
Pollen Flow of Winter Triticale (x Triticosecale Wittmack) Investigated with Transgenic Line Expressing β-Glucuronidase Gene




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
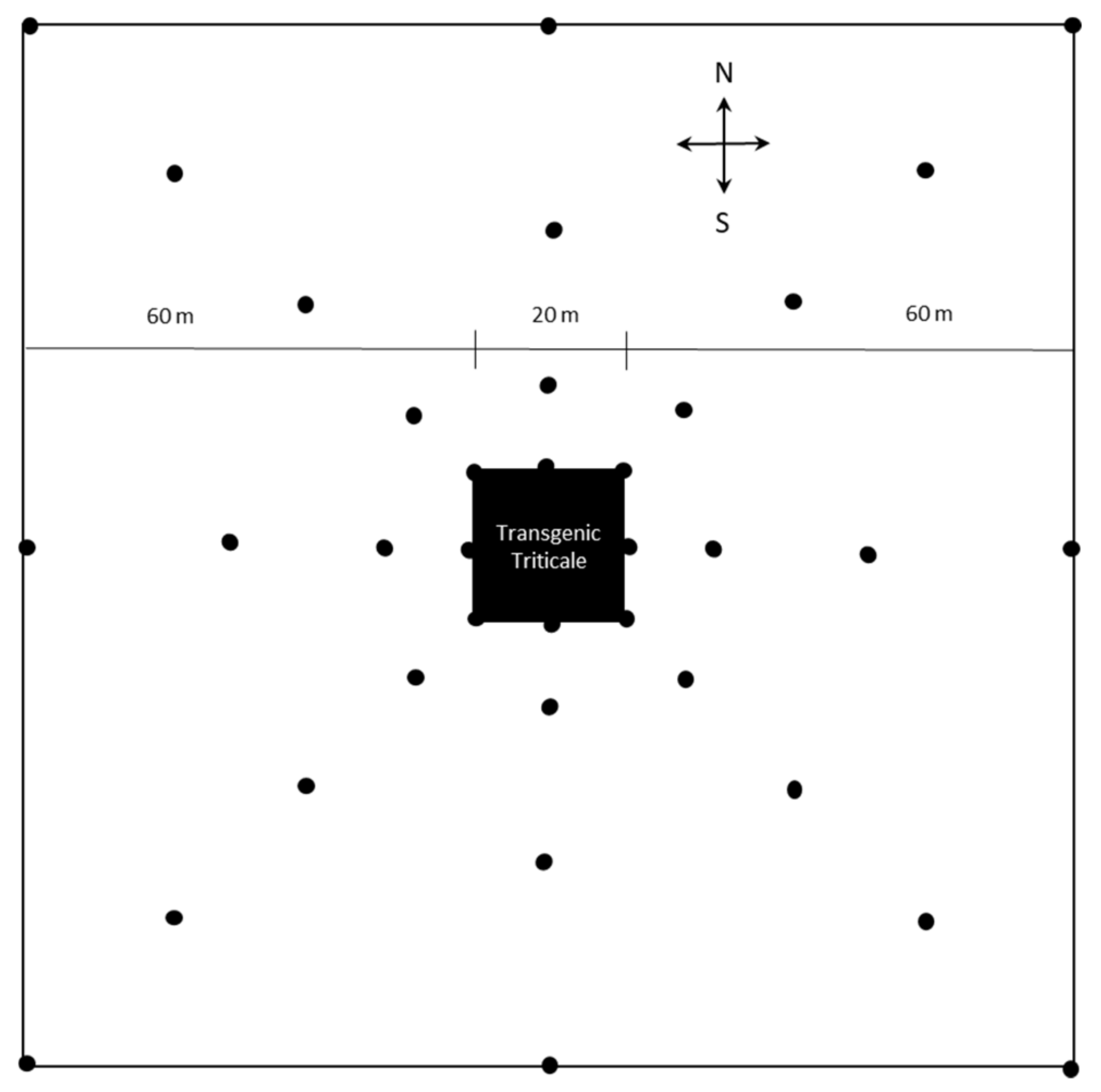
2. Materials and Methods
3. Results
3.1. Weather during the Anthesis Phase
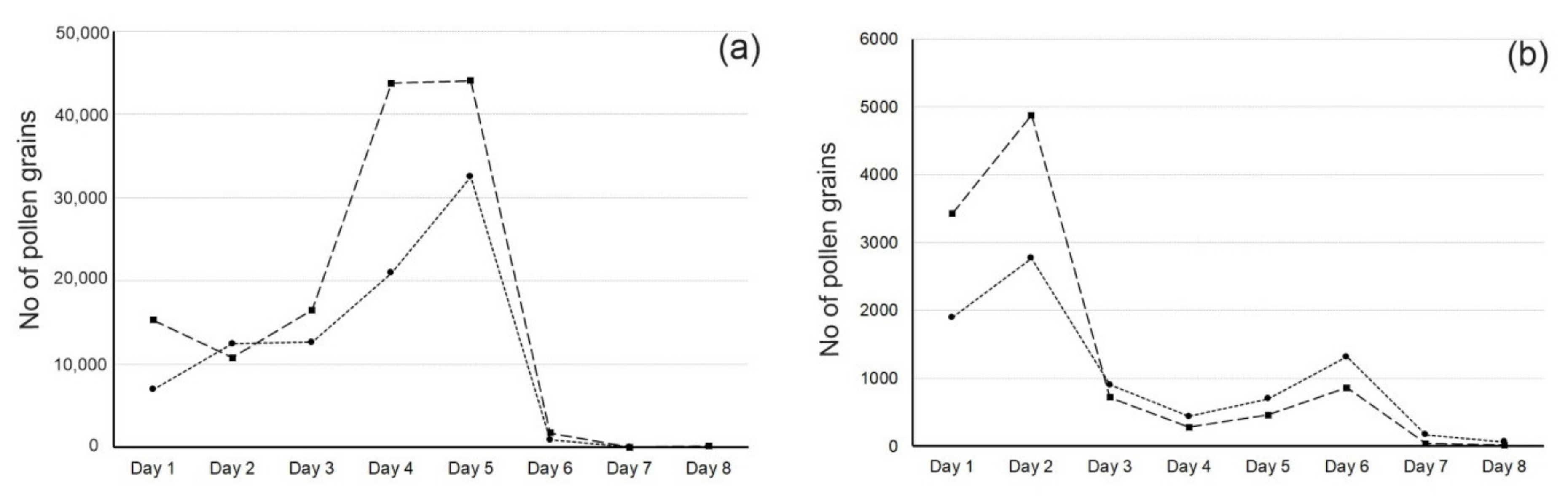
3.2. Pollen Flow
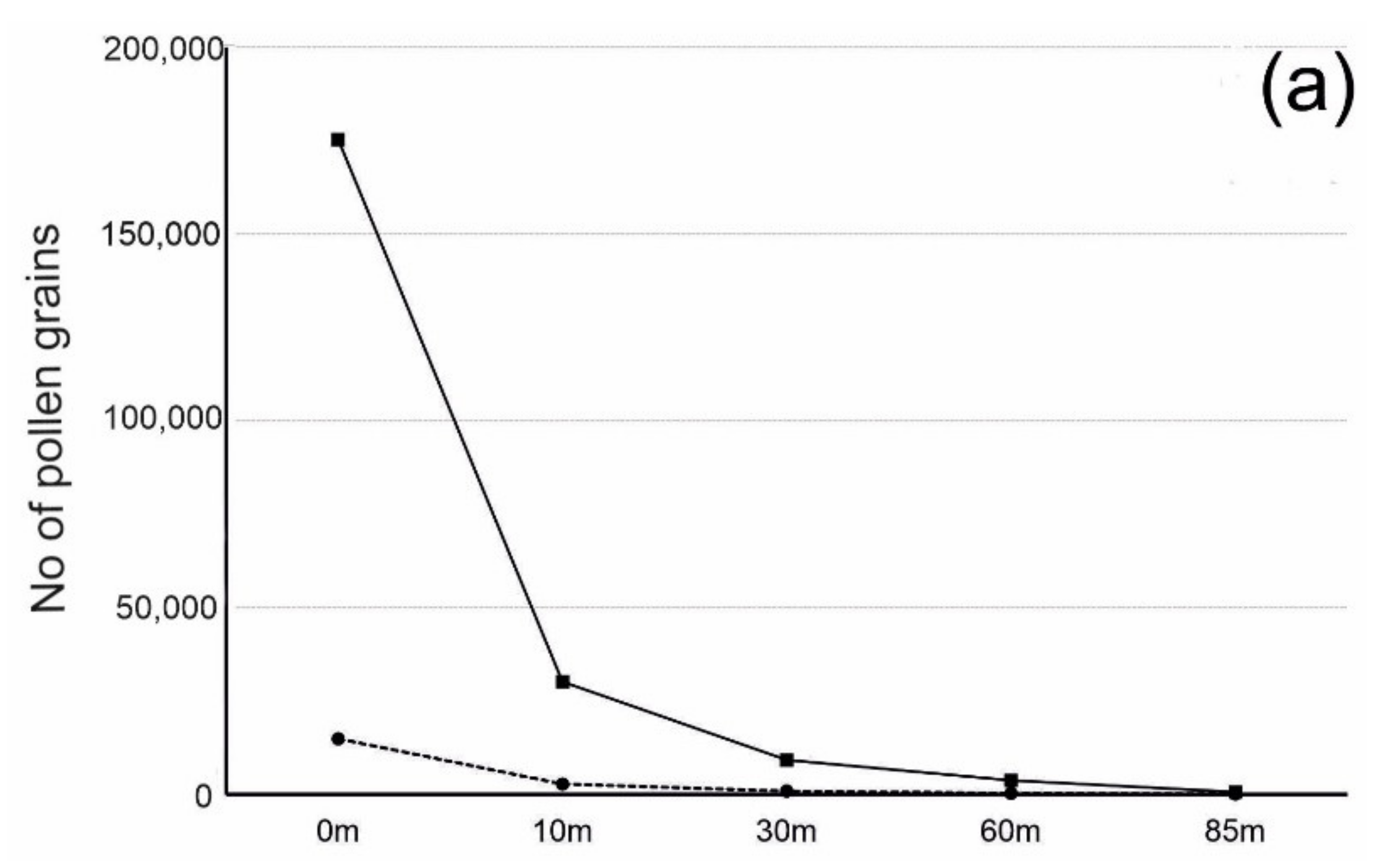
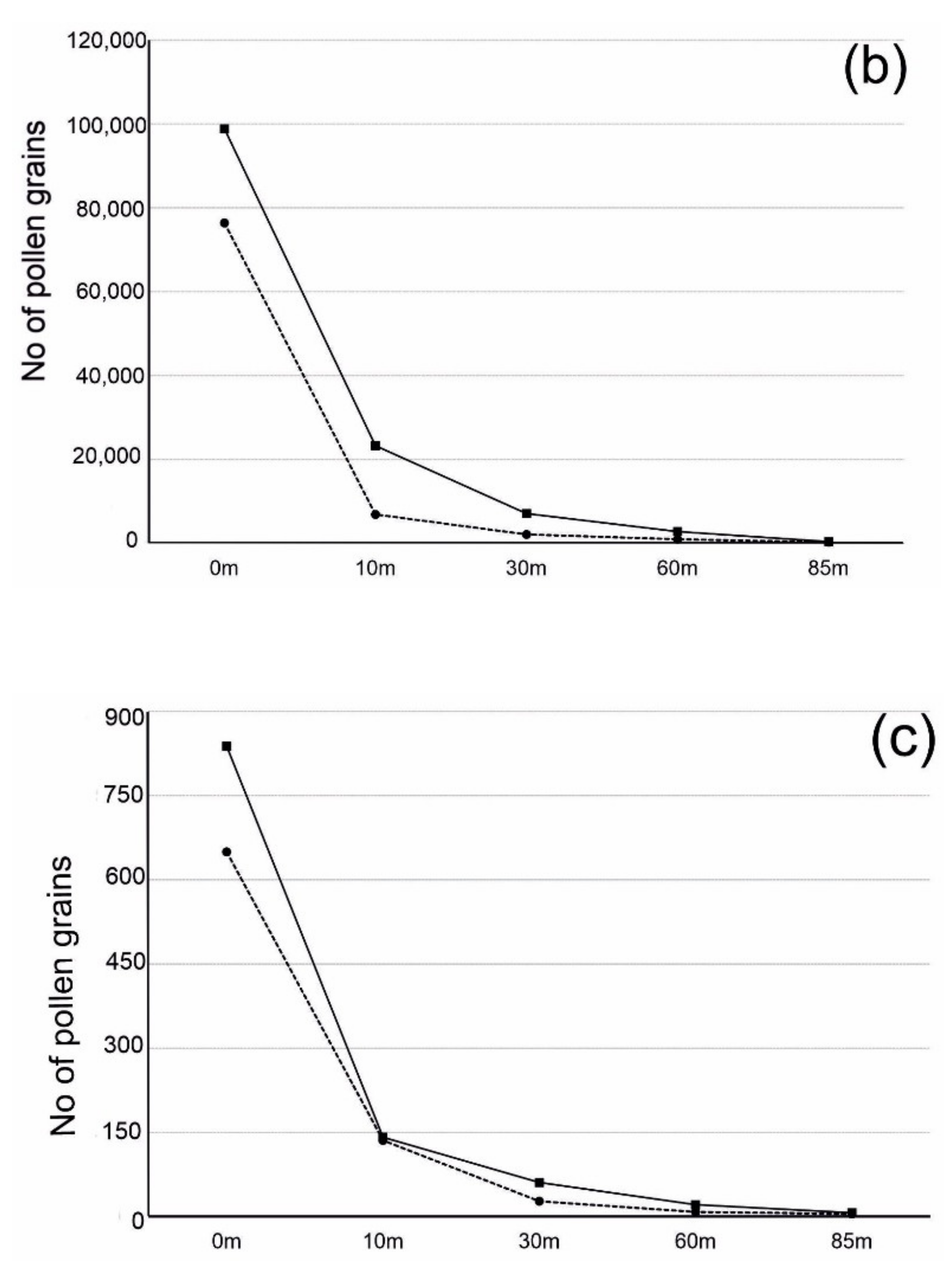
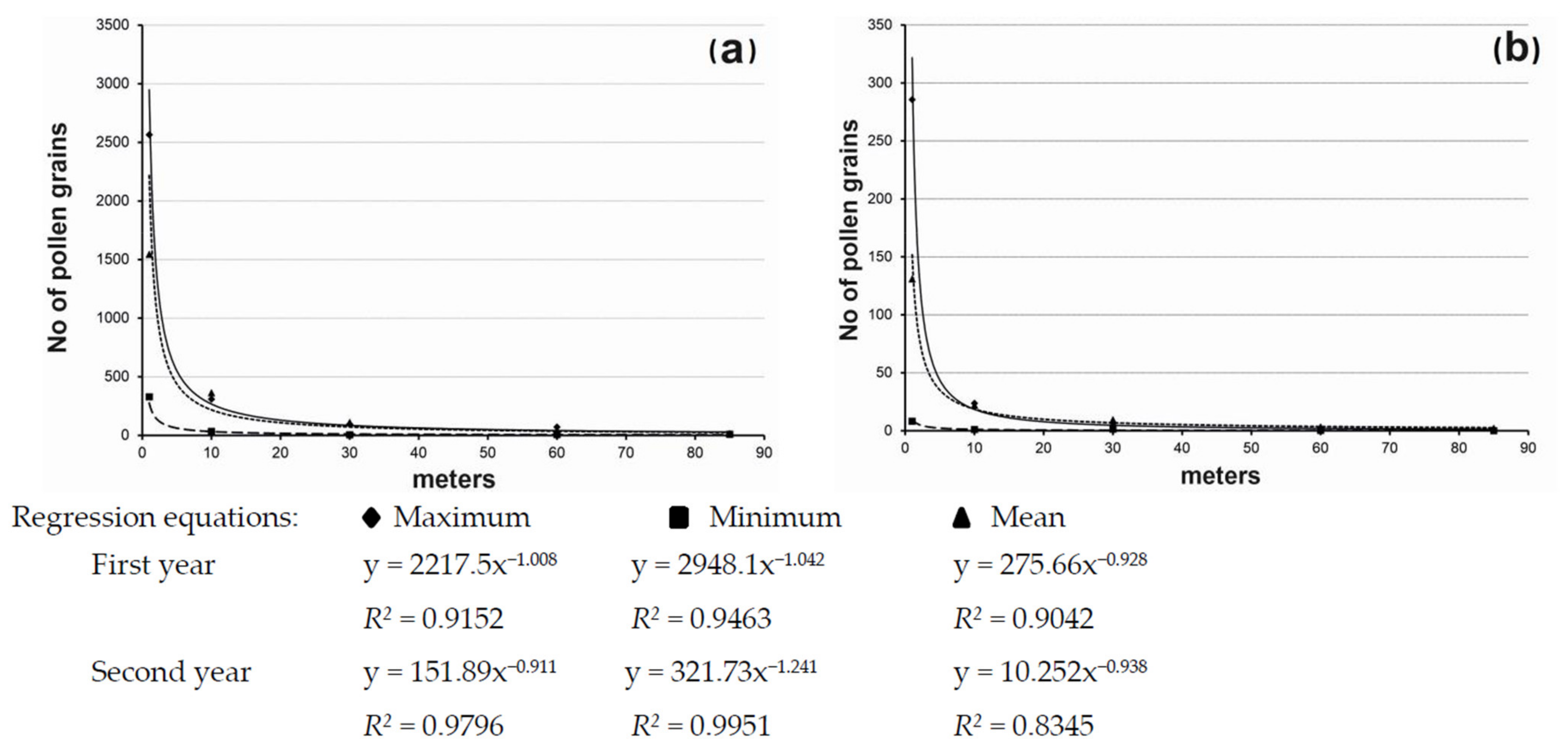
3.3. Number of Pollen Grains and Distance from the Source
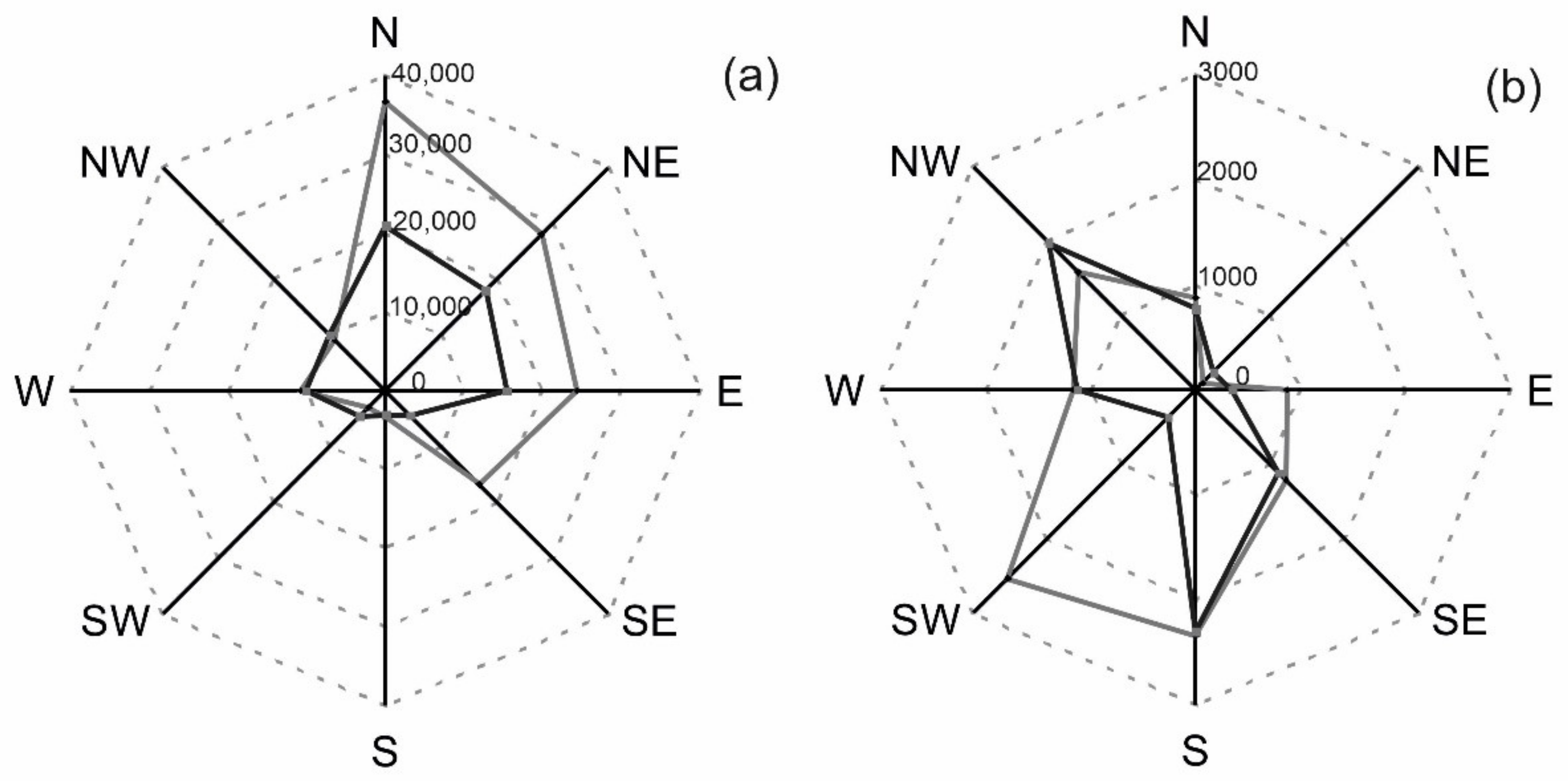
3.4. Number of Pollen Grains and Geographical Directions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Zhang, X.; Chen, X.; Romeis, J.; Yin, X.; Peng, Y. Consumption of Bt rice pollen containing Cry1C or Cry2A does not pose a risk to Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Sci. Rep. 2015, 5, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, D.; Fang, Z.; Yang, L. Effects of Bt cabbage pollen on the honeybee Apis mellifera L. Sci. Rep. 2018, 8, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzyk, I.; Kaszewski, B.M.; Weryszko-Chmielewska, E.; Nowak, M.; Sulborska, A.; Kaczmarek, J.; Szymanska, A.; Haratym, W.; Jedryczka, M. Warm and dry weather accelerates and elongates Cladosporium spore seasons in Poland. Aerobiologia 2016, 32, 109–129. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Hovmoller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.S. Plant pathogen dispersal. In eLS; John Wiley & Sons Chichester: West Sussex, UK, 2014. [Google Scholar]
- Weryszko-Chmielewska, E.; Kasprzyk, I.; Nowak, M.; Sulborska, M.; Kaczmarek, J.; Szymanska, A.; Haratym, W.; Gilski, M.; Jedryczka, M. Health hazards related to conidia of Cladosporium—Biological air pollutants in Poland, central Europe. J. Environ. Sci. 2018, 65, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Jedryczka, M.; Strzelczak, A.; Grinn-Gofron, A.; Nowak, M.; Wolski, T.; Siwulski, M.; Sobieralski, K.; Kaczmarek, J. Advanced statistical models commonly applied in aerobiology cannot accurately predict the exposure of people to Ganoderma spore-related allergies. Agric. For. Meteorol. 2015, 201, 209–217. [Google Scholar] [CrossRef]
- Van Ginkel, M.; Flipphi, R.C.H. Why self-fertilizing plants still exist in wild populations: Diversity assurance through stress-induced male sterility may promote selective outcrossing and recombination. Agronomy 2020, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wei, W.; Mab, K.; Lia, J.; Liangd, Y.; Darmenc, H. Consequences of gene flow between oilseed rape (Brassica napus) and its relatives. Plant Sci. 2013, 211, 42–51. [Google Scholar] [CrossRef]
- Eastham, K.; Sweet, J. Genetically Modified Organisms (GMOs): The Significance of Gene Flow through Pollen Transfer; Environmental Issue Report No 28; European Environment Agency: Copenhagen, Denmark, 2002; pp. 1–75. [Google Scholar]
- Coetzee, K. Evaluation of the Crossability between Small Grains. MSc Thesis, University of Stellenbosch, South Africa, 2011. [Google Scholar]
- Kavanagh, V.B.; Hills, M.J.; Eudes, F.; Topinka, K.; Yang, R.-C.; Hall, L.M. Pollen-mediated gene flow in triticale. Crop Sci. 2012, 52, 2293–2303. [Google Scholar] [CrossRef]
- Kavanagh, V.B.; Hall, L.M.; Hall, J.C. Potential hybridization of genetically engineered triticale with wild and weedy relatives in Canada. Crop Sci. 2010, 50, 1128–1140. [Google Scholar] [CrossRef]
- Kociuba, W.; Skulimowska, R. Observation of flowering, pollination and fertility of triticale plants in comparison to wheat and rye. Biul. Inst. Hod. Aklim. Roślin 1995, 195/196, 99–106, (in Polish with English abstract). [Google Scholar]
- Sowa, W.; Krysiak, H. Outcrossing in winter triticale, measured by occurrence of tall plants. In Triticale: Today and Tomorrow Developments in Plant, Breeding; Guedes-Pinto, H., Darvey, N., Carnide, V.P., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 593–596. [Google Scholar]
- De Vries, A.P. Some aspects of cross-pollination in wheat (Triticum aestivum L.) Pollen concentration in the field as influenced by variety, diurnal pattern, weather conditions and levels as compared to the height of the pollen donor. Euphytica 1972, 21, 185–203. [Google Scholar] [CrossRef]
- Virmani, S.S.; Edwards, I.B. Factors affecting cross-fertilization. In: Current status and future prospects for breeding hybrid rice and wheat. Adv. Agron. 1983, 36, 183–190. [Google Scholar]
- Waines, J.G.; Hegde, S.G. Intraspecific gene flow in bread wheat as affected by reproductive biology and pollination ecology of wheat flowers. Crop Sci. 2003, 43, 451–463. [Google Scholar] [CrossRef]
- Hills, M.J.; Hall, L.M.; Messenger, D.F.; Graf, R.J.; Beres, B.L.; Eudes, F. Evaluation of crossability between triticale (X Triticosecale Wittmack) and common wheat, durum wheat and rye. Environ. Biosaf. Res. 2007, 6, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, V.B.; Hills, M.J.; Goyal, A.; Randhawa, H.S.; Topinka, A.K.; Eudes, F.; Hall, L.M. Molecular markers as a complementary tool in risk assessment: Quantifying interspecific gene flow from triticale to spring wheat and durum wheat. Transgenic Res. 2013, 22, 767–778. [Google Scholar] [CrossRef]
- Kuparinen, A.; Katul, G.; Nathan, R.; Schurr, F.M. Increases in air temperature can promote wind-driven dispersal and spread of plants. Proc. R. Soc. B. 2009, 276, 3081–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, K.C.; Larter, E.N. Pollen production and disseminating properties of triticale relative to wheat. Can. J. Plant Sci. 1972, 52, 569–574. [Google Scholar] [CrossRef]
- Zimny, J.; Becker, D.; Brettschneider, R.; Lörz, H. Fertile transgenic Triticale (x Triticosecale Wittmack). Mol. Breed. 1995, 1, 155–164. [Google Scholar] [CrossRef]
- Baranowski, A.; Rosochacki, S.; Parada, R.; Jaszczak, K.; Zimny, J.; Połoszynowicz, J. The effect of diet containing genetically modified triticale on growth and transgenic DNA fate in selected tissues of mice. Anim. Sci. Pap. Rep. 2006, 24, 129–142. [Google Scholar]
- Jaszczak, K.; Kruszewski, M.; Baranowski, A.; Parada, R.; Bartłomiejczyk, T.; Zimny, J.; Rosochacki, S. Micronucleus test and comet assay on mice fed over five generations a diet containing genetically modified triticale. J. Anim. Feed Sci. 2008, 17, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Krzyżowska, M.; Wincenciak, M.; Winnicka, A.; Baranowski, A.; Jaszczak, K.; Zimny, J.; Niemiałtowski, M. The effect of multigenerational diet containing genetically modified triticale on immune system in mice. Pol. J. Vet. Sci. 2010, 13, 423–430. [Google Scholar]
- Becker, D.; Brettschneider, R.; Lörz, H. Fertile transgenic wheat from microprojectile bombardment of scutellar tissue. Plant J. 1994, 5, 299–307. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Rabiza-Swider, J.; Zimny, J.; Lukaszewski, A.J. Aneuploidy among androgenic progeny of hexaploid triticale (X Triticosecale Wittmack). Plant Cell Rep. 2011, 30, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Lacey, M.; West, J.S. The Air Spora: A Manual for Catching and Identifying Airborne Biological Particles; Springer GmbH: New York, NY, USA, 2006; pp. 1–156. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Kubáček, L. On a linearization of regression models. Appl. Math. 1995, 40, 61–78. [Google Scholar] [CrossRef]
- Rao, C.R. Linears Statistical Inference and Its Applications, 2nd ed.; J. Wiley: New York, NY, USA, 1973. [Google Scholar]
- Rawlings, J.O.; Pantula, S.G.; Dickey, D.A. Applied Regression Analysis—A Research Tool, 2nd ed.; Springer Inc: New York, NY, USA, 2001. [Google Scholar]
- Gatford, K.T.; Basri, Z.; Edlington, J.; Lloyd, J.; Qureshi, J.A.; Brettell, R.; Fincher, G.B. Gene flow from transgenic wheat and barley under field conditions. Euphytica 2006, 151, 383–391. [Google Scholar] [CrossRef]
- Hucl, P. Outcrossing in early-stage spring wheat breeder seed development. Crop Sci. 2010, 50, 29–34. [Google Scholar] [CrossRef]
- Sapra, V.T.; Hughes, J.L. Pollen production in hexaploid triticale. Euphytica 1975, 24, 237–243. [Google Scholar] [CrossRef]
- Komaki, M.K.; Tsunewaki, K. Genetical studies on the difference of anther length among common wheat cultivars. Euphytica 1981, 30, 45–53. [Google Scholar] [CrossRef]
- Singh, S.K.; Arun, B.; Joshi, A.K. Comparative evaluation of exotic and adapted germplasm of spring wheat in the Indo-Gangetic Plains of northern India. Plant Breed. 2007, 126, 559–564. [Google Scholar] [CrossRef]
- Athwal, R.S.; Kimber, G. Anther size and pollen longevity in wheat/rye addition lines. Wheat Inf. Serv. 1970, 30, 30–32. [Google Scholar]
- Nguyen, V.; Fleury, D.; Timmins, A.; Laga, H.; Hayden, M.; Mather, D.; Okada, T. Addition of rye chromosome 4R to wheat increases anther length and pollen grain number. Theor. Appl. Genet. 2015, 128, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Aylor, D.E. Settling speed of corn (Zea mays) pollen. J. Aerosol Sci. 2002, 33, 1601–1607. [Google Scholar] [CrossRef]
- Hanson, B.D.; Mallory-Smith, C.A.; Shaffi, B.; Thill, D.C.; Zemetra, R.S. Pollen-mediated gene flow from aleurone wheat to other wheat cultivars. Crop Sci. 2005, 45, 1610–1617. [Google Scholar] [CrossRef]
- Dong, S.; Liu, Y.; Yu, C.; Zhang, Z.; Chen, M.; Wang, C. Investigating Pollen and gene flow of WYMV-resistant transgenic wheat N12-1 using a dwarf male-sterile line as the pollen receptor. PLoS ONE 2016, 14, e0151373. [Google Scholar] [CrossRef]
- Loureiro, I.; Escorial, M.C.; González, Á.; Chueca, M.C. Pollen-mediated gene flow in wheat (Triticum aestivum L.) in a semiarid field environment in Spain. Transgenic Res. 2012, 21, 1329–1339. [Google Scholar] [CrossRef]
- Treu, R.; Emberlin, J. Pollen Dispersal in the Crops, Maize (Zea mays), Oil Seed Rape (Brassica napus ssp oleifera), Potatoes (Solanum tuberosum), Sugar Beet (Beta vulgaris ssp vulgaris) and Wheat (Triticum aestivum); Soil Association: Bristol, UK, 2000. [Google Scholar]
- Joo-Hee, A.; Kang-Hyun, C. Temporal and spatial characteristics in the pollen flow of living modified rice. Korean J. Crop Sci. 2009, 54, 210–217. [Google Scholar]
- Ritala, A.; Nuutila, A.M.; Aikasalo, R.; Kauppinen, V.; Tammisola, J. Measuring gene flow in the cultivation of transgenic barley. Crop Sci. 2002, 42, 278–285. [Google Scholar] [CrossRef]
- Van De Wiel, C.C.M.; Groeneveld, R.; Dolstra, O.; Kok, E.J.; Scholtens, I.; Thissen, J.; Smulders, M.J.M.; Lotz, L. Pollen-mediated gene flow in maize tested for coexistence of GM and non-GM crops in the Netherlands: Effect of isolation distances. NJAS Wageningen. J. Life Sci. 2009, 56, 405–423. [Google Scholar]
- Loureiro, I.; Escorial, M.C.; Garcia-Baudin, J.M.; Chueca, M.C. Evidence of natural hybridization between Aegilops geniculata and wheat under field conditions in Central Spain. Environ. Biosaf. Res. 2006, 5, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Zhu, W.; Zhang, B.; Zhang, X.; Zhu, J.; Shi, J.; Wu, P.; Wu, F.; Li, X.; Zhang, Q.; et al. Pollen-mediated gene flow from transgenic cotton constrained by physical isolation measures. Sci. Rep. 2018, 8, 2862. [Google Scholar] [CrossRef]
- OECD Seed Schemes Rules and Regulations. 2021. Available online: https://www.oecd.org/agriculture/seeds/documents/oecd-seed-schemes-rules-and-regulations.pdf (accessed on 19 February 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimny, J.; Sowa, S.; Otręba, P.; Kozdój, J.; Zimny, A.; Kaczmarek, J.; Oleszczuk, S.; Czaplicki, A.; Jędryczka, M. Pollen Flow of Winter Triticale (x Triticosecale Wittmack) Investigated with Transgenic Line Expressing β-Glucuronidase Gene. Agronomy 2021, 11, 431. https://doi.org/10.3390/agronomy11030431
Zimny J, Sowa S, Otręba P, Kozdój J, Zimny A, Kaczmarek J, Oleszczuk S, Czaplicki A, Jędryczka M. Pollen Flow of Winter Triticale (x Triticosecale Wittmack) Investigated with Transgenic Line Expressing β-Glucuronidase Gene. Agronomy. 2021; 11(3):431. https://doi.org/10.3390/agronomy11030431
Chicago/Turabian StyleZimny, Janusz, Sławomir Sowa, Piotr Otręba, Janusz Kozdój, Aleksandra Zimny, Joanna Kaczmarek, Sylwia Oleszczuk, Andrzej Czaplicki, and Małgorzata Jędryczka. 2021. "Pollen Flow of Winter Triticale (x Triticosecale Wittmack) Investigated with Transgenic Line Expressing β-Glucuronidase Gene" Agronomy 11, no. 3: 431. https://doi.org/10.3390/agronomy11030431
APA StyleZimny, J., Sowa, S., Otręba, P., Kozdój, J., Zimny, A., Kaczmarek, J., Oleszczuk, S., Czaplicki, A., & Jędryczka, M. (2021). Pollen Flow of Winter Triticale (x Triticosecale Wittmack) Investigated with Transgenic Line Expressing β-Glucuronidase Gene. Agronomy, 11(3), 431. https://doi.org/10.3390/agronomy11030431