
Protection of Strawberry Plants against Charcoal Rot Disease (Macrophomina phaseolina) Induced by Azospirillum brasilense

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fungal Cultures
2.3. Inoculum
2.4. Biocontrol Related Enzymatic Activity of A. brasilense Strains
2.5. Antifungal Activity of A. brasilense Strains
2.6. Histological Determinations
2.6.1. Stomatal Closure
2.6.2. Callose Deposition and Lignification
2.7. Induction of Resistance against Charcoal Rot Disease
2.8. Statistical Analyses
3. Results
3.1. Biocontrol Enzymatic Activity
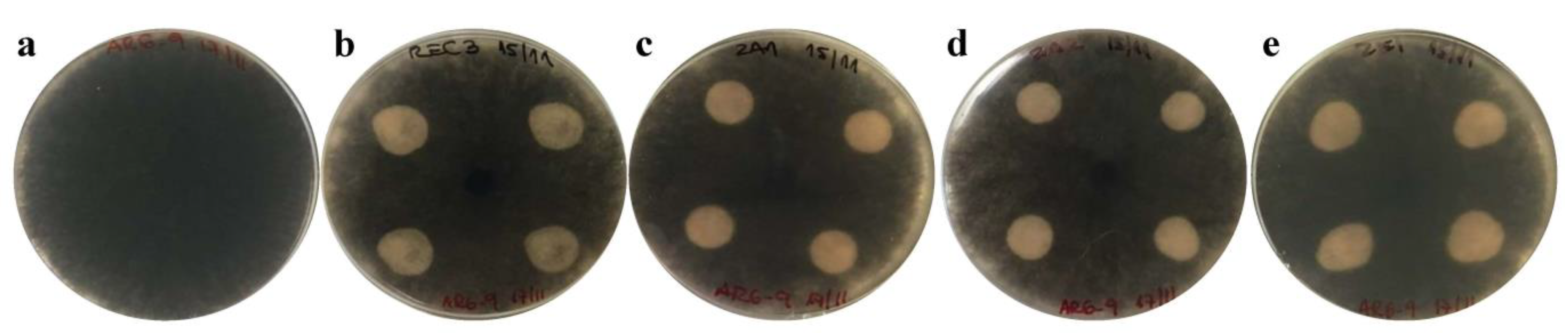
3.2. In Vitro Antifungal Activity
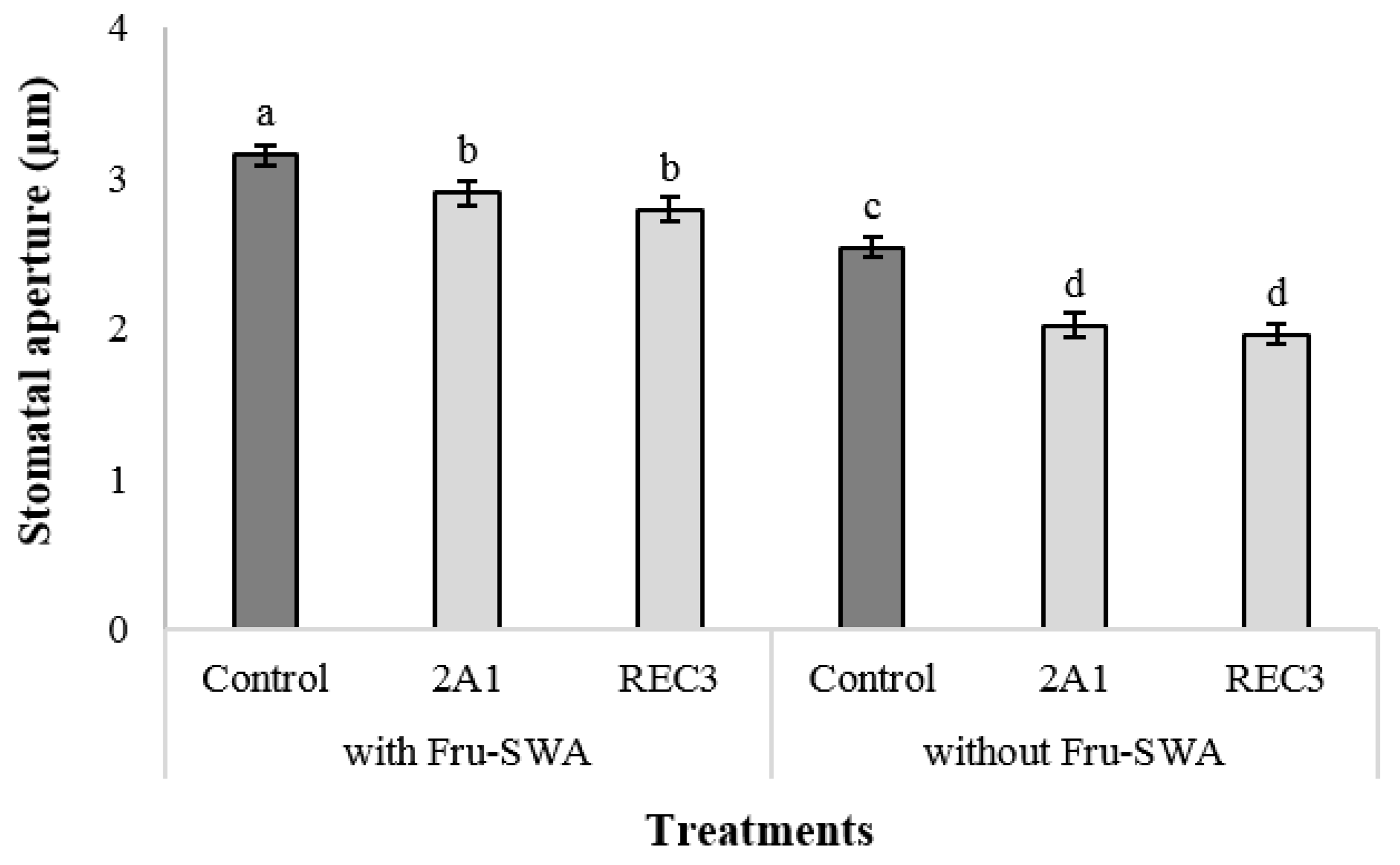
3.3. Stomatal Closure
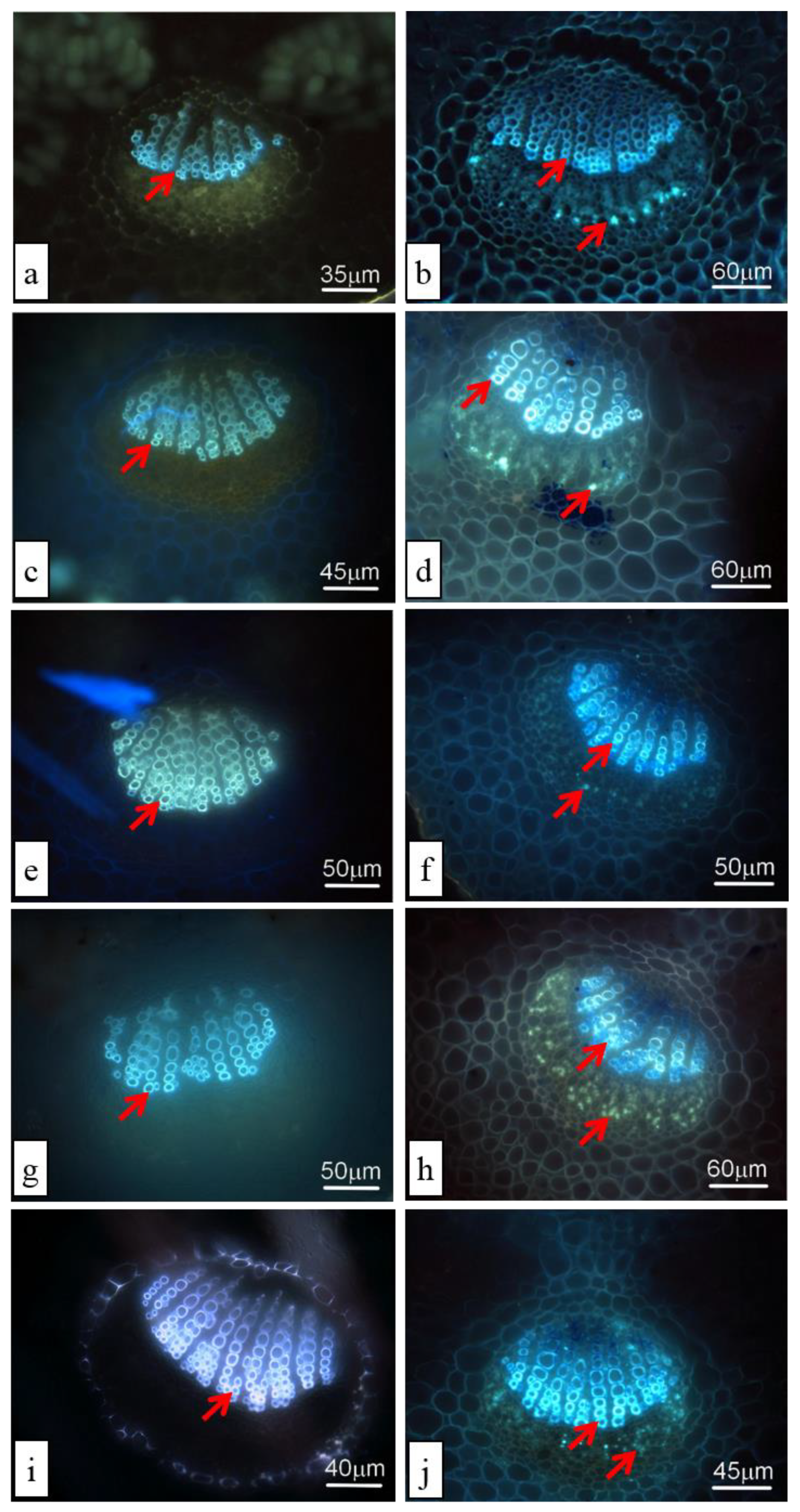
3.4. Lignin and Callose Accumulation
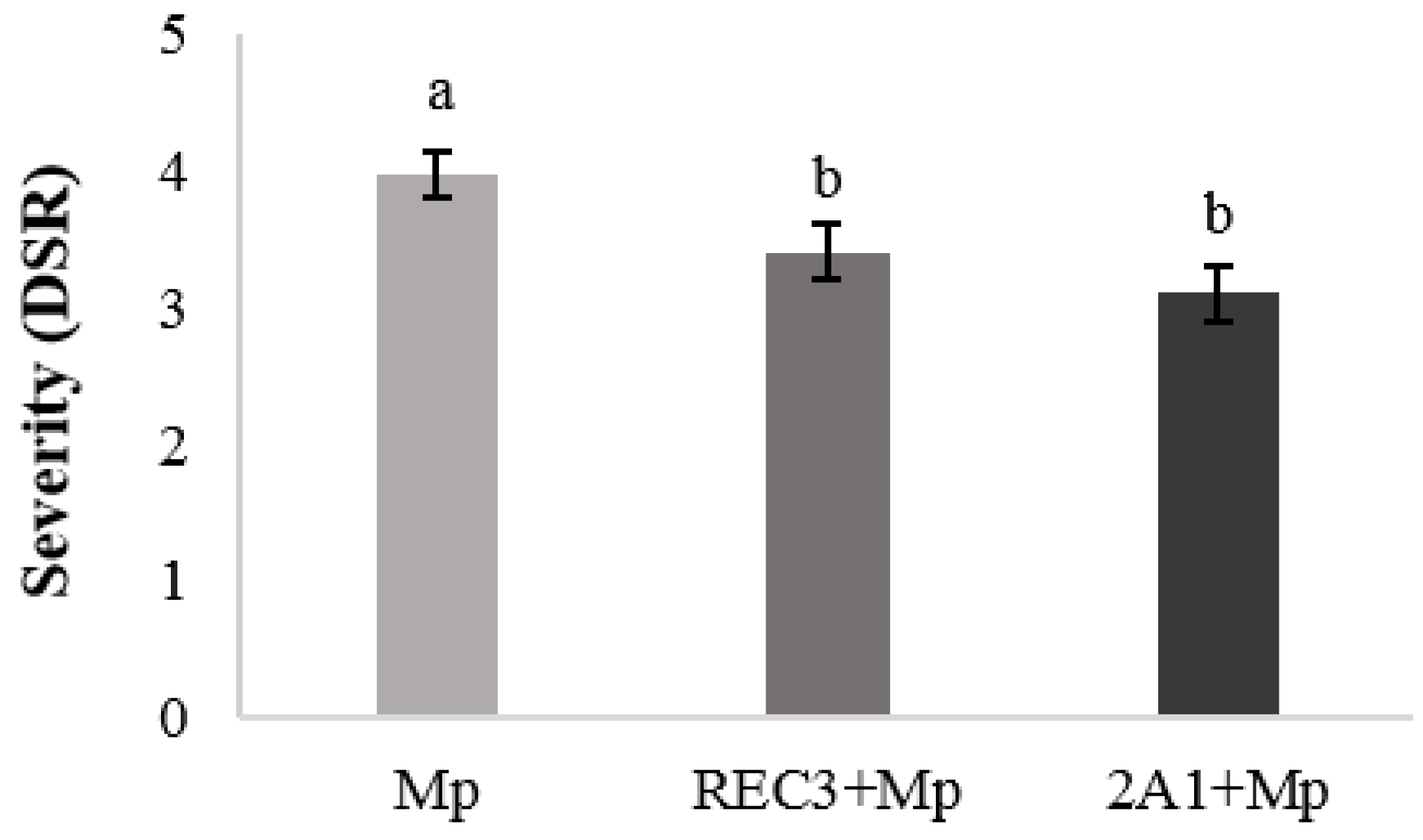
3.5. Induction of Resistance Against Charcoal Rot Disease
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ijaz, S.; Sadaqat, H.A.; Khan, M.N. A review of the impact of charcoal rot (Macrophomina phaseolina) on sunflower. J. Agric. Sci. 2012, 151, 222–227. [Google Scholar] [CrossRef]
- Koike, S.T. Crown rot of strawberry caused by Macrophomina phaseolina in California. Plant Dis. 2008, 92, 1253. [Google Scholar] [CrossRef] [PubMed]
- Baino, O.M.; Salazar, S.M.; Ramallo, A.C.; Kirschbaum, D.S. First report of Macrophomina phaseolina causing strawberry crown and root rot in Northwestern Argentina. Plant Dis. 2011, 95, 1477. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, D.S.; Vicente, C.E.; Cano-Torres, M.A.; Gambardella, M.; Veizaga-Pinto, F.K.; Antunes, L.E.C. Strawberry in South America: From the Caribbean to Patagonia. Acta Hortic. 2017, 1156, 947–956. [Google Scholar] [CrossRef]
- Camejo, D.; Guzmán-Cedeño, Á.; Moreno, A. Reactive oxygen species, essential molecules, during plant–pathogen interactions. Plant Physiol. Biochem. 2016, 103, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced systemic resistance (ISR) and Fe deficiency responses in dicot plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef]
- Nie, P.; Li, X.; Wang, S.; Guo, J.; Zhao, H.; Niu, D. Induced Systemic Resistance against Botrytis cinerea by Bacillus cereus AR156 through a JA/ET- and NPR1-Dependent Signaling Pathway and Activates PAMP-Triggered Immunity in Arabidopsis. Front. Plant Sci. 2017, 8, 238. [Google Scholar] [CrossRef]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Lodha, S.; Mawar, R. Population dynamics of Macrophomina phaseolina in relation to disease management: A review. J. Phytopathol. 2019, 168, 1–17. [Google Scholar] [CrossRef]
- El-Barougy, E.; Awad, N.M.; Turky, A.S.; Hamed, H.A. Antagonistic activity of selected strains of rhizobacteria against Macrophomina phaseolina of soybean plants. Am. J. Agric. Environ. Sci. 2009, 5, 337–347. [Google Scholar]
- Saba, I.; Khan, T.A. Management of disease complex of balsam caused by Meloidogyne javanica and Macrophomina phaseolina by using biofertilisers and pesticides. Arch. Phytopathol. Plant Prot. 2010, 43, 1381–1386. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Basallote-Ureba, M.J.; Aguado, A.; Khalid, A.K.D.I.; Capote, N. Biological control of strawberry soil-borne pathogens Macrophomina phaseolina and Fusarium solani, using Trichoderma asperellum and Bacillus spp. Phytopathol. Mediterr. 2016, 55, 109–120. [Google Scholar] [CrossRef]
- Tortora, M.L.; Díaz-Ricci, J.C.; Pedraza, R.O. Protection of strawberry plants (Fragaria ananassa Duch.) against anthracnose disease induced by Azospirillum brasilense. Plant Soil 2012, 356, 279–290. [Google Scholar] [CrossRef]
- Viejobueno, J.; Medina, J.J.; Aguado, A.; Camacho, M.; Camacho-Sánchez, M.; Salazar, S.M.; de los Santos, B. Caracterización morfológica, cultural y molecular de aislados de Macrophomina phaseolina de España y Argentina. In Proceedings of the Actas de Horticultura 83. IV Jornadas del Grupo Fresón y Otros Frutos Rojos, Valladolid, Spain, 21–22 May 2019; pp. 93–98. [Google Scholar]
- Pedraza, R.O.; Motok, J.; Tortora, M.L.; Salazar, S.M.; Díaz-Ricci, J.C. Natural occurrence of Azospirillum brasilense in strawberry plants. Plant Soil 2007, 295, 169–178. [Google Scholar] [CrossRef]
- Toffoli, L.M. Aplicación de bacterias promotoras del crecimiento vegetal en plantas ornamentales: Respuestas bioquímicas, estructurales y agronómicas de la interacción. PhD Thesis, Universidad Nacional de Tucumán, Tucumán, Argentina, 2019. [Google Scholar]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Teather, R.M.; Wood, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [CrossRef] [Green Version]
- Cowan, S.T.; Steel, K.J. Manual for the Identification of Medical Bacteria; Cambridge University Press: Cambridge, UK, 1965. [Google Scholar]
- Harley, J.P.; Prescott, L.M. Laboratory Exercises in Microbiology, 6th ed.; McGraw-Hill Science Engineering: New York, NY, USA, 2002. [Google Scholar]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Endophytic cultivable bacteria of the metal bioaccumulator Spartina maritima improve plant growth but not metal uptake in polluted marshes soils. Front. Microbiol. 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Reetha, A.K.; Pavani, S.L.; Mohan, S. Hydrogen cyanide production ability by bacterial antagonist and their antibiotics inhibition potential on Macrophomina phaseolina (Tassi.) Goid. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 172–178. [Google Scholar]
- Edmunds, L.K. Combined relation of plant maturity, temperature, and soil moisture to charcoal stalk rot development in grain sorghum. Phytopathology 1964, 54, 514–517. [Google Scholar]
- D’Ambrogio de Argüeso, A. Manual de Técnicas en Histología Vegetal; Hemisferio Sur. Buenos Aires: Buenos Aires, Argentina, 1986. [Google Scholar]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Comparisons of virulence of pathogens associated with crown and root diseases of strawberry in Western Australia with special reference to the effect of temperature. Sci. Hortic. 2011, 131, 39–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 19 January 2020).
- Karimi, E.; Sadeghi, A.; Peyman, A.D.; Dalvand, Y.; Omidvar, M.; Nezhad, M.K. Biocontrol science and technology biocontrol activity of salt tolerant Streptomyces isolates against phytopathogens causing root rot of sugar beet. Biocontrol Sci. Technol. 2012, 22, 333–349. [Google Scholar] [CrossRef]
- Abed, H.; Rouag, N.; Mouatassem, D.; Rouabhi, A. Screening for Pseudomonas and Bacillus antagonistic rhizobacteria strains for the biocontrol of Fusarium wilt of chickpea. Eurasian J. Soil Sci. 2016, 5, 182. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, F.; Alameda, M.; Schroder, E.C. In vitro evaluation of bacteria for the biological control of Macrophomina phaseolina. World J. Microbiol. Biotechnol. 1995, 11, 183–185. [Google Scholar] [CrossRef]
- Russo, A.; Vettori, L.; Felici, C.; Fiaschi, G.; Morini, S.; Toffanin, A. Enhanced micropropagation response and biocontrol effect of Azospirillum brasilense Sp245 on Prunus cerasifera L. clone Mr.S 2/5 plants. J. Biotechnol. 2008, 134, 312–319. [Google Scholar] [CrossRef]
- Cohen, A.; Bottini, R.; Pontin, M.; Berli, F.J.; Moreno, D.; Boccanlandro, H.; Travaglia, C.N.; Piccoli, P.N. Azospirillum brasilense ameliorates the response of Arabidopsis thaliana to drought mainly via enhancement of ABA levels. Physiol. Plant. 2015, 153, 79–90. [Google Scholar] [CrossRef]
- Bulegon, L.G.; Battistus, A.G.; Guimarães, V.F.; Inagaki, A.M.; Offemann, L.C.; Pomini de Souza, A.K.; Rabello de Oliveira, P.S. Physiological responses of Urochloa ruziziensis inoculated with Azospirillum brasilense to severe drought and rehydration conditions. Aust. J. Crop Sci. 2017, 11, 1283–1289. [Google Scholar] [CrossRef]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Nitric oxide as a secondary messenger during stomatal closure as a part of plant immunity response against pathogens. Nitric Oxide 2014, 43, 89–96. [Google Scholar] [CrossRef]
- Dehgahi, R.; Subramaniam, S.; Zakaria, L.; Joniyas, A.; Firouzjahi, F.B.; Haghnama, K.; Razinataj, M. Review of research on fungal pathogen attack and plant defense mechanism against pathogen. Int. J. Sci. Res. Agric. Sci. 2016, 2, 197–208. [Google Scholar] [CrossRef]
- Ferreira, R.B.; Monteiro, S.; Freitas, R.; Santos, C.N.; Chen, Z.; Batista, L.M.; Duarte, J.; Borges, A.; Teixeira, A.R. The role of plant defense proteins in fungal pathogenesis. Mol. Plant Pathol. 2007, 8, 677–700. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Molina, M.F.; Lovaisa, N.C.; Salazar, S.M.; Martínez-Zamora, M.G.; Díaz-Ricci, J.C.; Pedraza, R.O. Physiological, structural and molecular traits activated in strawberry plants after inoculation with the plant growth-promoting bacterium Azospirillum brasilense REC3. Plant Biol. 2014, 17, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, N. Elicitor-induced plant defence pathways. Trends Plant Sci. 1996, 1, 233–240. [Google Scholar] [CrossRef]
- Kombrink, E.; Somssich, I.E. Defense responses of plants to pathogens. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 1995; pp. 1–34. ISBN 0120059215. [Google Scholar]
- Chalfoun, N.R.; Castagnaro, A.; Díaz Ricci, J.C. Induced resistance activated by a culture filtrate derived from an avirulent pathogen as a mechanism of biological control of anthracnose in strawberry. Biol. Control 2011, 58, 319–329. [Google Scholar] [CrossRef]
- Elías, J.M.; Guerrero-Molina, M.F.; Martínez-Zamora, M.G.; Díaz-Ricci, J.C.; Pedraza, R.O. Role of ethylene and related gene expression in the interaction between strawberry plants and the plant growth-promoting bacterium Azospirillum brasilense. Plant Biol. 2018, 20, 490–496. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Fukami, J.; Ollero, F.J.; de la Osa, C.; Valderrama-Fernández, R.; Nogueira, M.A.; Megías, M.; Hungria, M. Antioxidant activity and induction of mechanisms of resistance to stresses related to the inoculation with Azospirillum brasilense. Arch. Microbiol. 2018, 200, 1191–1203. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic Acquired Resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Cellulase | Protease | Amylase | Chitinase | HCN |
|---|---|---|---|---|---|
| 2A1 | ++ | - | - | - | - |
| 2A2 | +++ | - | - | - | - |
| 2E1 | ++ | - | - | - | - |
| REC3 | + | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viejobueno, J.; Albornoz, P.L.; Camacho, M.; de los Santos, B.; Martínez-Zamora, M.G.; Salazar, S.M. Protection of Strawberry Plants against Charcoal Rot Disease (Macrophomina phaseolina) Induced by Azospirillum brasilense. Agronomy 2021, 11, 195. https://doi.org/10.3390/agronomy11020195
Viejobueno J, Albornoz PL, Camacho M, de los Santos B, Martínez-Zamora MG, Salazar SM. Protection of Strawberry Plants against Charcoal Rot Disease (Macrophomina phaseolina) Induced by Azospirillum brasilense. Agronomy. 2021; 11(2):195. https://doi.org/10.3390/agronomy11020195
Chicago/Turabian StyleViejobueno, Josefina, Patricia Liliana Albornoz, María Camacho, Berta de los Santos, Martín Gustavo Martínez-Zamora, and Sergio Miguel Salazar. 2021. "Protection of Strawberry Plants against Charcoal Rot Disease (Macrophomina phaseolina) Induced by Azospirillum brasilense" Agronomy 11, no. 2: 195. https://doi.org/10.3390/agronomy11020195
APA StyleViejobueno, J., Albornoz, P. L., Camacho, M., de los Santos, B., Martínez-Zamora, M. G., & Salazar, S. M. (2021). Protection of Strawberry Plants against Charcoal Rot Disease (Macrophomina phaseolina) Induced by Azospirillum brasilense. Agronomy, 11(2), 195. https://doi.org/10.3390/agronomy11020195