
The Morphological Responses of Calendula officinalis L. “Radio” to the Foliar Application of Benzyladenine and Different Light Spectra




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Treatment of Cytokines and Light
2.3. Plant Measurements and Experimental Design
2.4. Model Construction
3. Results
3.1. The Earliness of Florescence
3.2. Biometric Parameters
3.3. Relative Content of Chlorophyll, Fluorescence, Dry Mass
3.4. The Morphological Parameters in the 8th, 10th, 12th and 14th Week of Pot Marigold Growth
4. Discussion
4.1. Flowering
4.2. Growth
4.3. BA
4.4. Dry Matter
4.5. Chlorophyll
4.6. Fv/Fm
4.7. Daily Light Integral (DLI)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohammad, M.S.; Kashani, H.H. Pot marigold (Calendula officinalis) medicinal usage and cultivation. Sci. Res. Essays 2012, 7, 1468–1472. [Google Scholar] [CrossRef]
- Van Wyk, B.E.; Wink, M. Medicinal Plants of the World; Briza Publications: Pretoria, South Africa, 2004. [Google Scholar]
- Gazim, Z.; Rezende, C.; Fraga, S.; Dias Filho, B.; Nakamura, C.; Cortez, D. Analysis of the essential oils from Calendula officinalis growing in Brazil using three different extraction procedures. Braz. J. Pharm. 2008, 44, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Klimek-Szczykutowicz, M.; Jafernik, K.; Koc, K.; Ekiert, H. Pot marigold (Calendula officinalis L.)—A position in classical phytotherapy and newly documented activities. Acta Scientiarum Polonorum Hortorum Cultus 2020, 19, 47–61. [Google Scholar] [CrossRef]
- Pogroszewska, E.; Sadkowska, P. The effect of benzyladenine on the flowering of Campanula persicifolia L. ‘Alba’ cultivated in an unheated plastic tunnel and in the field. Acta Agrobotanica 2008, 7, 57–63. [Google Scholar]
- Pobudkiewicz, A.K.; Nowak, J. The influence of florprimidol and uniconazole on growth of the CMM draft Dianthus caryophyllus L. ‘Snowmass’. J. Fruit Ornam. Plant Res. 1994, 2, 135–142. [Google Scholar]
- James, L.; Gibson, B.; Whipker, E. The effect of B9 + cycocel on the growth of Brassica juncea var. ‘Rugosa Red Giant’. SNA Res. Conf. 1999, 4, 284–287. [Google Scholar]
- Al-Khassawneh, N.M.; Karam, N.S.; Shibli, R.A. Growth and flowering of black iris (Iris nigricans Dinsm.) following treatment with plant growth regulators. Sci. Hort. 2006, 107, 187–193. [Google Scholar] [CrossRef]
- Magnitskiy, S.V.; Pasian, C.C.; Bennett, M.A.; Metzger, J.D. Controlling plug height of verbena, celosia, and pansy by treating seeds with paclobutrazol. Hort. Sci. 2006, 47, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Kieber, J.J.; Schaller, G.E. Cytokinins. In Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2014; Volume 12, p. e0168. [Google Scholar] [CrossRef] [Green Version]
- Dobisova, T.; Hrdinova, V.; Cuesta, C.; Michlickova, S.; Urbankova, I.; Hejatkova, R.; Zadnikova, P.; Pernisova, M.; Benkova, E.; Hejatko, J. Light controls cytokinin signaling via transcriptional regulation of constitutively active sensor histidine kinase CKI1. Plant Physiol. 2017, 174, 387–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janowska, B. Effect of growth regulators on flower and leaf yield of the calla lily (Zantedeschia Spreng.). Hortic. Sci. 2013, 40, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Gislerød, H.R.; Torre, S.; Olsen, J.E. Control of shoot elongation and hormone physiology in poinsettia by light quality provided by light emitting diodes—A minireview. Acta Horticulturae 2015, 1104, 131–136. [Google Scholar] [CrossRef]
- Bergstrand, K.J.I. Methods for growth regulation of greenhouse produced ornamental pot- and bedding plants—A current review. Folia Horticulturae 2017, 29, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, H.; Ma, Z.; Tazawa, S.; Douzono, M.; Runkle, E.S.; Heins, R.D. Blue light inhibits stem elongation of chrysanthemum. Acta Horticulturae 2006, 711, 363–368. [Google Scholar] [CrossRef]
- Ouzounis, T.; Frette, X.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of supplementary lighting on the secondary metabolities in roses, chrysanthemum and campanulas. J. Plant Physiol. 2014, 171, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Frąszczak, B.; Gąsecka, M.; Golcz, A.; Zawirska-Wojtasiak, R. The effect of radiation of LED modules on the growth of dill (Anethum graveolens L.). Open Life Sci. 2016, 11, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED lighting in horticulture. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 113–147. [Google Scholar]
- Ibaraki, Y. LED Supplementary Lighting. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 27–36. [Google Scholar]
- Park, Y.; Runkle, E.S. Blue radiation attenuates the effects of the red to far-red ratio on extension growth but not on flowering. Environ. Exp. Bot. 2019, 168, 103871. [Google Scholar] [CrossRef]
- Sager, J.C.; Smith, W.O.; Edwards, J.L.; Cyr, K.L. Photosynthetic efficiency and phytochrome photoequilibria determination using special data. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Botany 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Fruit Products and Vegetable. The Preparation of Samples and Method of Physic Chemist Investigations. Marking the Content of Dry Mass with Weight Method; PN-90/A-75101/03; PKN: Warsaw, Poland, 1990. (In Polish)
- Baloch, J.U.D.; Munir, M. Effects of supplementary irradiance on flowering time of obligate long day ornamental annuals under non-inductive environment. Pak. J. Bot. 2014, 46, 1253–1259. [Google Scholar]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of flowering time by Arabidopsis photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Mockler, T.C.; Guo, H.; Yang, H.; Duong, H.; Lin, C. Antagonistic actions of Arabidopsis cryptochromes and phytochrome B in the regulation of floral induction. Development 1999, 126, 2073–2082. [Google Scholar]
- Mockler, T.; Yang, H.; Yu, X.; Parikh, D.; Cheng, Y.C.; Dolan, S.; Lin, C. Regulation of photoperiodic flowering by Arabidopsis photoreceptors. PNAS 2003, 100, 2140–2145. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Runkle, E.S. Moderate intensity blue radiation can regulate flowering, but not extension growth, of several photoperiodic ornamental crops. Environ. Exp. Bot. 2017, 134, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Cope, K.R.; Bugbee, B. Spectral effects of three types of white light-emitting diodes on plant growth and development: Absolute versus relative amounts of blue light. Hort. Sci. 2013, 48, 504–509. [Google Scholar] [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. Hort. Sci. 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- King, R.W.; Hisamatsu, T.; Gildschmidt, E.E.; Blundell, C. The nature of floral signals in Arabidopsis. I. photosynthesis and a far-red photoresponse independently regulate flowering by increasing expression of FLOWERING LOCUS T (FT). J. Exp. Bot. 2008, 59, 3811–3820. [Google Scholar] [CrossRef] [Green Version]
- Owen, W.G.; Meng, Q.; Lopez, R.G. Promotion of flowering from far-red radiation depends on the photosynthetic daily light integral. Hort. Sci. 2018, 53, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Wollaeger, H.M.; Runkle, E.S. Growth and acclimation of impatiens, salvia, petunia, and tomato seedlings to blue and red light. Hort. Sci. 2015, 50, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Frąszczak, B. The effect of different doses of blue light on the biometric traits and photosynthesis of dill plants. Not. Bot. Horti. Agrobot. Cluj Napoca 2016, 44, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Lin, C. Plant blue-light receptors. Trends Plant Sci. 2000, 5, 337–342. [Google Scholar] [CrossRef]
- Banerjee, R.; Batschauer, A. Plant blue-light receptors. Planta 2005, 20, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Kuwar, G.; Clarke, J.L.; Blystad, D.R.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Sci. Hortic. 2012, 147, 136–143. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Heydarizadeh, P.; Zahedi, M.; Boroomand, A.; Agharokh, M.; Sahba, M.R.; Schoefs, B. High performance of vegetables, flowers and medicinal plants in a red-blue LED incubator for indoor plant production. Agron. Sustain. Dev. 2014, 34, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic performance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar] [CrossRef]
- Schouten, R.E.; van Kooten, O.; Jalink, H.; Kappers, L.F.; Snel, J.F.H.; Jordi, W. Genetic and physiological factors affecting colour and firmness. In Understanding and Measuring the Shelf-Life of Food; Steel, R., Ed.; Woodhead Publishing: Cambridge, UK, 2004; pp. 69–90. [Google Scholar]
- Schwend, T.; Prucker, D.; Mempel, H. Red light promotes compact growth of sunflowers. Eur. J. Hortic. Sci. 2015, 80, 56–61. [Google Scholar] [CrossRef]
- Nanya, K.; Ishigami, Y.; Hikosaka, S.; Goto, E. Effects of blue and red light on stem elongation and flowering of tomato seedlings. Acta Horticulturae 2012, 956, 261–266. [Google Scholar] [CrossRef]
- Kazemi, S.S.; Hashemabadi, D.; Torkashvand, A.M.; Kaviani, B. Effect of cycocel and daminozide on vegetative growth, flowering and the content of essence of pot marigold (Calendula officinalis). J. Ornam. Plants 2014, 4, 107–114. [Google Scholar]
- Fukuda, N.; Yoshida, T.; Olsen, J.E.; Senaha, C.; Jikumaru, Y.; Kamiya, Y. Short main shoot length and inhibition of floral bud development under red light condition can be recovered by application of gibberellin and cytokinin. Acta Horticulturae 2012, 956, 215–222. [Google Scholar] [CrossRef]
- Hashemabadi, D.; Lipaei, S.R.; Shadparvar, V.; Zarchini, M.; Kaviani, B. The effect of cycocel and daminozide on some growth and flowering characteristics of Calendula officinalis L., an ornamental and medicinal plant. J. Med. Plant. Res. 2012, 6, 1752–1757. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Habricot, Y.; Condiff, A.S.; Maldiney, R.; van der Straeten, D.; Ahmad, M. HY5 is a point of convergence between cryptochrome and cytokinin signalling pathways in Arabidopsis thaliana. Plant J. 2007, 49, 428–444. [Google Scholar] [CrossRef]
- Cohen, L.; Gepstein, S.; Horwitz, B.A. Similarity between cytokinin and blue light inhibition of cucumber hypocotyl elongation. Plant Physiol. 1991, 95, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Howell, S.H. The effects of cytokinin and light on hypocotyl elongation in Arabidopsis seedlings are independent and additive. Plant Physiol. 1995, 108, 1423–1430. [Google Scholar] [CrossRef] [Green Version]
- Janowska, B.; Stanecki, M. Effect of rhizome soaking in a mixture of BA and GA3 on the earliness of flowering and quality of the yield of flowers and leaves in the calla lily (Zantedeschia Spreng.). Acta Scientiarum Polonorum Hortorum Cultus 2013, 12, 3–12. [Google Scholar]
- Mahmood, S.; Reza, R.; Hossain, G.; Hauser, B. Response of cytokinins on in vitro shoot multiplication of rose cv. Frisco. Res. Rev. 2017, 5, 8–12. [Google Scholar] [CrossRef]
- Cioć, M.; Kalisz, A.; Żupnik, M.; Pawłowska, B. Different LED light intensities and 6-BA concentrations in relation to shoot development, leaf architecture, and photosynthetic pigments of Gerbera jamesonii Bolus in vitro. Agronomy 2019, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Cope, K.; Snowden, M.C.; Bugbee, B. Photobiological interactions of blue light and photosynthetic photon flux: Effects of monochromatic and broad-spectrum light sources. Photochem. Photobiol. 2014, 90, 574–584. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Q.; Tong, Y.; Lu, W. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Pilarska, M.; Skowron, E.; Niewiadomska, E. Cytokinins and photosynthesis. Postępy Biochemii 2015, 61, 61–68. (In Polish) [Google Scholar] [PubMed]
- Aremu, A.O.; Bairu, M.W.; Szüčová, L.; Finnie, J.F.; van Staden, J. The role of meta-topolins on the photosynthetic pigment profiles and foliar structures of micropropagated ‘Williams’ bananas. J. Plant Physiol. 2012, 169, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Dobránszki, J.; Mendler-Drienyovszki, N. Cytokinin-induced changes in the chlorophyll content and fluorescence of in vitro apple leaves. J. Plant Physiol. 2014, 171, 1472–1478. [Google Scholar] [CrossRef]
- Bolhàr-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Öquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Borowski, E.; Michałek, S.; Rubinowska, K.; Hawrylak-Nowak, B.; Grudziński, W. The effects of light quality on photosynthetic parameters and yield of lettuce plants. Acta Scientiarum Polonorum Hortorum Cultus 2005, 14, 177–188. [Google Scholar]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (Nicotiana tabacum L.) plants. Photosynthetica 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Oh, W.; Cheon, I.H.; Kim, K.S.; Runkle, E.S. Photosynthetic daily light integral influences flowering time and crop characteristics of Cyclamen persicum. Hort. Sci. 2009, 44, 341–344. [Google Scholar] [CrossRef] [Green Version]
- Faust, J.E.; Holcombe, V.; Rajapakse, N.G.; Layne, D.R. The effect of daily light integral on bedding plant growth and flowering. Hort. Sci. 2005, 40, 645–649. [Google Scholar] [CrossRef] [Green Version]



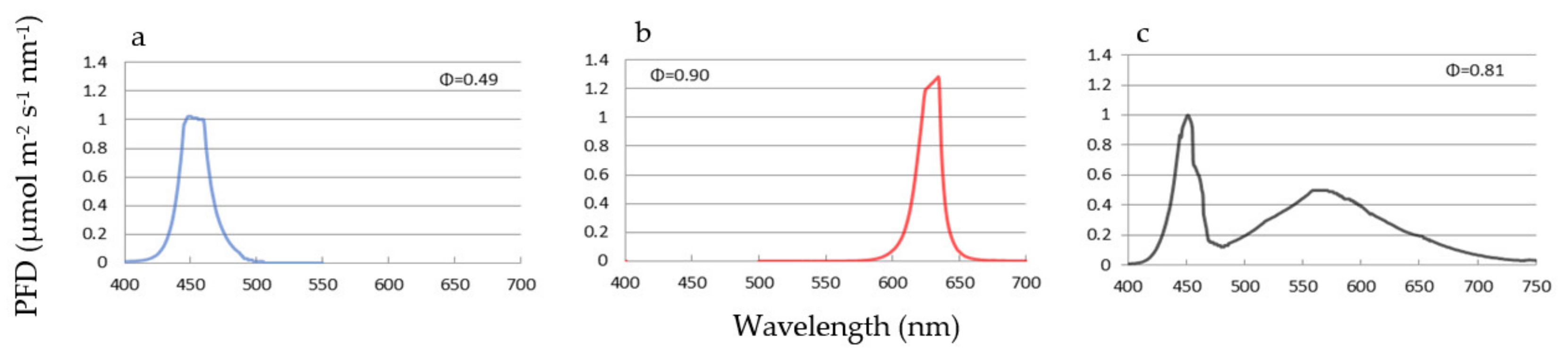{kind=link}
{kind=link}
| Light Colour | Wavelength (nm) | PFD * (μmol m−2 s−1) | % for W Light | % for W+R | % for W+B |
|---|---|---|---|---|---|
| UV | 320–380 | 0.5 | 0.3 | 0.2 | 0.2 |
| Violet | 380–450 | 15.4 | 8.9 | 6.6 | 6.6 |
| Blue | 450-495 | 30.3 | 17.6 | 13.0 | 38.9 |
| Green | 495–570 | 53.5 | 31.1 | 23.0 | 23.0 |
| Yellow | 570–590 | 18.7 | 10.9 | 8.1 | 8.1 |
| Orange | 590–620 | 21.8 | 12.7 | 9.4 | 9.4 |
| Red (R) | 620–700 | 26.4 | 15.3 | 37.2 | 11.4 |
| Far Red (FR) | 700–780 | 5.6 | 3.3 | 2.4 | 2.4 |
| sum | 320–780 | 172.2 | 100 | 100 | 100 |
| R:FR | 4.7 | - | 15.5 | - |
| Light Colour | Benzyladenine (BA) Concentrations (mg dm−3) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 100 | 150 | 200 | Mean for Light | 0 | 100 | 150 | 200 | Mean for Light | 0 | 100 | 150 | 200 | Mean for Light | |
| first inflorescence | second inflorescence | third inflorescence | |||||||||||||
| Earliness of flowering (days) | |||||||||||||||
| W | 44.2 bc | 49.0 bc | 49.0 bc | 52.0 c | 48.5 b | 49.7 a | 56.8 bc | 75.1 cd | 74.5 cd | 64.0 b | 68.0 bc | 62.3 b | 86.8 de | 81.5 d | 74.7 b |
| W+R | 34.5 a | 42.2 ab | 48.7 bc | 46.7 bc | 43.0 a | 47.1 a | 54.1 bc | 56.9 bc | 56.4 bc | 52.5 a | 57.6 a | 61.7 b | 63.4 b | 65.3 bc | 62.0 a |
| W+B | 47.3 bc | 67.3 d | 63.0 d | 64.0 d | 60.4 c | 51.8 ab | 71.4 cd | 67.3 c | 81.4 d | 68.0 b | 68.2 bc | 80.3 d | 74.2 c | 90.0 e | 78.2 b |
| Mean for BA | 42.0 a | 52.8 b | 53.6 b | 54.2 b | 49.5 a | 60.8 b | 71.2 c | 70.8 c | 64.6 a | 68.1 ab | 74.8 b | 78.9 b | |||
| Number of inflorescences buds | |||||||||||||||
| W | 1.4 ab | 1.4 ab | 1.3 b | 0.6 b | 1.2 a | 1.0 ab | 0.8 ab | 1.0 ab | 0.0 b | 0.7 b | 0.4 cd | 0.0 d | 0.7 bd | 1.0 bd | 0.5 b |
| W+R | 0.9 b | 1.2 b | 2.2 a | 1.2 b | 1.4 a | 1.0 ab | 1.5 a | 1.5 a | 1.5 ab | 1.4 a | 3.0 a | 0.4 cd | 0.0 d | 0.0 d | 0.9 ab |
| W+B | 1.0 b | 1.3 b | 1.2 b | 0.5 b | 1.0 a | 0.0 b | 0.5 ab | 0.5 ab | 1.0 ab | 0.5 b | 1.7 b | 1.4 bc | 1.0 bd | 0.7 bd | 1.2 a |
| Mean for BA | 1.1 ab | 1.3 a | 1.6 a | 0.8 b | 0.7 a | 0.9 a | 1.0 a | 0.8 a | 1.7 a | 0.6 b | 0.6 b | 0.6 b | |||
| Diameter of inflorescence (cm) | |||||||||||||||
| W | 7.0 a | 5.6 c | 6.1 ac | 6.4 ac | 6.3 a | 5.8 bc | 5.4 bc | 4.9 c | 6.0 bc | 5.5 b | 5.7 b | 5.6 b | 4.4 c | 6.3 a | 5.5 a |
| W+R | 5.7 c | 5.9 c | 6.0 bc | 5.6 c | 5.8 b | 6.1 bc | 7.8 a | 5.5 bc | 6.5 b | 6.5 a | 6.4 a | 6.2 a | 5.0 bc | 5.6 b | 5.8 a |
| W+B | 6.9 ab | 6.3 ac | 5.8 c | 6.0 bc | 6.3 a | 5.0 bc | 6.2 bc | 5.9 bc | 5.9 bc | 5.8 b | 5.7 b | 5.3 bc | 5.8 b | 6.2 a | 5.8 a |
| Mean for BA | 6.5 a | 5.9 b | 6.0 b | 6.0 b | 5.6 b | 6.5 a | 5.4 b | 6.1 ab | 5.9 a | 5.7 a | 5.1 b | 6.0 a | |||
| Height (cm) | |||||||||||||||
| W | 26.4 bc | 26.3 bc | 23.5 cd | 24.5 cd | 25.2 b | 30.0 c | 27.0 cd | 26.7 cd | 26.3 cd | 27.5 b | 36.0 b | 29.4 cd | 33.0 c | 30.3 cd | 32.2 b |
| W+R | 28.4 b | 37.9 a | 37.0 a | 38.3 a | 35.4 a | 33.9 bc | 41.4 a | 39.0 ab | 41.0 a | 38.8 a | 34.0 c | 48.3 a | 41.3 ab | 43.0 ab | 41.7 a |
| W+B | 27.0 bc | 20.5 e | 20.7 e | 20.9 e | 22.3 c | 30.7 c | 23.0 d | 22.8 d | 32.7 bc | 27.3 b | 32.0 c | 26.9 d | 25.4 d | 33.3 c | 29.4 b |
| Mean for BA | 27.3 a | 28.2 a | 27.1 a | 27.9 a | 31.5 a | 30.5 a | 29.5 a | 33.3 a | 34.0 a | 34.9 a | 33.2 a | 35.5 a | |||
| Number of lateral shoots | |||||||||||||||
| W | 2.2 b | 2.3 b | 2.2 b | 1.8 b | 2.1 b | 2.8 c | 3.0 c | 2.8 c | 2.0 d | 2.7 b | 3.4 c | 3.0 c | 3.0 c | 3.0 c | 3.1 b |
| W+R | 1.8 b | 4.0 a | 4.5 a | 2.1 b | 3.1 a | 4.3 ab | 5.5 a | 4.7 ab | 4.0 ab | 4.6 a | 5.7 a | 6.4 a | 5.1 ab | 4.7 ab | 5.5 a |
| W+B | 1.9 b | 2.0 b | 1.9 b | 1.2 b | 1.8 b | 2.3 cd | 2.5 cd | 2.5 cd | 3.0 c | 2.6 b | 3.7 c | 3.0 c | 3.0 c | 3.4 c | 3.3 b |
| Mean for BA | 2.0 b | 2.8 a | 2.9 a | 1.7 b | 3.1 a | 3.7 a | 3.3 a | 3.0 a | 4.3 a | 4.1 a | 3.7 a | 3.7 a | |||
| Light Colour | Benzyloadenine (BA) Concentrations (mg dm−3) | Mean for Light | |||
|---|---|---|---|---|---|
| 0 | 100 | 150 | 200 | ||
| the relative content of chlorophyll | |||||
| W | 18.3 b | 16.5 c | 12.3 d | 12.9 d | 15.0 b |
| W+R | 22.0 a | 21.8 a | 18.9 b | 15.5 c | 19.6 a |
| W+B | 14.5 cd | 14.8 cd | 12.3 d | 12.2 d | 13.4 b |
| Mean for BA | 18.3 a | 17.7 a | 14.5 b | 13.5 b | |
| fluorescence Fv/Fm | |||||
| W | 0.805 cd | 0.815 ac | 0.809 cd | 0.814 bc | 0.811 b |
| W+R | 0.824 ab | 0.826 a | 0.823 ab | 0.826 a | 0.825 a |
| W+B | 0.801 d | 0.809 cd | 0.813 bc | 0.821 ab | 0.811 b |
| Mean for BA | 0.810 b | 0.817 a | 0.815 ab | 0.820 ab | |
| dry mass (g) | |||||
| W | 0.69 b | 0.60 bc | 0.61 bc | 0.55 c | 0.61 b |
| W+R | 0.86 a | 0.75 b | 0.89 a | 0.74 b | 0.81 a |
| W+B | 0.65 b | 8.7 0.59 c | 0.49 d | 0.57 c | 0.57 b |
| Mean for BA | 0.73 a | 0.64 b | 0.66 b | 0.62 b | |
| number of variables included: 5 | |||
| number of variables rejected: 0 | |||
| number of permutations: 9999 | |||
| E % | p | F | |
| NFB The number of flower buds | 11.59 | 0.001 | 19.72 |
| PH The plant height | 9.78 | 0.001 | 16.69 |
| NLS The number of lateral shoots | 9.22 | 0.001 | 16.21 |
| NF The number of inflorescences | 8.74 | 0.001 | 11.42 |
| DFH Diameter of flower heads | 6.47 | 0.001 | 8.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frąszczak, B.; Janowska, B.; Świerk, D. The Morphological Responses of Calendula officinalis L. “Radio” to the Foliar Application of Benzyladenine and Different Light Spectra. Agronomy 2021, 11, 460. https://doi.org/10.3390/agronomy11030460
Frąszczak B, Janowska B, Świerk D. The Morphological Responses of Calendula officinalis L. “Radio” to the Foliar Application of Benzyladenine and Different Light Spectra. Agronomy. 2021; 11(3):460. https://doi.org/10.3390/agronomy11030460
Chicago/Turabian StyleFrąszczak, Barbara, Beata Janowska, and Dariusz Świerk. 2021. "The Morphological Responses of Calendula officinalis L. “Radio” to the Foliar Application of Benzyladenine and Different Light Spectra" Agronomy 11, no. 3: 460. https://doi.org/10.3390/agronomy11030460
APA StyleFrąszczak, B., Janowska, B., & Świerk, D. (2021). The Morphological Responses of Calendula officinalis L. “Radio” to the Foliar Application of Benzyladenine and Different Light Spectra. Agronomy, 11(3), 460. https://doi.org/10.3390/agronomy11030460