
Reducing N Fertilization without Yield Penalties in Maize with a Commercially Available Seed Dressing

 ,
,  ,
,  , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
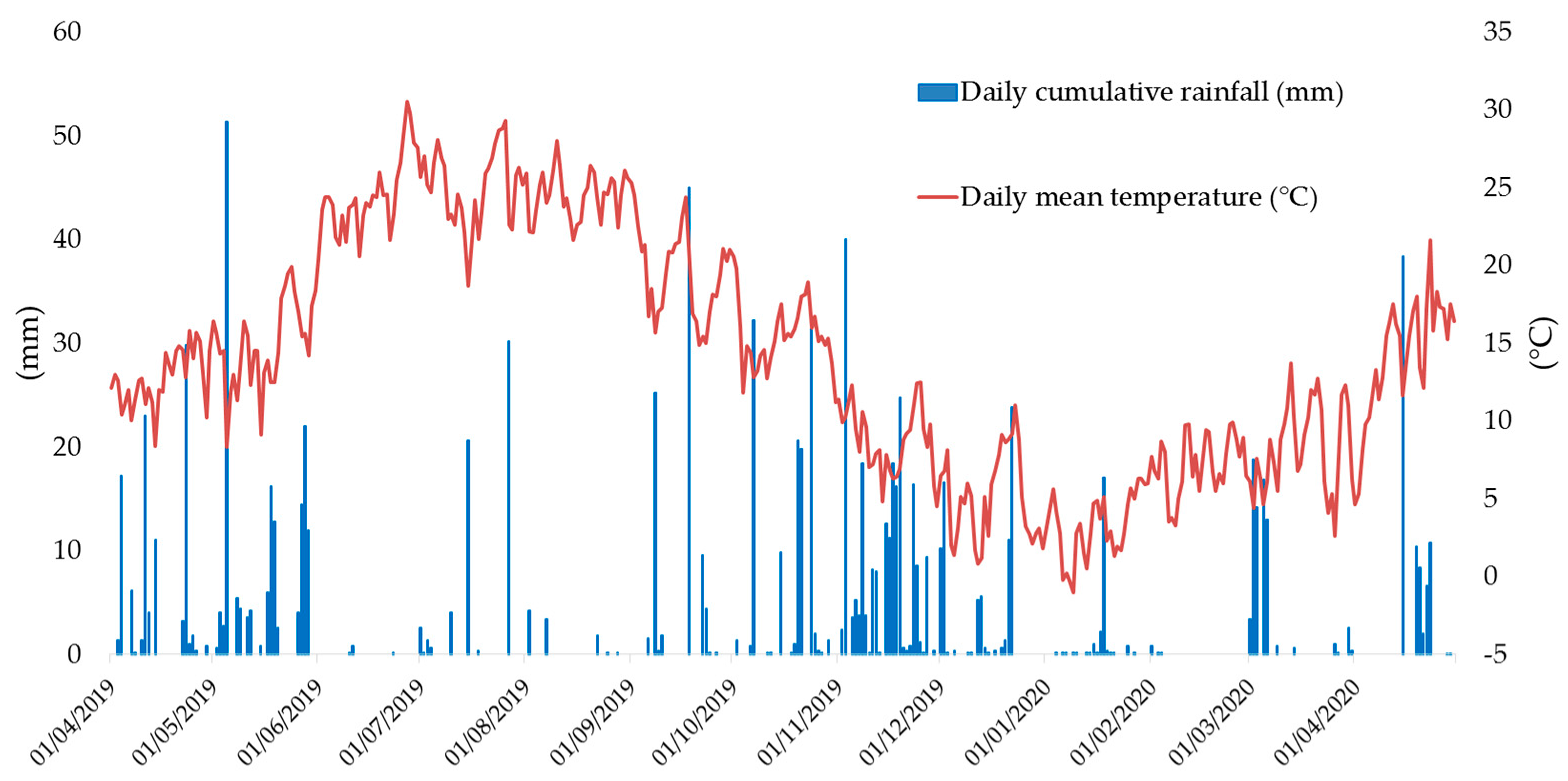
2.1. Site and Soil Characteristics
2.2. Experimental Design, Treatments and Crop Management
2.3. Measurements of Maize Grain Yield and Root Density
2.4. Soil DNA Extraction, Amplification and Bioinformatics Analyses
2.5. Gas Sampling and Quantification
2.6. Soil Properties Affecting GHGs
2.7. Statistical Analyses
3. Results
3.1. Grain Yield of Maize
3.2. Root Length Density (RLD) and Diameter Class Length (DCL)
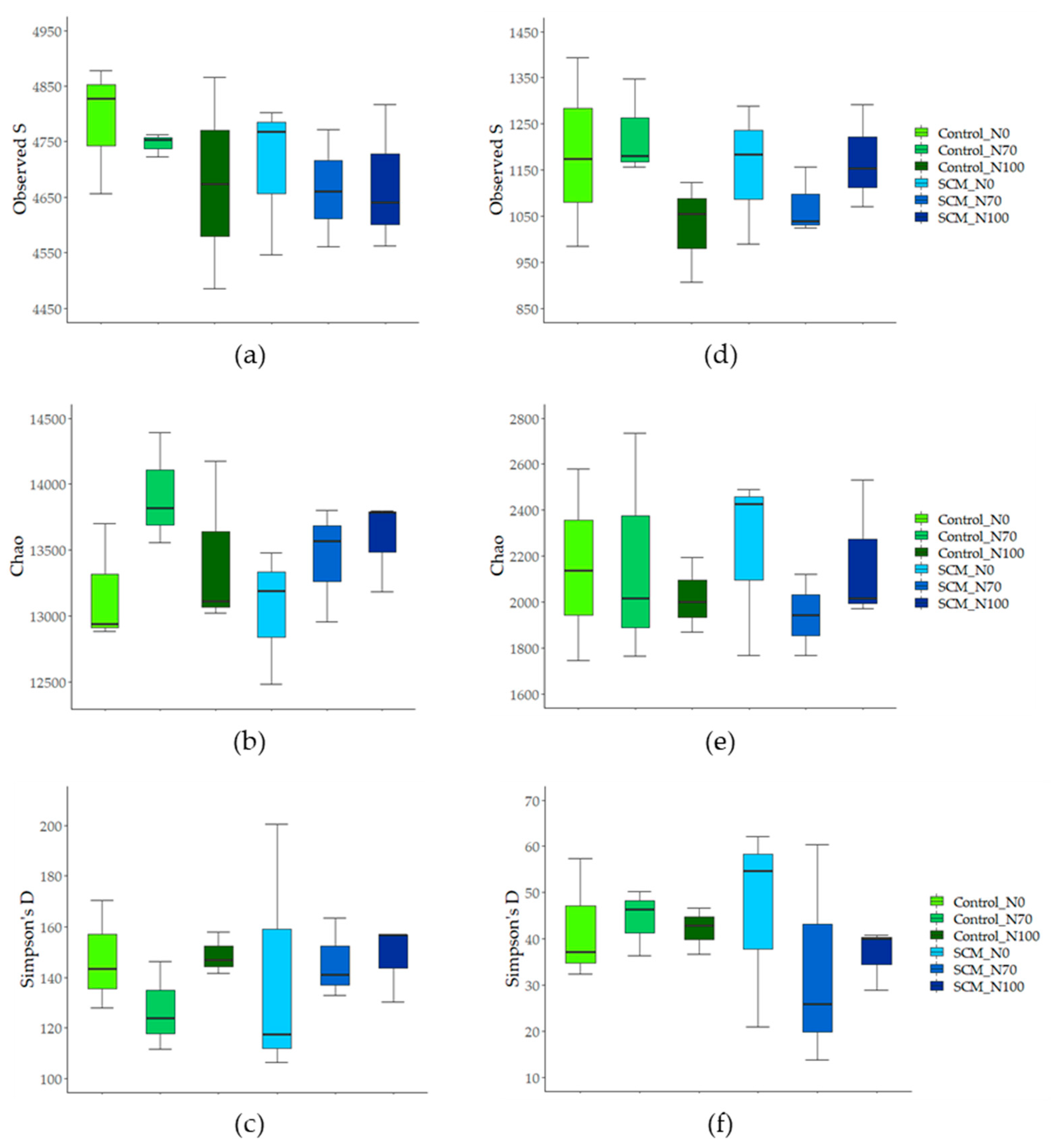
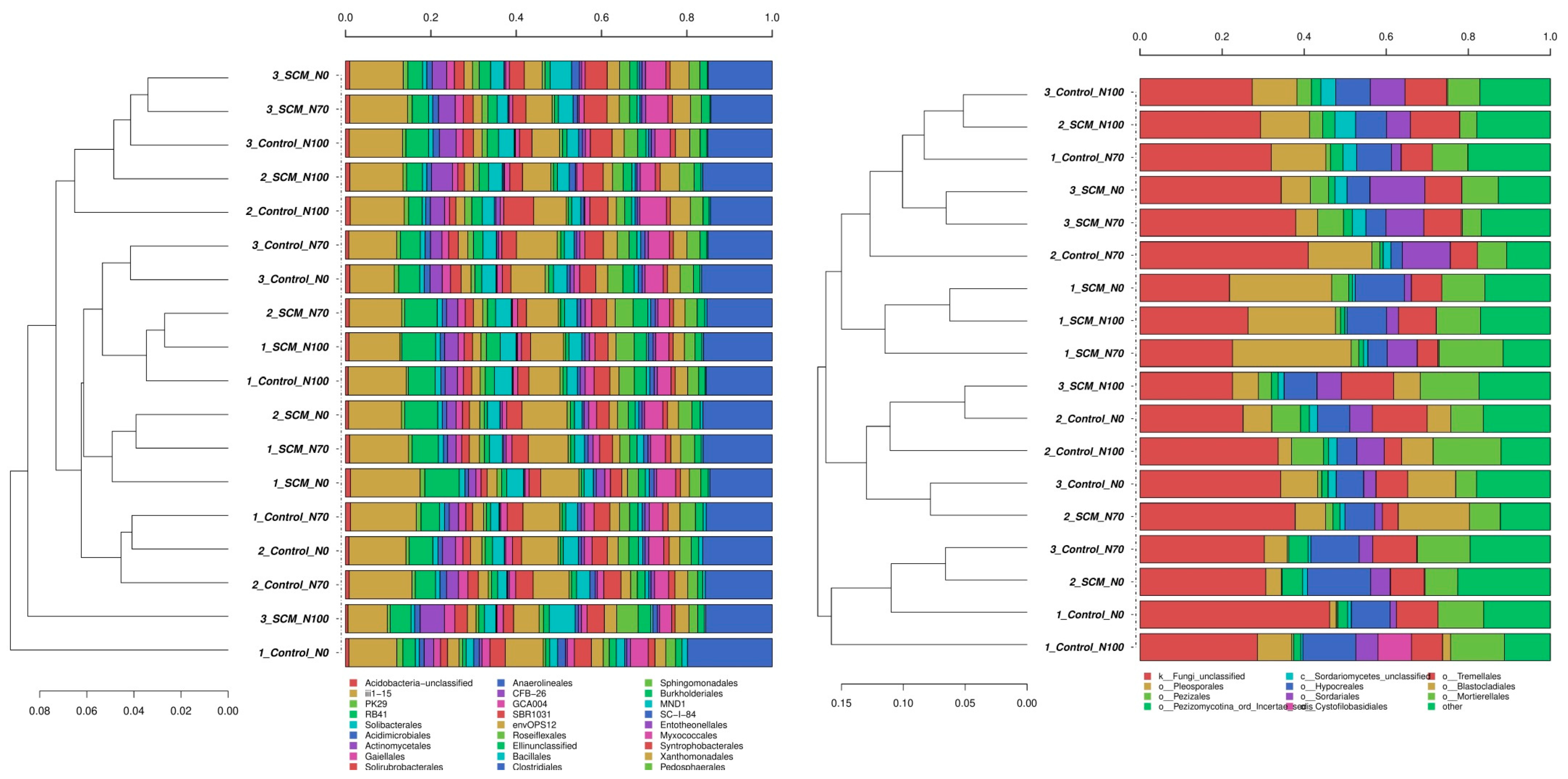
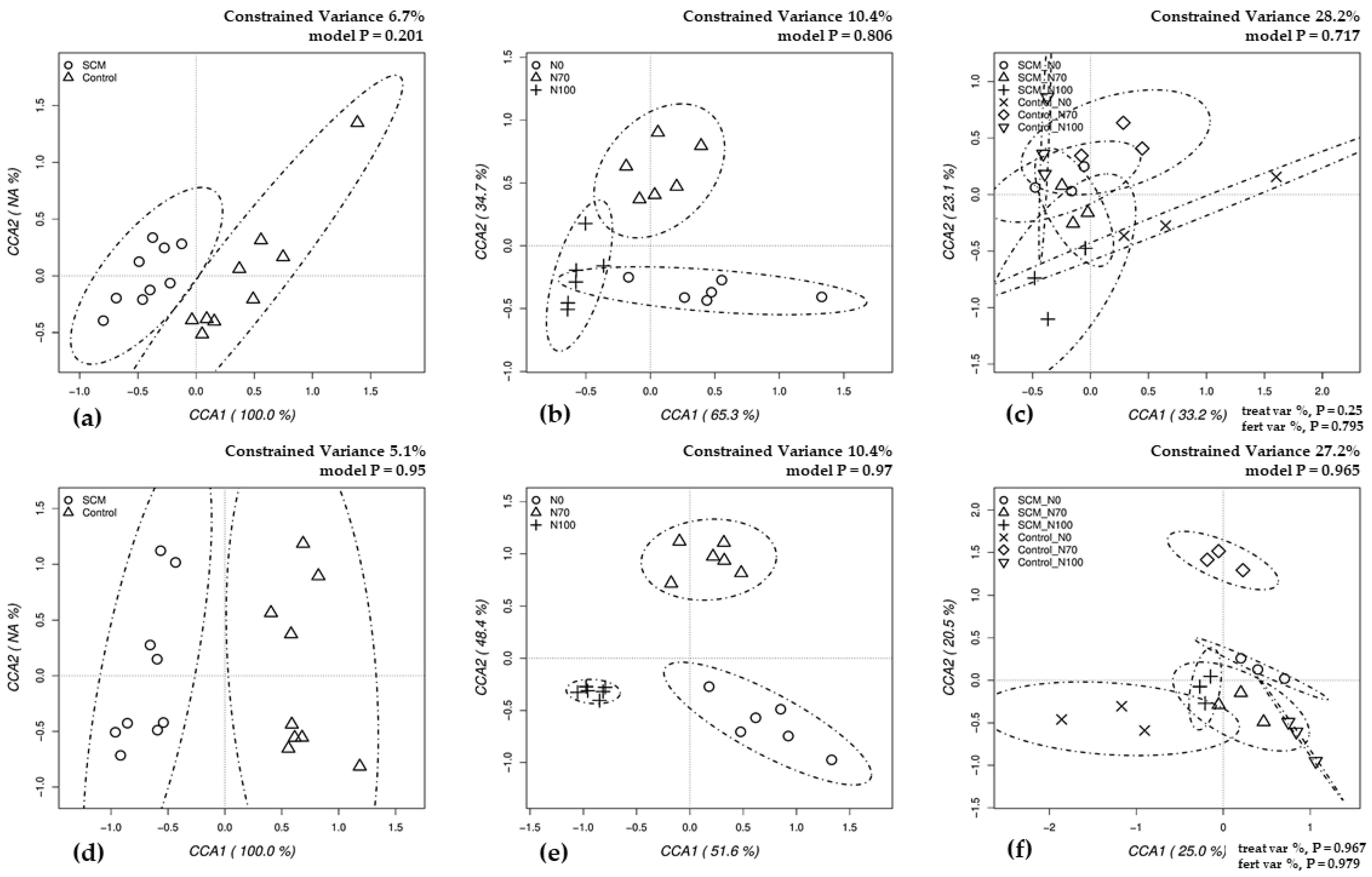
3.3. Biodiversity of Soil Bacteria and Fungi
3.4. Greenhouse Gas Emissions: Nitrous Oxide, Carbon Dioxide and Methane
4. Discussion
4.1. Responses of Maize Grain Yield, RLD, and DCL to SCM and N-Fertilization
4.2. Relationships between Treatments and Biodiversity of Soil Bacteria and Fungi
4.3. Nitrous Oxide, Carbon Dioxide, and Methane Emissions as Affected by SCM and N-Fertilization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fiorini, A.; Boselli, R.; Maris, S.C.; Santelli, S.; Perego, A.; Acutis, M.; Brenna, S.; Tabaglio, V. Soil type and cropping system as drivers of soil quality indicators response to no-till: A 7-year field study. Appl. Soil Ecol. 2020, 155, 103646. [Google Scholar] [CrossRef]
- Buckwell, A.; Nadeu, E. Nutrient Recovery and Reuse (NRR) in European Agriculture. A Review issues, Opportunities and Actions; RISE Foundundation: Brussels, Belgium, 2016; Available online: https://www.organicseurope.bio/content/uploads/2020/06/2016_RISE_NRR_Full_EN_compressed.pdf?dd (accessed on 15 December 2020).
- Jones, A.; Panagos, P.; Barcelo, S.; Bouraoui, F.; Bosco, C.; Dewitte, O.; Gardi, C.; Erhard, M.; Hervás, J.; Hiederer, R.; et al. The State of Soil in Europe: A Contribution of the JRC to the European Environment Agency’s Environment State and Outlook Report—SOER 2010; European Commission: Luxemburg, 2012. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO, Organization of Economic Co-operation and Development (OECD). Food Security and Nutrition: Challenges for Agriculture and the Hidden Potential of Soil. A Report to the G20 Agriculture Deputies; FAO and OECD: Rome, Italy, 2018; Available online: http://www.fao.org/3/CA0917EN/ca0917en.pdf (accessed on 15 December 2020).
- Lipper, L.; McCarthy, N.; Zilberman, D.; Asfaw, S.; Branca, G. Climate Smart Agriculture: Building Resilience to Climate Change; Springer Nature: London, UK, 2018; ISBN 978-3-319-61193-8. [Google Scholar]
- Chen, L.; Dick, W.A.; Smith, K.L. Gypsum as an Agricultural Amendment: General Use Guidelines; Bulletin 945; OSU—Ohio State University Extension: Columbus, OH, USA, 2011; Available online: https://fabe.osu.edu/sites/fabe/files/imce/files/Soybean/Gypsum%20Bulletin.pdf (accessed on 15 December 2020).
- Shainberg, I.; Sumner, M.E.; Miller, W.P.; Farina, M.P.W.; Pavan, M.A.; Fey, M.V. Use of Gypsum on Soils: A Review. In Advances in Soil Science; Springer: Berlin/Heidelberg, Germany, 1989; Volume 9, pp. 1–111. ISBN 978-1-4612-8144-3. [Google Scholar] [CrossRef]
- Zoca, S.M.; Penn, C. An important tool with no instruction manual: A review of gypsum use in agriculture. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2017; Volume 144, pp. 1–44. ISBN 9780128124192. [Google Scholar] [CrossRef]
- Kost, D.; Ladwig, K.J.; Chen, L.; DeSutter, T.M.; Espinoza, L.; Norton, L.D.; Smeal, D.; Torbert, H.A.; Watts, D.B.; Wolkowski, R.P.; et al. Meta-Analysis of Gypsum Effects on Crop Yields and Chemistry of Soils, Plant Tissues, and Vadose Water at Various Research Sites in the USA. J. Environ. Qual. 2018, 47, 1284–1292. [Google Scholar] [CrossRef]
- Ritchey, K.D.; Feldhake, C.M.; Clark, R.B.; De Sousa, D.M.G.; Karlen, D.; Wright, R.; Kemper, W. Improved Water and Nutrient Uptake from Subsurface Layers of Gypsum-Amended Soils. Soil Eros. Conserv. Trop. 2015, 58, 157–181. [Google Scholar] [CrossRef]
- Da Silva, E.A.; de Oliveira, G.C.; Carducci, C.E.; Silva, B.M.; de Oliveira, L.M.; Costa, J.C. Increasing doses of agricultural gypsum, aggregate stability and organic carbon in Cerrado Oxisol under coffee crop. Rev. Ciências Agrárias/Amazon. J. Agric. Environ. Sci. 2013, 56, 25–32. [Google Scholar]
- Basu, S.; Kumar, G.; Chhabra, S.; Prasad, R. Role of soil microbes in biogeochemical cycle for enhancing soil fertility. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier BV: Amsterdam, The Netherlands, 2021; pp. 149–157. ISBN 9780444635075. [Google Scholar]
- Kurzemann, F.R.; Plieger, U.; Probst, M.; Spiegel, H.; Sandén, T.; Ros, M.; Insam, H. Long-Term Fertilization Affects Soil Microbiota, Improves Yield and Benefits Soil. Agronomy 2020, 10, 1664. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Wei, D.; Zhao, B.; Ma, M.; Chen, S.; Cao, F.; Shen, D.; Guan, D.; Li, J. Consistent effects of nitrogen fertilization on soil bacterial communities in black soils for two crop seasons in China. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nifong, R.L.; Taylor, J.M.; Moore, M.T. Mulch-Derived Organic Carbon Stimulates High Denitrification Fluxes from Agricultural Ditch Sediments. J. Environ. Qual. 2019, 48, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Ma, F.; Xiao, G. Effect of varying fertilization patterns on bacteria and archaea communities in saline-alkali soil under rice cultivation. J. Agro-Environ. Sci. 2018, 37, 495–504. [Google Scholar]
- Holland, J.; Bennett, A.; Newton, A.; White, P.; McKenzie, B.; George, T.; Pakeman, R.; Bailey, J.; Fornara, D.; Hayes, R.; et al. Liming impacts on soils, crops and biodiversity in the UK: A review. Sci. Total. Environ. 2018, 316–332. [Google Scholar] [CrossRef]
- Pariz, C.M.; Costa, C.; Crusciol, C.A.; Castilhos, A.M.; Meirelles, P.R.; Roça, R.O.; Pinheiro, R.S.; Kuwahara, F.A.; Martello, J.M.; Cavasano, F.A.; et al. Lamb production responses to grass grazing in a companion crop system with corn silage and oversowing of yellow oat in a tropical region. Agric. Syst. 2017, 151, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ladha, J.K.; Jat, M.L.; Stirling, C.M.; Chakraborty, D.; Pradhan, P.; Krupnik, T.J.; Sapkota, T.B.; Pathak, H.; Rana, D.S.; Tesfaye, K.; et al. Chapter Two – Achieving the sustainable development goals in agriculture: The crucial role of nitrogen in cereal-based systems. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 163, pp. 39–116. [Google Scholar]
- European Commission Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. A Farm to Fork Strategy for a fair, Healthy and Environmentally-Friendly Food System, Brussels, 20.5.2020 COM(2020) 381 Final (2020). Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=COM:2020:0381:FIN (accessed on 15 December 2020).
- Good, A.G.; Beatty, P.H. Fertilizing Nature: A Tragedy of Excess in the Commons. PLoS Biol. 2011, 9, e1001124. [Google Scholar] [CrossRef]
- Buendia, E.C.; Guendehou, S.; Limmeechokchai, B.; Pipatti, R.; Rojas, Y.; Sturgiss, R.; Tanabe, K.; Wirth, T.; Romano, D.; Witi, J.; et al. 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories — IPCC. Available online: https://www.ipcc.ch/site/assets/uploads/2019/12/19R_V0_01_Overview.pdf (accessed on 15 December 2020).
- Chantigny, M.H.; Rochette, P.; Angers, D.A.; Bittman, S.; Buckley, K.; Massé, D.; Bélanger, G.; Eriksen-Hamel, N.; Gasser, M.-O. Soil Nitrous Oxide Emissions Following Band-Incorporation of Fertilizer Nitrogen and Swine Manure. J. Environ. Qual. 2010, 39, 1545–1553. [Google Scholar] [CrossRef]
- Venterea, R.T.; Strock, J.; Rosen, C. Agricultural management effects on nitrous oxide gas emissions. In Proceedings of the Lamberton and Outreach Center Soil and Water Management Field Day, Lamberton, MN, USA, 13 August 2008; p. 8. Available online: https://naldc.nal.usda.gov/download/29362/PDF (accessed on 15 December 2020).
- Davidson, E.A. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nat. Geosci. 2009, 2, 659–662. [Google Scholar] [CrossRef]
- Syakila, A.; Kroeze, C. The global nitrous oxide budget revisited. Greenh. Gas Meas. Manag. 2011, 1, 17–26. [Google Scholar] [CrossRef]
- Niu, S.; Wu, M.; Han, Y.; Xia, J.; Zhang, Z.; Yang, H.; Wan, S. Nitrogen effects on net ecosystem carbon exchange in a temperate steppe. Glob. Chang. Biol. 2010, 16, 144–155. [Google Scholar] [CrossRef]
- Morell, F.; Álvaro-Fuentes, J.; Lampurlanés, J.; Cantero-Martínez, C. Soil CO2 fluxes following tillage and rainfall events in a semiarid Mediterranean agroecosystem: Effects of tillage systems and nitrogen fertilization. Agric. Ecosyst. Environ. 2010, 139, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Snyder, C.; Bruulsema, T.; Jensen, T.; Fixen, P. Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agric. Ecosyst. Environ. 2009, 133, 247–266. [Google Scholar] [CrossRef]
- Barton, L.; Murphy, D.V.; Butterbach-Bahl, K. Influence of crop rotation and liming on greenhouse gas emissions from a semi-arid soil. Agric. Ecosyst. Environ. 2013, 167, 23–32. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Roy, R.; Ethier, G. Effect of nitrogen fertilization on soil CH4 and N2O fluxes, and soil and bole respiration. Geoderma 2011, 162, 182–186. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA. Available online: https://www.nrcs.usda.gov/wps/PA_NRCSConsumption/download?cid=stelprdb1252094&ext=pdf (accessed on 15 December 2020).
- Sainju, U.M. Determination of nitrogen balance in agroecosystems. MethodsX 2017, 4, 199–208. [Google Scholar] [CrossRef]
- European Commission. Directive 91/676/EEC. Council Directive of 12 December 1991 concerning the protection of waters against pollution caused by nitrates from agricultural sources. Off. J. Eur. Community 1991, 375, 1–8. [Google Scholar]
- Fiorini, A.; Boselli, R.; Maris, S.C.; Santelli, S.; Ardenti, F.; Capra, F.; Tabaglio, V. May conservation tillage enhance soil C and N accumulation without decreasing yield in intensive irrigated croplands? Results from an eight-year maize monoculture. Agric. Ecosyst. Environ. 2020, 296, 106926. [Google Scholar] [CrossRef]
- Qin, R.; Stamp, P.; Richner, W. Impact of tillage on maize rooting in a Cambisol and Luvisol in Switzerland. Soil Tillage Res. 2006, 85, 50–61. [Google Scholar] [CrossRef]
- Fiorini, A.; Boselli, R.; Amaducci, S.; Tabaglio, V. Effects of no-till on root architecture and root-soil interactions in a three-year crop rotation. Eur. J. Agron. 2018, 99, 156–166. [Google Scholar] [CrossRef]
- Reinhardt, D.R.; Miller, R.M. Size classes of root diameter and mycorrhizal fungal colonization in two temperate grassland communities. New Phytol. 1990, 116, 129–136. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Cocconcelli, P.S.; Trevisan, M. Soil bacterial diversity screening using single 16S rRNA gene V regions coupled with multi-million read generating sequencing technologies. PLoS ONE 2012, 7, e42671. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.A.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91, fiv114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Van Veen, J.A.; Cocconcelli, P.S.; Trevisan, M. Soil microbial diversity patterns of a lowland spring environment. FEMS Microbiol. Ecol. 2013, 86, 172–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Wien, Austria, 2013; ISBN 3-900051-07-01. [Google Scholar]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z., Jr.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. NAST: A multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef]
- Schloss, P.D. The Effects of Alignment Quality, Distance Calculation Method, Sequence Filtering, and Region on the Analysis of 16S rRNA Gene-Based Studies. PLoS Comput. Biol. 2010, 6, e1000844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.K.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.J.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2011, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Rolston, D.E. Gas Flux. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods, 5.1, 2nd ed.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1986; pp. 1103–1119. ISBN 9780891180883. [Google Scholar] [CrossRef]
- Maris, S.C.; Lloveras, J.; Vallejo, A.; Teira-Esmatges, M.R. Effect of Stover Management and Nitrogen Fertilization on N2O and CO2 Emissions from Irrigated Maize in a High Nitrate Mediterranean Soil. Water Air Soil Pollut. 2018, 229, 11. [Google Scholar] [CrossRef]
- Peyron, M.; Bertora, C.; Pelissetti, S.; Said-Pullicino, D.; Celi, L.; Miniotti, E.; Romani, M.; Sacco, D. Greenhouse gas emissions as affected by different water management practices in temperate rice paddies. Agric. Ecosyst. Environ. 2016, 232, 17–28. [Google Scholar] [CrossRef]
- Maris, S.; Teira-Esmatges, M.; Arbones, A.; Rufat, J. Effect of irrigation, nitrogen application, and a nitrification inhibitor on nitrous oxide, carbon dioxide and methane emissions from an olive (Olea europaea L.) orchard. Sci. Total. Environ. 2015, 538, 966–978. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984; ISBN 978-0-471-87092-0. [Google Scholar]
- Doussan, C.; Pierret, A.; Garrigues, E.; Pagès, L. Water Uptake by Plant Roots: II-Modelling of Water Transfer in the Soil Root-system with Explicit Account of Flow within the Root System-Comparison with Experiments. Plant Soil 2006, 283, 99–117. [Google Scholar] [CrossRef]
- Ma, Q.; Tang, H.; Rengel, Z.; Shen, J. Banding phosphorus and ammonium enhances nutrient uptake by maize via modifying root spatial distribution. Crop. Pasture Sci. 2013, 64, 965–975. [Google Scholar] [CrossRef]
- Wen, Z.; Shen, J.; Blackwell, M.; Li, H.; Zhao, B.; Yuan, H. Combined Applications of Nitrogen and Phosphorus Fertilizers with Manure Increase Maize Yield and Nutrient Uptake via Stimulating Root Growth in a Long-Term Experiment. Pedosphere 2016, 26, 62–73. [Google Scholar] [CrossRef]
- Maris, S.C.; Fiorini, A.; Boselli, R.; Santelli, S.; Tabaglio, V. Cover crops, compost, and conversion to grassland to increase soil C and N stock in intensive agrosystems. Nutr. Cycl. Agroecosyst. 2021, 119, 83–101. [Google Scholar] [CrossRef]
- Da Costa, C.H.M.; Crusciol, C.A.C. Long-term effects of lime and phosphogypsum application on tropical no-till soybean–oat–sorghum rotation and soil chemical properties. Eur. J. Agron. 2016, 74, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Crusciol, C.A.; Artigiani, A.C.; Arf, O.; Filho, A.C.C.; Soratto, R.P.; Nascente, A.S.; Alvarez, R.C. Soil fertility, plant nutrition, and grain yield of upland rice affected by surface application of lime, silicate, and phosphogypsum in a tropical no-till system. Catena 2016, 137, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Caires, E.; Garbuio, F.; Churka, S.; Barth, G.; Corrêa, J. Effects of soil acidity amelioration by surface liming on no-till corn, soybean, and wheat root growth and yield. Eur. J. Agron. 2008, 28, 57–64. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press, Elsevier: London, UK, 2011; ISBN 9780123849052. [Google Scholar]
- Hammad, H.M.; Ahmad, A.; Abbas, F.; Farhad, W.; Cordoba, B.C.; Hoogenboom, G. Water and Nitrogen Productivity of Maize under Semiarid Environments. Crop. Sci. 2015, 55, 877–888. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Motes, C.M.; Scheible, W.-R.; Chen, R.; Blancaflor, E.B.; Monteros, M.J. Root Traits and Phenotyping Strategies for Plant Improvement. Plants 2015, 4, 334–355. [Google Scholar] [CrossRef]
- Hu, X.-Y.; Xiang, Q.-J.; Mu, Z.-J. [Effects of Gypsum on CH4 Emission and Functional Microbial Communities in Paddy Soil]. Huan Jing Ke Xue= Huanjing Kexue 2018, 39, 3894–3900. [Google Scholar]
- Wörner, S.; Zecchin, S.; Dan, J.; Todorova, N.H.; Loy, A.; Conrad, R.; Pester, M. Gypsum amendment to rice paddy soil stimulated bacteria involved in sulfur cycling but largely preserved the phylogenetic composition of the total bacterial community. Environ. Microbiol. Rep. 2016, 8, 413–423. [Google Scholar] [CrossRef]
- Cavaglieri, L.; Orlando, J.; Etcheverry, M. Rhizosphere microbial community structure at different maize plant growth stages and root locations. Microbiol. Res. 2009, 164, 391–399. [Google Scholar] [CrossRef]
- Phazna Devi, T.A.; Sahoo, D.; Setti, A.; Sharma, C.; Kalita, M.C.; Indira Devi, S. Bacterial rhizosphere community profile at different growth stages of Umorok (Capsicum chinense) and its response to the root exudates. Int. Microbiol. 2019, 23, 241–251. [Google Scholar] [CrossRef]
- Leite, M.F.A.; Pan, Y.; Bloem, J.; Berge, H.T.; Kuramae, E.E. Organic nitrogen rearranges both structure and activity of the soil-borne microbial seedbank. Sci. Rep. 2017, 7, srep42634. [Google Scholar] [CrossRef]
- Fiorini, A.; Maris, S.C.; Abalos, D.; Amaducci, S.; Tabaglio, V. Combining no-till with rye (Secale cereale L.) cover crop mitigates nitrous oxide emissions without decreasing yield. Soil Tillage Res. 2020, 196, 104442. [Google Scholar] [CrossRef]
- Skiba, U.; Ball, B. The effect of soil texture and soil drainage on emissions of nitric oxide and nitrous oxide. Soil Use Manag. 2006, 18, 56–60. [Google Scholar] [CrossRef]
- Yao, Z.; Zhou, Z.; Zheng, X.; Xie, B.; Mei, B.; Wang, R.; Butterbach-Bahl, K.; Zhu, J. Effects of organic matter incorporation on nitrous oxide emissions from rice-wheat rotation ecosystems in China. Plant Soil 2009, 327, 315–330. [Google Scholar] [CrossRef]
- Abalos, D.; Sanchez-Martin, L.; Garcia-Torres, L.; Van Groenigen, J.W.; Vallejo, A. Management of irrigation frequency and nitrogen fertilization to mitigate GHG and NO emissions from drip-fertigated crops. Sci. Total. Environ. 2014, 490, 880–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Walter, M.T.; Drinkwater, L.E. N2O emissions from grain cropping systems: A meta-analysis of the impacts of fertilizer-based and ecologically-based nutrient management strategies. Nutr. Cycl. Agroecosyst. 2017, 107, 335–355. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Emissions of N2O and NO from fertilized fields: Summary of available measurement data. Glob. Biogeochem. Cycles 2002, 16, 1058. [Google Scholar] [CrossRef]
- Hansen, S.; Frøseth, R.B.; Stenberg, M.; Stalenga, J.; Olesen, J.E.; Krauss, M.; Radzikowski, P.; Doltra, J.; Nadeem, S.; Torp, T.; et al. Reviews and syntheses: Review of causes and sources of N2O emissions and NO3 leaching from organic arable crop rotations. Biogeosciences 2019, 16, 2795–2819. [Google Scholar] [CrossRef] [Green Version]
- Shcherbak, I.; Millar, N.; Robertson, G.P. Global metaanalysis of the nonlinear response of soil nitrous oxide (N2O) emissions to fertilizer nitrogen. Proc. Natl. Acad. Sci. USA 2014, 111, 9199–9204. [Google Scholar] [CrossRef] [Green Version]
- Firestone, M.K.; Davidson, E.A. Microbiological basis of NO and N2O production and consumption in soil. In Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere; Andreae, M.O., Schimel, D.S., Eds.; John Willey and Sons: New York, NY, USA, 1989; pp. 7–21. [Google Scholar]
- Mosier, A.R.; Halvorson, A.D.; Reule, C.A.; Liu, X.J. Net Global Warming Potential and Greenhouse Gas Intensity in Irrigated Cropping Systems in Northeastern Colorado. J. Environ. Qual. 2006, 35, 1584–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapuis-Lardy, L.; Wrage, N.; Metay, A.; Chotte, J.-L.; Bernoux, M. Soils, a sink for N2O? A review. Glob. Chang. Biol. 2007, 13, 1–17. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Cantero-Martínez, C.; Bareche, J.; Arrúe, J.L.; Álvaro-Fuentes, J. Soil carbon dioxide and methane fluxes as affected by tillage and N fertilization in dryland conditions. Plant Soil 2014, 381, 111–130. [Google Scholar] [CrossRef] [Green Version]
- Deforest, J.L.; Zak, D.R.; Pregitzer, K.S.; Burton, A.J. Atmospheric Nitrate Deposition, Microbial Community Composition, and Enzyme Activity in Northern Hardwood Forests. Soil Sci. Soc. Am. J. 2004, 68, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.J.; Pregitzer, K.S.; Crawford, J.N.; Zogg, G.P.; Zak, D.R. Simulated chronic NO3 - deposition reduces soil respiration in northern hardwood forests. Glob. Chang. Biol. 2004, 10, 1080–1091. [Google Scholar] [CrossRef] [Green Version]
- Abalos, D.; Sanz-Cobena, A.; Garcia, A.V.; Van Groenigen, J.W.; Vallejo, A. Role of maize stover incorporation on nitrogen oxide emissions in a non-irrigated Mediterranean barley field. Plant Soil 2013, 364, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Gulledge, J.; Hrywna, Y.; Cavanaugh, C.; Steudler, P.A. Effects of long-term nitrogen fertilization on the uptake kinetics of atmospheric methane in temperate forest soils. FEMS Microbiol. Ecol. 2004, 49, 389–400. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Main Factor | Secondary Factor | Grain Yield (Mg ha−1) |
|---|---|---|---|
| presence/absence of SOP® COCUS MAIZE (SCM) (P/A) | SCM | - | 13.36 ± 2.14 a |
| Control | - | 12.35 ± 1.85 b | |
| p (F) | <0.01 | ||
| N-fertilization rate (N) | - | 0% N-fertilization | 10.34 ± 0.59 c |
| - | 70% N-fertilization | 13.49 ± 0.79 b | |
| - | 100% N-fertilization | 14.74 ± 0.83 a | |
| p (F) | <0.001 | ||
| P/A × N | SCM | 0% N-fertilization | 10.63 ± 0.40 |
| 70% N-fertilization | 14.14 ± 0.32 | ||
| 100% N-fertilization | 15.32 ± 0.83 | ||
| Control | 0% N-fertilization | 10.05 ± 0.65 | |
| 70% N-fertilization | 12.85 ± 0.50 | ||
| 100% N-fertilization | 14.17 ± 0.22 | ||
| p (F) | n.s. |
| Source of Variation | Main Factor | Secondary Factor | RLD (cm cm−3) |
|---|---|---|---|
| presence/absence of SCM (P/A) | SCM | - | 3.71 ± 1.45 a |
| Control | - | 3.32 ± 1.17 b | |
| p (F) | <0.05 | ||
| N-fertilization rate (N) | - | 0% N-fertilization | 2.70 ± 0.80 c |
| - | 70% N-fertilization | 3.54 ± 1.19 b | |
| - | 100% N-fertilization | 4.30 ± 1.42 a | |
| p (F) | <0.001 | ||
| P/A × N | SCM | 0% N-fertilization | 2.80 ± 0.96 |
| 70% N-fertilization | 3.85 ± 1.45 | ||
| 100% N-fertilization | 4.47 ± 1.56 | ||
| Control | 0% N-fertilization | 2.59 ± 0.67 | |
| 70% N-fertilization | 3.23 ± 0.88 | ||
| 100% N-fertilization | 4.13 ± 1.40 | ||
| p (F) | n.s. |
| 0% N-Fertilization | 70% N-Fertilization | 100% N-Fertilization | |||||
|---|---|---|---|---|---|---|---|
| SCM | Control | SCM | Control | SCM | Control | ||
| Root Diameter Class | Distance from the Row | DCL (cm cm−3) | DCL (cm cm−3) | DCL (cm cm−3) | |||
| ø = 0–0.075 mm | 0 cm | 0.54 ± 0.16 | 0.49 ± 0.05 | 1.20 ± 0.08 a | 0.83 ± 0.10 b | 1.24 ± 0.17 a | 0.89 ± 0.19 b |
| 37.5 cm | 0.30 ± 0.16 | 0.30 ± 0.10 | 0.43 ± 0.07 | 0.50 ± 0.16 | 0.62 ± 0.10 | 0.56 ± 0.10 | |
| ø = 0.075–0.2 mm | 0 cm | 1.39 ± 0.24 | 1.31 ± 0.21 | 2.25 ± 0.14 a | 1.48 ± 0.29 b | 2.20 ± 0.11 | 2.05 ± 0.37 |
| 37.5 cm | 0.68 ± 0.08 | 0.82 ± 0.06 | 1.02 ± 0.22 | 1.02 ± 0.10 | 1.30 ± 0.26 | 1.07 ± 0.12 | |
| ø = 0.2–1 mm | 0 cm | 1.55 ± 0.30 | 1.29 ± 0.26 | 1.93 ± 0.10 a | 1.46 ± 0.16 b | 2.23 ± 0.57 | 2.00 ± 0.46 |
| 37.5 cm | 1.02 ± 0.14 | 0.88 ± 0.09 | 1.06 ± 0.28 | 0.93 ± 0.32 | 1.17 ± 0.24 | 1.41 ± 0.29 | |
| ø > 1 mm | 0 cm | 0.10 ± 0.05 | 0.06 ± 0.01 | 0.08 ± 0.01 b | 0.20 ± 0.01 a | 0.15 ± 0.03 b | 0.27 ± 0.12 a |
| 37.5 cm | 0.01 ± 0.01 | 0.03 ± 0.02 | 0.02 ± 0.01 | 0.03 ± 0.03 | 0.03 ± 0.02 | 0.02 ± 0.01 | |
| Source of Variation | Main Factor | Secondary Factor | N2O (mg N-N2O m−2 y−1) | CO2 (g C-CO2 m−2 y−1) | CH4 (mg C-CH4 m−2 y−1) |
|---|---|---|---|---|---|
| presence/absence of SCM (P/A) | SCM | - | 1171 ± 627 | 3230 ± 925 | −110 ± 47 |
| Control | - | 1067 ± 574 | 3220 ± 1032 | −75 ± 56 | |
| p (F) | n.s. | n.s. | n.s. | ||
| N-fertilization rate (N) | - | 0% N-fertilization | 464 ± 131 a | 4135 ± 652 a | −78 ± 57 |
| - | 70% N-fertilization | 1192 ± 234 b | 2689 ± 855 b | −80 ± 53 | |
| - | 100% N-fertilization | 1701 ± 408 c | 2850 ± 641 b | −120 ± 47 | |
| p (F) | < 0.001 | < 0.01 | n.s. | ||
| P/A × N | SCM | 0% N-fertilization | 494 ± 118 | 4219 ± 584 | −93 ± 40 |
| 70% N-fertilization | 1253 ± 318 | 2415 ± 408 | −115 ± 39 | ||
| 100% N-fertilization | 1766 ± 479 | 3057 ± 639 | −122 ± 70 | ||
| Control | 0% N-fertilization | 434 ± 162 | 4051 ± 838 | −62 ± 76 | |
| 70% N-fertilization | 1132 ± 158 | 2964 ± 1197 | −45 ± 44 | ||
| 100% N-fertilization | 1635 ± 417 | 2643 ± 700 | −117 ± 25 | ||
| p (F) | n.s. | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maris, S.C.; Capra, F.; Ardenti, F.; Chiodini, M.E.; Boselli, R.; Taskin, E.; Puglisi, E.; Bertora, C.; Poggianella, L.; Amaducci, S.; et al. Reducing N Fertilization without Yield Penalties in Maize with a Commercially Available Seed Dressing. Agronomy 2021, 11, 407. https://doi.org/10.3390/agronomy11030407
Maris SC, Capra F, Ardenti F, Chiodini ME, Boselli R, Taskin E, Puglisi E, Bertora C, Poggianella L, Amaducci S, et al. Reducing N Fertilization without Yield Penalties in Maize with a Commercially Available Seed Dressing. Agronomy. 2021; 11(3):407. https://doi.org/10.3390/agronomy11030407
Chicago/Turabian StyleMaris, Stefania Codruta, Federico Capra, Federico Ardenti, Marcello E. Chiodini, Roberta Boselli, Eren Taskin, Edoardo Puglisi, Chiara Bertora, Lorenzo Poggianella, Stefano Amaducci, and et al. 2021. "Reducing N Fertilization without Yield Penalties in Maize with a Commercially Available Seed Dressing" Agronomy 11, no. 3: 407. https://doi.org/10.3390/agronomy11030407
APA StyleMaris, S. C., Capra, F., Ardenti, F., Chiodini, M. E., Boselli, R., Taskin, E., Puglisi, E., Bertora, C., Poggianella, L., Amaducci, S., Tabaglio, V., & Fiorini, A. (2021). Reducing N Fertilization without Yield Penalties in Maize with a Commercially Available Seed Dressing. Agronomy, 11(3), 407. https://doi.org/10.3390/agronomy11030407