The Effect of Nitrogen Reduction at Different Stages on Grain Yield and Nitrogen Use Efficiency for Nitrogen Efficient Rice Varieties



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Treatments
2.3. Sampling and Measurements
2.4. Statistical Analysis
3. Results
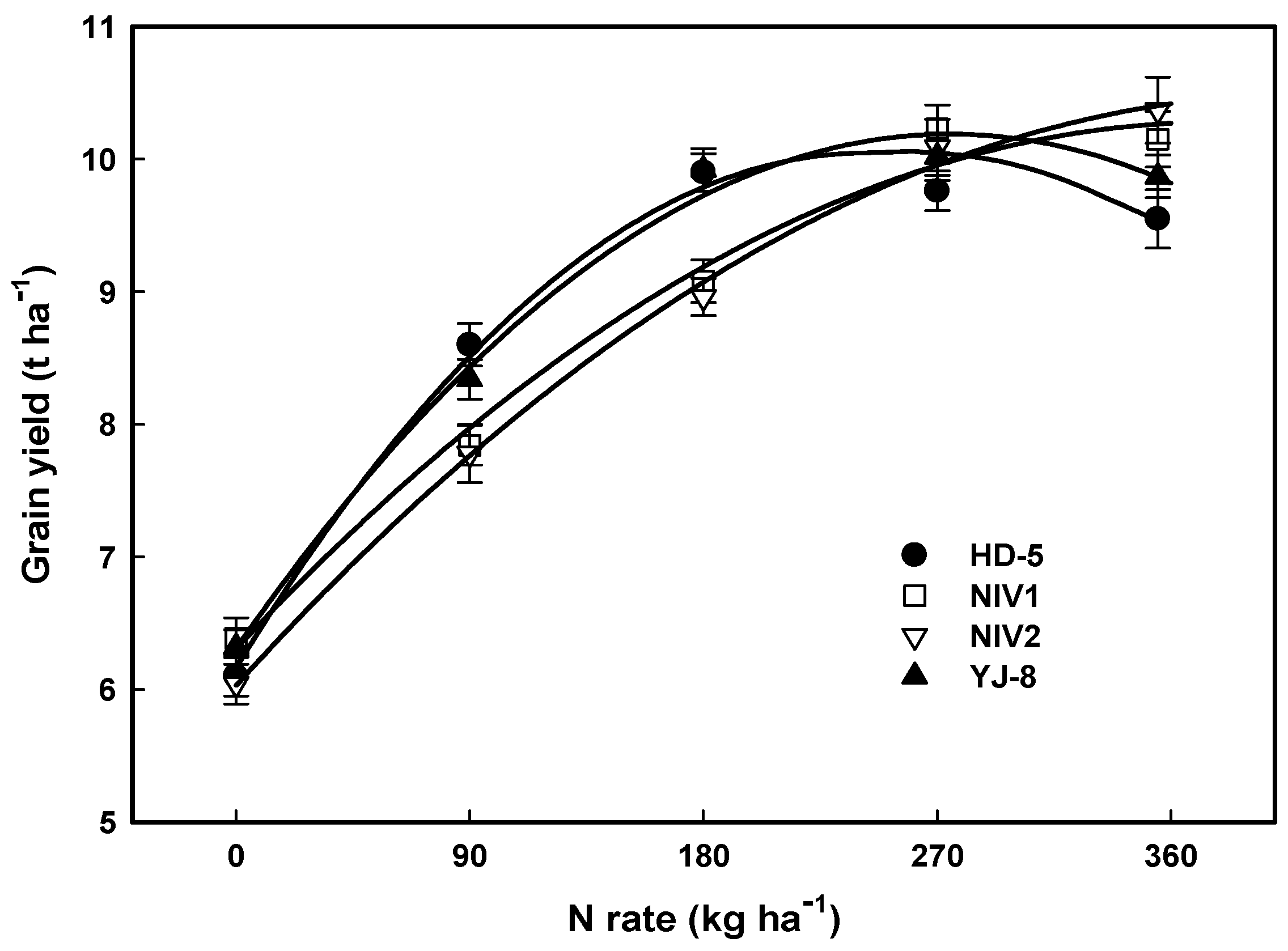
3.1. Grain Yield and NUE
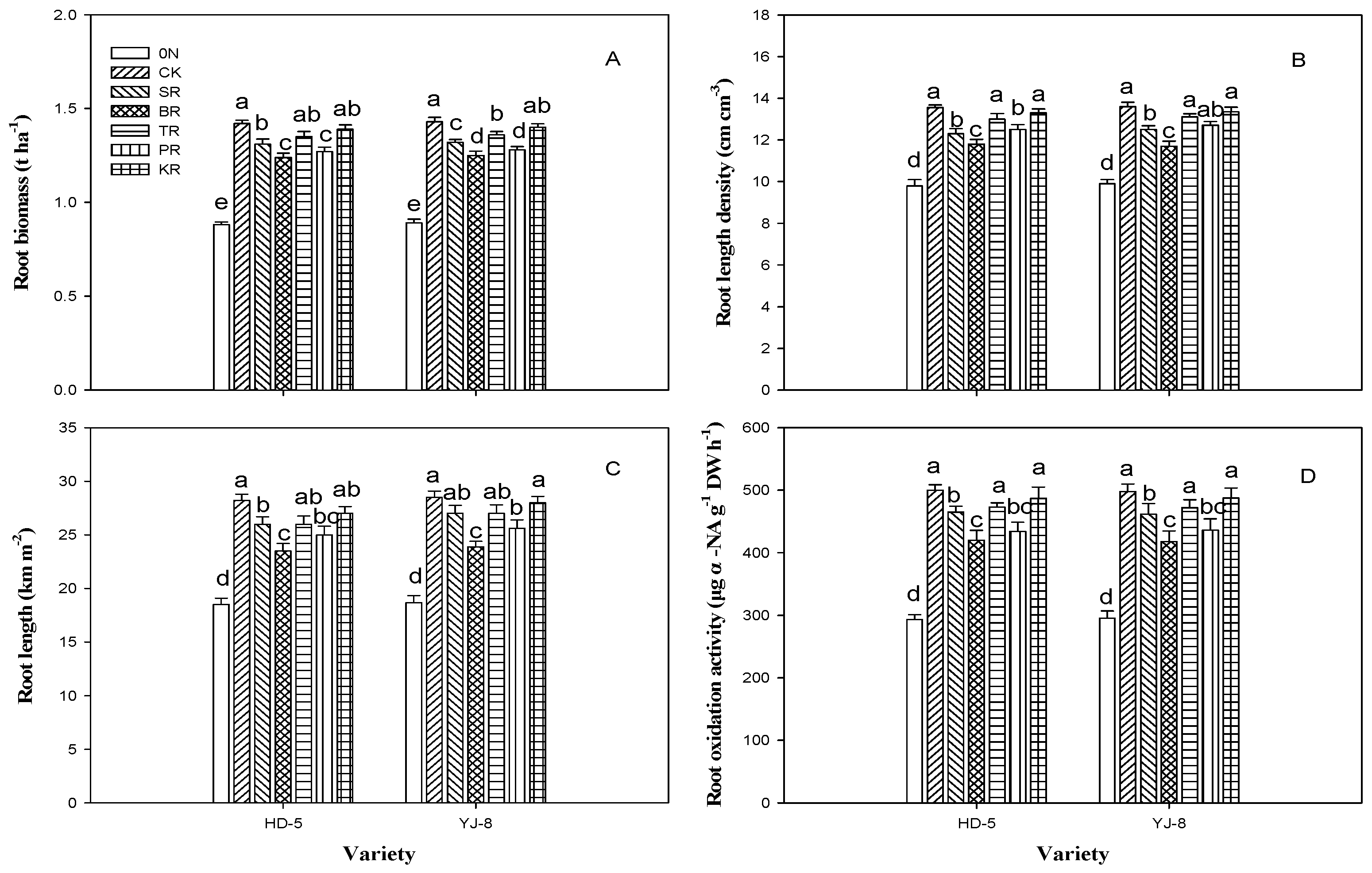
3.2. Root Biomass, Root Length, Root Length Density and Root Oxidation Activity
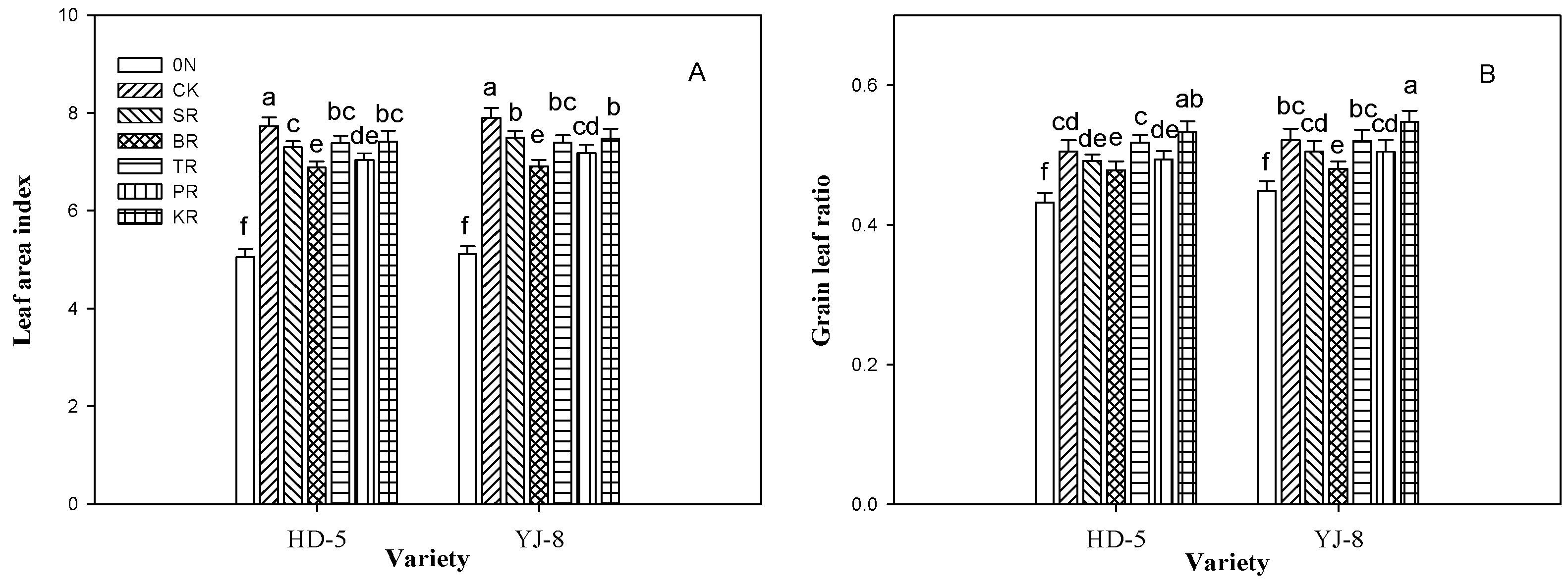
3.3. Leaf Area Index and Grain Leaf Ratio
3.4. Shoot Biomass and Crop Growth Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: Lessons from Arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef]
- Peng, S.; Buresh, R.J.; Huang, J.; Zhong, X.; Zou, Y.; Yang, J.; Wang, G.; Liu, Y.; Hu, R.; Tang, Q.; et al. Improving nitrogen fertilization in rice by site-specific N management: A review. Agron. Sustain. Dev. 2010, 30, 649–656. [Google Scholar] [CrossRef]
- Huang, L.; Yang, D.; Li, X.; Peng, S.; Wang, F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice. Field Crops Res. 2019, 233, 49–58. [Google Scholar] [CrossRef]
- Yu, C.; Huang, X.; Chen, H.; Godfray, H.C.; Wright, J.S.; Hall, J.; Gong, P.; Ni, S.; Qiao, S.; Huang, G.; et al. Managing nitrogen to restore water quality in China. Nature 2019, 567, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application. Field Crops Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.M.; Rothstein, S.J. Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1490–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Q. The Quality of Crop Population; Shanghai Scientific & Technical Publishers: Shanghai, China, 2005; pp. 154–196. [Google Scholar]
- Li, Y.; Chen, Y.; Wu, C.; Tang, X.; Ji, X. Determination of optimum nitrogen application rates in Zhejiang Province, China, based on rice yields and ecological security. J. Integr. Agric. 2015, 14, 2426–2433. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, G.; Liu, L.; Wang, Z.; Wang, X.; Zhang, H.; Yang, J.; Zhang, J. Mid-season nitrogen application strategies for rice varieties differing in panicle size. Field Crops Res. 2013, 150, 9–18. [Google Scholar] [CrossRef]
- Fan, L.; Xu, S.; Hou, P.; Xue, L.; Li, G.; Ding, Y.; Yang, L. Effect of different ratios of basal to tiller nitrogen on rice yield and nitrogen utilization under different soil fertility. Sci. Agric. Sin. 2016, 49, 1872–1884. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain-filling problem in ‘super’ rice. J. Exp. Bot. 2010, 61, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Emran, S.; Krupnik, T.J.; Kumar, V.; Ali, M.Y.; Pittelkow, C.M. Agronomic, economic, and environmental performance of nitrogen rates and source in Bangladesh’s coastal rice agroecosystems. Field Crops Res. 2019, 241, 107567. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.W.; Gutierrez-Suson, J.; Kim, P.J. Optimum N rate for grain yield coincides with minimum greenhouse gas intensity in flooded rice fields. Field Crops Res. 2019, 237, 23–31. [Google Scholar] [CrossRef]
- Qiao, J.; Yang, L.; Yan, T.; Xue, F.; Zhao, D. Rice dry matter and nitrogen accumulation, soil mineral N around root and N leaching, with increasing application rates of fertilizer. Eur. J. Agron. 2013, 49, 93–103. [Google Scholar] [CrossRef]
- Chu, G.; Chen, T.; Wang, Z.; Yang, J.; Zhang, J. Morphological and physiological traits of roots and their relationships with water productivity in water-saving and drought-resistant rice. Field Crops Res. 2014, 162, 108–119. [Google Scholar] [CrossRef]
- Fu, P.; Wang, J.; Zhang, T.; Huang, J.; Peng, S. High nitrogen input causes poor grain filling of spikelets at the panicle base of super hybrid rice. Field Crops Res. 2019, 244, 107635. [Google Scholar] [CrossRef]
- Chen, L.; Peng, X.; Liu, Y.; Li, Z.; Zhang, M.; Li, J. Effect of nutrient management od dry matter accumulation and translocation of rice in cold area. J. Northeast Agric. Univ. 2010, 41, 52–55. [Google Scholar] [CrossRef]
- Ju, C. Agronomic and Physiological Traits for Rice Cultivars Differing in Response to Nitrogen. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2017. [Google Scholar]
- Wang, D.; Chang, Y.; Xu, C.; Wang, Z.; Chen, S.; Chu, G.; Zhang, X. Soil nitrogen distribution and plant nitrogen utilization in direct-seeded rice in response to deep placement of basal fertilizer-nitrogen. Rice Sci. 2019, 26, 404–415. [Google Scholar] [CrossRef]
- Yan, J.; Shen, Q.; Yin, B.; Wan, X. Fertilizer-N uptake and distribution in rice plants using 15N tracer technique. J. Nucl. Agric. Sci. 2009, 23, 487–491. [Google Scholar] [CrossRef]
- Yuan, Q. The Effect of Nitrogen Fertilizer Treatments on the Grain Yield and Root Traits of Indica Rice Varieties with Different Nitrogen Uptake Efficiency. Master’s Thesis, Yangzhou University, Yangzhou, China, 2013. [Google Scholar]
- Zhou, W.; Tian, Y.; Yin, B. Ammonia volatilization and nitrogen balance after topdressing fertilization in paddy fields on Taihu Lake region. Chin. J. Eco-Agric. 2011, 19, 32–36. [Google Scholar] [CrossRef]
- Peng, S. Reflection on China’s rice production strategies during the transition period. Sci. Sin. Vitae 2014, 44, 845–850. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, H.; Zhang, J. Root morphology and physiology in relation to the yield formation of rice. J. Integr. Agric. 2012, 11, 920–926. [Google Scholar] [CrossRef]
- Wu, W.; Cheng, S. Root genetic research, an opportunity and challenge to rice improvement. Field Crops Res. 2014, 165, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Chen, C.; Yuan, Q.; Yang, B.; Zhu, Z.; Cao, W.; Zhong, J.; Zhou, J.; Luo, G.; Wang, Y.; et al. The effect of nitrogen fertilizer treatments on root traits and nitrogen use efficiency in indica rice varieties with high nitrogen absorption efficiency. Acta Ecol. Sin. 2016, 36, 642–651. [Google Scholar] [CrossRef]
- Takai, T.; Matauura, S.; Nishio, T.; Ohsumi, A.; Shiraiwa, T.; Horie, T. Rice yield potential is closely related to crop growth rate during late reproductive period. Field Crops Res. 2006, 96, 328–335. [Google Scholar] [CrossRef]
- Fu, J.; Huang, Z.; Wang, Z.; Yang, J.; Zhang, J. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice. Field Crops Res. 2011, 123, 170–182. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment 1 | Proportion | N Rate (kg ha−1) | ||||
|---|---|---|---|---|---|---|
| PT 2 | ET | PI | SD | Total | ||
| N0 | - | 0 | 0 | 0 | 0 | 0 |
| CK | 4:2:2:2 | 100 | 50 | 50 | 50 | 250 |
| SR | 4:2:2:2 | 80 | 40 | 40 | 40 | 200 |
| BR | 2:2:2:2 | 50 | 50 | 50 | 50 | 200 |
| TR | 4:0:2:2 | 100 | 0 | 50 | 50 | 200 |
| PR | 4:2:0:2 | 100 | 50 | 0 | 50 | 200 |
| KR | 4:2:2:0 | 100 | 50 | 50 | 0 | 200 |
| Source of Variation | df | Grain Yield | Panicles per m2 | Spikelets per Panicle | Filled Grains Rate | N Uptake | Shoot Biomass | Agronomic N Use Efficiency |
|---|---|---|---|---|---|---|---|---|
| Year(Y) | 1 | NS | NS | NS | NS | NS | NS | NS |
| Variety(V) | 1 | NS | NS | * | * | NS | NS | NS |
| N treatments (N) | 6 | ** | ** | ** | * | ** | ** | ** |
| Y × V | 1 | NS | NS | NS | NS | NS | NS | NS |
| Y × N | 6 | NS | NS | NS | NS | NS | NS | NS |
| V × N | 6 | ** | ** | ** | * | ** | ** | ** |
| Variety 1 | Treatment 2 | Grain Yield (t ha−1) | Panicles per m2 | Spikelets per Panicle | Total Spikelets (×103 m−2) | Filled Grain
(%) | Grain Weight (mg) |
|---|---|---|---|---|---|---|---|
| HD-5 | N0 | 5.65e 3 | 202 d | 108 e | 21.2 f | 93.4 a | 27.6 a |
| CK | 9.90 a | 277 ab | 141 ab | 39.1 ab | 92.4 a | 27.5 a | |
| SR | 9.07 c | 267 b | 135 bc | 36.0 cd | 91.8 ab | 27.4 a | |
| BR | 8.31 d | 253 c | 130 cd | 32.9 e | 92.2 ab | 27.4 a | |
| TR | 9.48 b | 270 ab | 140 ab | 37.8 bc | 91.9 ab | 27.3 a | |
| PR | 8.84 c | 276 ab | 126 d | 34.8 d | 92.1 ab | 27.6 a | |
| KR | 9.67 ab | 278 ab | 141 ab | 39.5 ab | 90.5 c | 27.1 ab | |
| YJ-8 | N0 | 5.74 e | 201 d | 114 e | 22.3 f | 92.6 a | 27.0 ab |
| CK | 9.97 a | 284 a | 145 a | 41.2 a | 91.1 bc | 26.6 ab | |
| SR | 9.12 c | 274 ab | 138 ab | 37.8 bc | 90.3 c | 26.7 ab | |
| BR | 8.28 d | 252 c | 134 bc | 33.8 de | 91.5 ab | 26.8 ab | |
| TR | 9.33 bc | 271 ab | 142 ab | 38.5 b | 90.5 c | 26.8 ab | |
| PR | 8.90 c | 283 a | 128 cd | 36.2 cd | 91.0 bc | 27.0 ab | |
| KR | 9.65 ab | 284 a | 144 a | 40.9 a | 89.7 d | 26.3 b |
| Variety 1 | Treatment 2 | N Uptake (kg ha−1) | IE (kg kg−1) | RE (%) | AE (kg kg−1) | PE (kg kg−1) | PFP (kg kg−1) |
|---|---|---|---|---|---|---|---|
| HD-5 | N0 | 72.4f 3 | 78.1 a | ||||
| CK | 163.6 a | 60.5 b | 36.5 c | 17.0 cd | 46.5 a | 39.6 c | |
| SR | 145.7 cd | 62.2 b | 36.7 c | 17.1 cd | 46.6 a | 45.3 a | |
| BR | 133.5 e | 62.2 b | 30.6 e | 13.3 e | 43.5 c | 41.5 b | |
| TR | 155.4 ab | 61.0 b | 41.5 a | 19.1 ab | 46.1 ab | 47.4 a | |
| PR | 138.8 de | 62.4 b | 34.6 d | 15.9 d | 46.0 ab | 44.2 ab | |
| KR | 156.8 ab | 61.7 b | 42.2 a | 20.1 a | 47.7 a | 48.4 a | |
| YJ-8 | N0 | 74.5 f | 77.0 a | ||||
| CK | 165.2 a | 60.3 b | 36.3 c | 16.9 cd | 46.6 a | 39.9 c | |
| SR | 149.7 bc | 60.9 b | 37.6 bc | 16.9 cd | 44.9 b | 45.6 a | |
| BR | 134.5 e | 61.6 b | 30.0 e | 12.7 e | 42.4 c | 41.4 b | |
| TR | 152.3 bc | 61.3 b | 38.9 b | 18.0 bc | 46.2 ab | 46.7 a | |
| PR | 140.4 de | 62.1 b | 34.5 d | 15.8 d | 45.9 ab | 44.5 ab | |
| KR | 161.1 a | 59.9 b | 43.3 a | 19.5 a | 45.1 b | 48.2 a |
| Variety 1 | Treatment 2 | Crop Growth Rate (g cm−2 d−1) | Shoot Biomass (t ha−1) | ||
|---|---|---|---|---|---|
| Jointing-Heading | Heading-Maturity | Heading | Maturity | ||
| HD-5 | N0 | 9.24d 3 | 9.38 d | 6.39 d | 11.36 g |
| CK | 18.83 a | 15.38 a | 11.75 a | 19.90 a | |
| SR | 17.94 b | 14.64 ab | 10.71 b | 18.47 cd | |
| BR | 16.32 c | 13.64 c | 9.73 c | 16.96 f | |
| TR | 18.09 ab | 15.28 a | 10.74 b | 18.84 bc | |
| PR | 16.76 c | 14.14 bc | 10.18 bc | 17.67 e | |
| KR | 18.58 ab | 15.02 ab | 11.03 ab | 18.99 bc | |
| YJ-8 | N0 | 9.15 d | 9.38 d | 6.32 d | 11.29 g |
| CK | 19.14 a | 15.19 a | 11.84 a | 19.89 a | |
| SR | 18.38 ab | 14.61 ab | 10.62 b | 18.38 cd | |
| BR | 16.48 c | 13.64 c | 9.65 c | 16.88 f | |
| TR | 18.51 ab | 14.77 ab | 10.83 b | 18.66 bc | |
| PR | 16.45 c | 14.70 ab | 10.22 bc | 18.01 de | |
| KR | 18.71 a | 15.31 a | 11.12 ab | 19.23 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, C.; Zhu, Y.; Liu, T.; Sun, C. The Effect of Nitrogen Reduction at Different Stages on Grain Yield and Nitrogen Use Efficiency for Nitrogen Efficient Rice Varieties. Agronomy 2021, 11, 462. https://doi.org/10.3390/agronomy11030462
Ju C, Zhu Y, Liu T, Sun C. The Effect of Nitrogen Reduction at Different Stages on Grain Yield and Nitrogen Use Efficiency for Nitrogen Efficient Rice Varieties. Agronomy. 2021; 11(3):462. https://doi.org/10.3390/agronomy11030462
Chicago/Turabian StyleJu, Chengxin, Yiwen Zhu, Tao Liu, and Chengming Sun. 2021. "The Effect of Nitrogen Reduction at Different Stages on Grain Yield and Nitrogen Use Efficiency for Nitrogen Efficient Rice Varieties" Agronomy 11, no. 3: 462. https://doi.org/10.3390/agronomy11030462
APA StyleJu, C., Zhu, Y., Liu, T., & Sun, C. (2021). The Effect of Nitrogen Reduction at Different Stages on Grain Yield and Nitrogen Use Efficiency for Nitrogen Efficient Rice Varieties. Agronomy, 11(3), 462. https://doi.org/10.3390/agronomy11030462