolic compounds of leaf, root and petiole of four accessions of Centella asiatica (L.) Urban. Food Chem. 2003, 81, 575–581. [Google Scholar] [CrossRef]
- Xu, S.C.; Li, Y.P.; Hu, J.; Guan, Y.J.; Ma, W.G.; Zheng, Y.Y.; Zhu, S.J. Responses of antioxidant enzymes to chilling stress in tobacco seedlings. Agric. Sci. China 2010, 9, 1594–1601. [Google Scholar] [CrossRef]
- Wu, J.; Lightner, J.; Warwick, N.; Browse, J. Low-temperature damage and subsequent recovery of fab1 mutant arabidopsis exposed to 2 °C. Plant Physiol. 1997, 113, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Noreen, Z.; Ashraf, M. Changes in antioxidant enzymes and some key metabolites in some genetically diverse cultivars of radish (Raphanus sativus L.). Environ. Exp. Bot. 2009, 67, 395–402. [Google Scholar] [CrossRef]
- Atici, Ö.; Demir, Y.; Kocaçalişkan, İ. Effects of low temperature on winter wheat and cabbage leaves. Biol. Plant. 2003, 46, 603–606. [Google Scholar] [CrossRef]
- Tewari, A.K.; Tripathy, B.C. Temperature-stress-induced impairment of chlorophyll biosynthetic reactions in cucumber and wheat. Plant Physiol. 1998, 117, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B.; Gilmore, A.M.; Iii, W.W.A. In vivo functions of carotenoids in higher plants. FASEB J. 1996, 10, 403–412. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytol. 2006, 172, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Boese, S.R.; Huner, N.P.A. Effect of growth temperature and temperature shifts on spinach leaf morphology and photosynthesis. Plant Physiol. 1990, 94, 1830–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Asthir, B.J.B.P. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Huang, M.; Guo, Z. Responses of antioxidative system to chilling stress in two rice cultivars differing in sensitivity. Biol. Plant. 2005, 49, 81–84. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Huang, X.; Chen, M.H.; Yang, L.T.; Li, Y.R.; Wu, J.M. Effects of Exogenous Abscisic Acid on Cell Membrane and Endogenous Hormone Contents in Leaves of Sugarcane Seedlings under Cold Stress. Sugar Tech 2015, 17, 59–64. [Google Scholar] [CrossRef]
- Lu, K.; Sun, J.; Li, Q.; Li, X.; Jin, S. Effect of cold stress on growth, physiological characteristics, and calvin-cycle-related gene expression of grafted watermelon seedlings of different gourd rootstocks. Horticulturae 2021, 7, 391. [Google Scholar] [CrossRef]
- Yuanyuan, M.; Yali, Z.; Jiang, L.; Hongbo, S. Roles of plant soluble sugars and their responses to plant cold stress. Afr. J. Biotechnol. 2009, 8, 2004–2010. [Google Scholar] [CrossRef]
- Uemura, M.; Steponkus, P.L. A contrast of the plasma membrane lipid composition of oat and rye leaves in relation to freezing tolerance. Plant Physiol. 1994, 104, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Freitas, G.M.; Thomas, J.; Liyanage, R.; Lay, J.O.; Basu, S.; Ramegowda, V.; do Amaral, M.N.; Benitez, L.C.; Braga, E.J.B.; Pereira, A. Cold tolerance response mechanisms revealed through comparative analysis of gene and protein expression in multiple rice genotypes. PLoS ONE 2019, 14, e0218019. [Google Scholar] [CrossRef]
- Morsy, M.R.; Almutairi, A.M.; Gibbons, J.; Yun, S.J.; De Los Reyes, B.G. The OsLti6 genes encoding low-molecular-weight membrane proteins are differentially expressed in rice cultivars with contrasting sensitivity to low temperature. Gene 2005, 344, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Fahimirad, S.; Karimzadeh, G.; Ghanati, F. Cold-induced Changes of Antioxidant Enzymes Activity and Lipid Peroxidation in Two Canola (Brassica napus L.) Cultivars. J. Plant Physiol. Breed. 2013, 3, 1–11. [Google Scholar]
- Khalofah, A.; Migdadi, H.; El-harty, E. Antioxidant enzymatic activities and growth response of quinoa (Chenopodium quinoa willd) to exogenous selenium application. Plants 2021, 10, 719. [Google Scholar] [CrossRef]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-induced reactive oxygen species accumulation by altering the enzymatic antioxidant system and associated cytotoxic, genotoxic, ultrastructural, and photosynthetic changes in plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posmyk, M.M.; Kontek, R.; Janas, K.M. Antioxidant enzymes activity and phenolic compounds content in red cabbage seedlings exposed to copper stress. Ecotoxicol. Environ. Saf. 2009, 72, 596–602. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Skyba, M.; Petijová, L.; Košuth, J.; Koleva, D.P.; Ganeva, T.G.; Kapchina-Toteva, V.M.; Čellárová, E. Oxidative stress and antioxidant response in Hypericum perforatum L. plants subjected to low temperature treatment. J. Plant Physiol. 2012, 169, 955–964. [Google Scholar] [CrossRef]
- Auh, C.K.; Scandalios, J.G. Spatial and temporal responses of the maize catalases to low temperature. Physiol. Plant. 1997, 101, 149–156. [Google Scholar] [CrossRef]
- Sevengor, S.; Yasar, F.; Kusvuran, S.; Ellialtioglu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.Z.; Chow, W.S.; Sun, L.L.; Chen, J.W.; Chen, Y.J.; Peng, C.L. The influence of low temperature on photosynthesis and antioxidant enzymes in sensitive banana and tolerant plantain (Musa sp.) cultivars. Photosynthetica 2011, 49, 201–208. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Tomaro, M.L.; Batlle, A.; Noriega, G.O. The role of 5-aminolevulinic acid in the response to cold stress in soybean plants. Phytochemistry 2010, 71, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Krasnovsky, A.A., Jr. Singlet molecular oxygen in photobiochemical systems: IR phosphorescence studies. Membr. Cell Biol. 1998, 12, 665–690. [Google Scholar]
- Tatari, M.; Fotouhi Ghazvini, R.; Mousavi, A.; Babaei, G. Comparison of some physiological aspects of drought stress resistance in two ground cover genus. J. Plant Nutr. 2018, 41, 1215–1226. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Almela, L.; Fernández-López, J.A.; Roca, M.J. High-performance liquid chromatographic screening of chlorophyll derivatives produced during fruit storage. J. Chromatogr. A 2000, 870, 483–489. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Sarikamiş, G.; Çakir, A. Influence of low temperature on aliphatic and indole glucosinolates in broccoli (Brassica oleracea var. italica L.). Acta Hortic. 2016, 1145, 79–84. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.S.; Wang, L. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Weidner, S.; Karolak, M.; Karamać, M.; Kosińska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots (Vitis vinifera L.) under drought stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Swigonska, S.; Amarowicz, R.; Król, A.; Mostek, A.; Badowiec, A.; Weidner, S. Influence of abiotic stress during soybean germination followed by recovery on the phenolic compounds of radicles and their antioxidant capacity. Acta Soc. Bot. Pol. 2014, 83, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Hakam, N.; Simon, J.P. Effect of low temperatures on the activity of oxygen-scavenging enzymes in two populations of the C4 grass Echinochloa crus-galli. Physiol. Plant. 1996, 97, 209–216. [Google Scholar] [CrossRef]
- Gechev, T.; Willekens, H.; Van Montagu, M.; Inzé, D.; Van Camp, W.; Toneva, V.; Minkov, I. Different responses of tobacco antioxidant enzymes to light and chilling stress. J. Plant Physiol. 2003, 160, 509–515. [Google Scholar] [CrossRef]
- Rajametov, S.N.; Lee, K.; Jeong, H.B.; Cho, M.C.; Nam, C.W.; Yang, E.Y. Physiological traits of thirty-five tomato accessions in response to low temperature. Agriculture 2021, 11, 792. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Variety | Origin (Province/Country) | |
|---|---|---|---|
| Y49 | C. capsularis | Huangma 971 | Hunan |
| Y38 | C. capsularis | Miandianyuanguo | Myanmar |
| Y1 | C. capsularis | Longxihongpi | Longxi county, Guangdong |
| T8 | C. olitorius | T8 | Zhejiang |
| W57 | C. olitorius | W57 | Zhejiang |
| M33 | C. olitorius | Funong 5 | Fujian |
| M18 | C. olitorius | Maliyengshengchangguo | Mali |
| Chlorophyll | Carotenoid | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | Chl a | Chl b | Total Chl | ||||||||
| Control | Treatment | CSI% | Control | Treatment | CSI% | Control | Treatment | CSI% | Control | Treatment | |
| Y49 | 1.77 ± 0.01 a | 1.73 ± 0.0 a | 97.81 | 0.61± 0.0 a | 0.60 ± 0.01 b | 98.42 | 2.39 ± 0.0 a | 2.34 ± 0.0 b | 97.97 | 0.127 ± 0.0 b | 0.129 ± 0.0 a |
| Y1 | 1.93 ± 0.02 a | 1.23 ± 0.01 b | 64.14 | 0.50 ± 0.04 a | 0.77 ± 0.02 b | 152.87 | 2.43 ± 0.01 a | 2.00 ± 0.03 b | 82.44 | 0.13 ± 0.0 a | 0.12 ± 0.0 b |
| Y38 | 1.94 ± 0.0 a | 1.10 ± 0.02 b | 56.85 | 0.44 ± 0.02 a | 0.31 ± 0.02 b | 69.18 | 2.39 ± 0.01 a | 1.41 ± 0.01 b | 59.17 | 0.12 ± 0.01 a | 0.10 ± 0.0 b |
| M33 | 1.82 ± 0.0 b | 1.85 ± 0.0 a | 101.67 | 0.55 ± 0.04 a | 0.31 ± 0.01 b | 55.62 | 2.37 ± 0.0 a | 2.16 ± 0.02 b | 90.97 | 0.12 ± 0.0 a | 0.10 ± 0.0 b |
| T8 | 1.75 ± 0.03 a | 1.10 ± 0.02 b | 63.23 | 0.46± 0.0 a | 0.44 ± 0.0 b | 94.93 | 2.20 ± 0.01 a | 1.54 ± 0.02 b | 69.88 | 0.11 ± 0.0 a | 0. 09 ± 0.0 b |
| W57 | 1.51 ± 0.0 a | 1.21 ± 0.0 b | 80.43 | 0.32 ± 0.0 a | 0.25 ± 0.0 b | 78.95 | 1.83 ± 0.01 a | 1.46 ± 0.0 b | 80.17 | 0.10 ± 0.0 a | 0.09 ± 0.0 a |
| M18 | 1.61 ± 0.02 a | 1.24 ± 0.03 b | 77.03 | 0.53 ± 0.0 a | 0.48 ± 0.01 a | 110.51 | 2.09 ± 0.0 a | 1.77 ± 0.03 b | 84.71 | 0.109 ± 0.0 a | 0.105 ± 0.0 b |
| Genotypes | Total chl | Carotenoid | Soluble Sugar | Proline | SOD | POD | CAT | APx | MDA | H2O2 | GSH | TFC | TPC | Mean STI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y49 | 0.98 ± 0.0 a | 1.00 ± 0.01 a | 1.25 ± 0.04 a | 1.68 ± 0.06 a | 1.14 ± 0.04 a | 1.21 ± 0.07 a | 1.52 ± 0.01 a | 1.44 ± 0.07 b | 1.17 ± 0.02 c | 1.12 ± 0.01 c | 1.79 ± 0.27 a | 1.24 ± 0.05 a | 1.18 ± 0.03 b | 1.29 |
| Y1 | 0.66 ± 0.01 c | 0.94 ± 0.01 bc | 0.79 ± 0.08 b | 1.17 ± 0.06 b | 0.86 ± 0.0 bc | 1.07 ± 0.03 bc | 0.69 ± 0.05 d | 1.16 ± 0.0 cd | 1.66 ± 0.17 ab | 1.15 ± 0.02 c | 0.59 ± 0.03 cd | 0.85.05 c | 0.88 ± 0.02 cd | 0.96 |
| Y38 | 0.59 ± 0.0 e | 0.82 ± 0.02 d | 0.72 ± 0.05 b | 1.11 ± 0.04 b | 0.92 ± 0.03 b | 0.95 ± 0.06 cd | 0.67 ± 0.05 d | 1.05 ± 0.0 cde | 1.91 ± 0.10 a | 1.47 ± 0.06 a | 0.61 ± 0.06 cd | 0.82 ± 0.02 c | 0.77 ± 0.02 d | 0.96 |
| M33 | 0.91 ± 0.03 b | 0.99 ± 0.01 a | 1.17 ± 0.02 a | 1.60 ± 0.02 a | 1.10 ± 0.02 a | 1.24 ± 0.02 a | 1.27 ± 0.01 b | 1.82 ± 0.17 a | 1.12 ± 0.01 c | 1.07 ± 0.01 c | 1.46 ± 0.11 a | 1.05 ± 0.0 b | 1.33 ± 0.04 a | 1.24 |
| T8 | 0.70 ± 0.01 d | 0.80 ± 0.0 d | 0.70 ± 0.0 b | 1.08 ± 0.03 b | 0.80 ± 0.05 c | 0.92 ± 0.03 d | 0.64 ± 0.0 d | 0.95 ± 0.01 de | 1.49 ± 0.06 b | 1.31 ± 0.03 b | 0.52 ± 0.08 d | 0.91 ± 0.03 c | 0.88 ± 0.04 cd | 0.90 |
| W57 | 0.80 ± 0.0 c | 0.9 ± 0.01 ab | 0.79 ± 0.05 b | 1.16 ± 0.06 b | 0.80 ± 0.02 c | 0.99 ± 0.01 bcd | 0.94 ± 0.02 c | 0.85 ± 0.0 e | 1.17 ± 0.05 c | 1.25 ± 0.01 b | 0.89 ± 0.02 bc | 0.92 ± 0.03 c | 0.95 ± 0.05 c | 0.96 |
| M18 | 0.85 ± 0.01 c | 0.96 ± 0.01 b | 0.83 ± 0.04 b | 1.15 ± 0.03 b | 0.77 ± 0.05 c | 1.11 ± 0.01 ab | 0.87 ± 0.02 c | 1.23 ± 0.03 bc | 1.18 ± 0.03 c | 1.45 ± 0.0 a | 1.05 ± 0.05 b | 0.92 ± 0 c | 0.90 ± 0.01 c | 1.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dey, S.; Biswas, A.; Huang, S.; Li, D.; Liu, L.; Deng, Y.; Xiao, A.; Birhanie, Z.M.; Zhang, J.; Li, J.; et al. Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage. Agronomy 2021, 11, 2547. https://doi.org/10.3390/agronomy11122547
Dey S, Biswas A, Huang S, Li D, Liu L, Deng Y, Xiao A, Birhanie ZM, Zhang J, Li J, et al. Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage. Agronomy. 2021; 11(12):2547. https://doi.org/10.3390/agronomy11122547
Chicago/Turabian StyleDey, Susmita, Ashok Biswas, Siqi Huang, Defang Li, Liangliang Liu, Yong Deng, Aiping Xiao, Ziggiju Mesenbet Birhanie, Jiangjiang Zhang, Jianjun Li, and et al. 2021. "Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage" Agronomy 11, no. 12: 2547. https://doi.org/10.3390/agronomy11122547
APA StyleDey, S., Biswas, A., Huang, S., Li, D., Liu, L., Deng, Y., Xiao, A., Birhanie, Z. M., Zhang, J., Li, J., & Gong, Y. (2021). Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage. Agronomy, 11(12), 2547. https://doi.org/10.3390/agronomy11122547