
Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple


Abstract
:1. Introduction
2. Materials and Methods
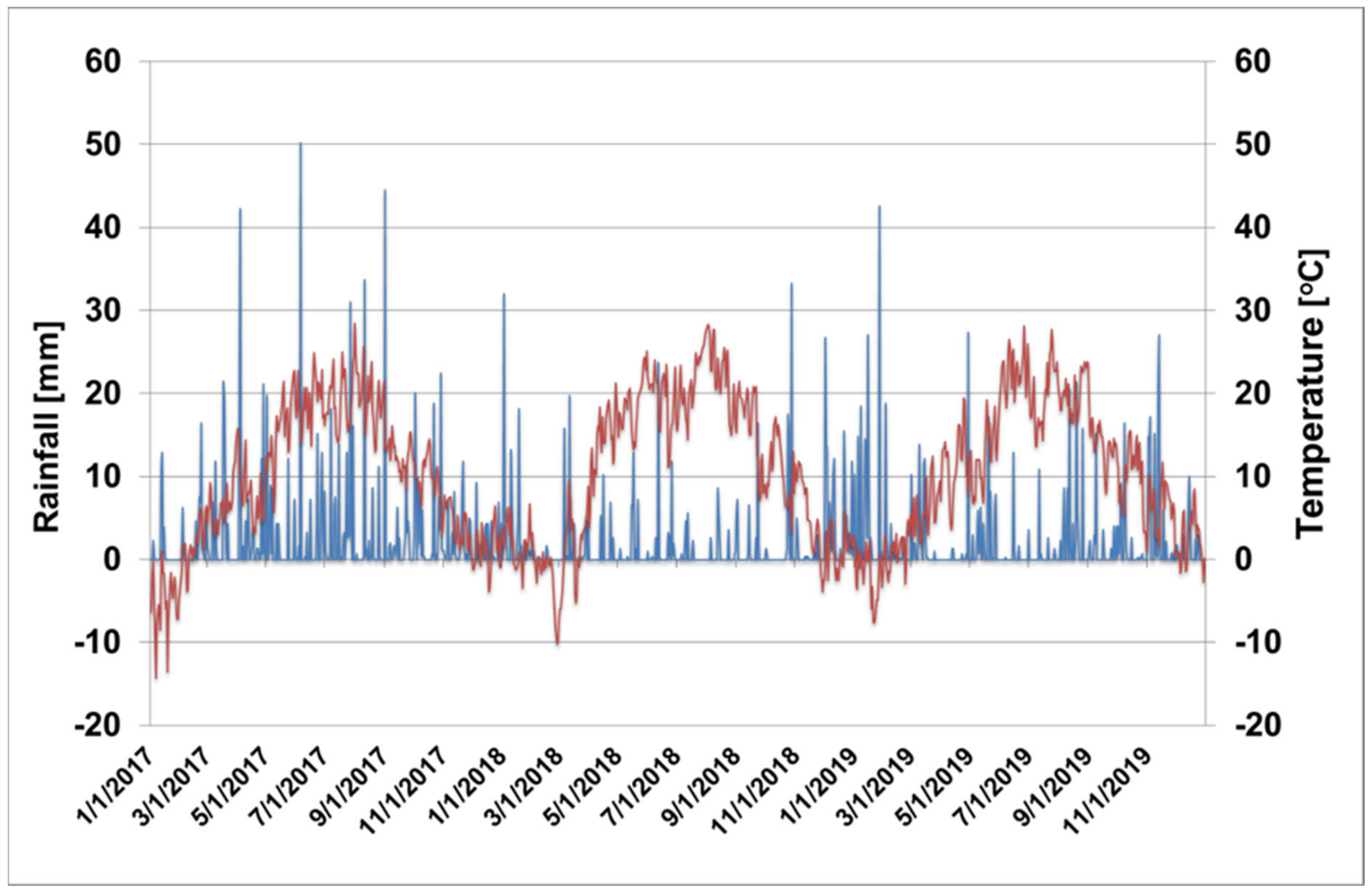
2.1. Plant Material, Orchard Management, and Environment
2.2. Methods and Treatments
2.3. Data Processing and Analyses
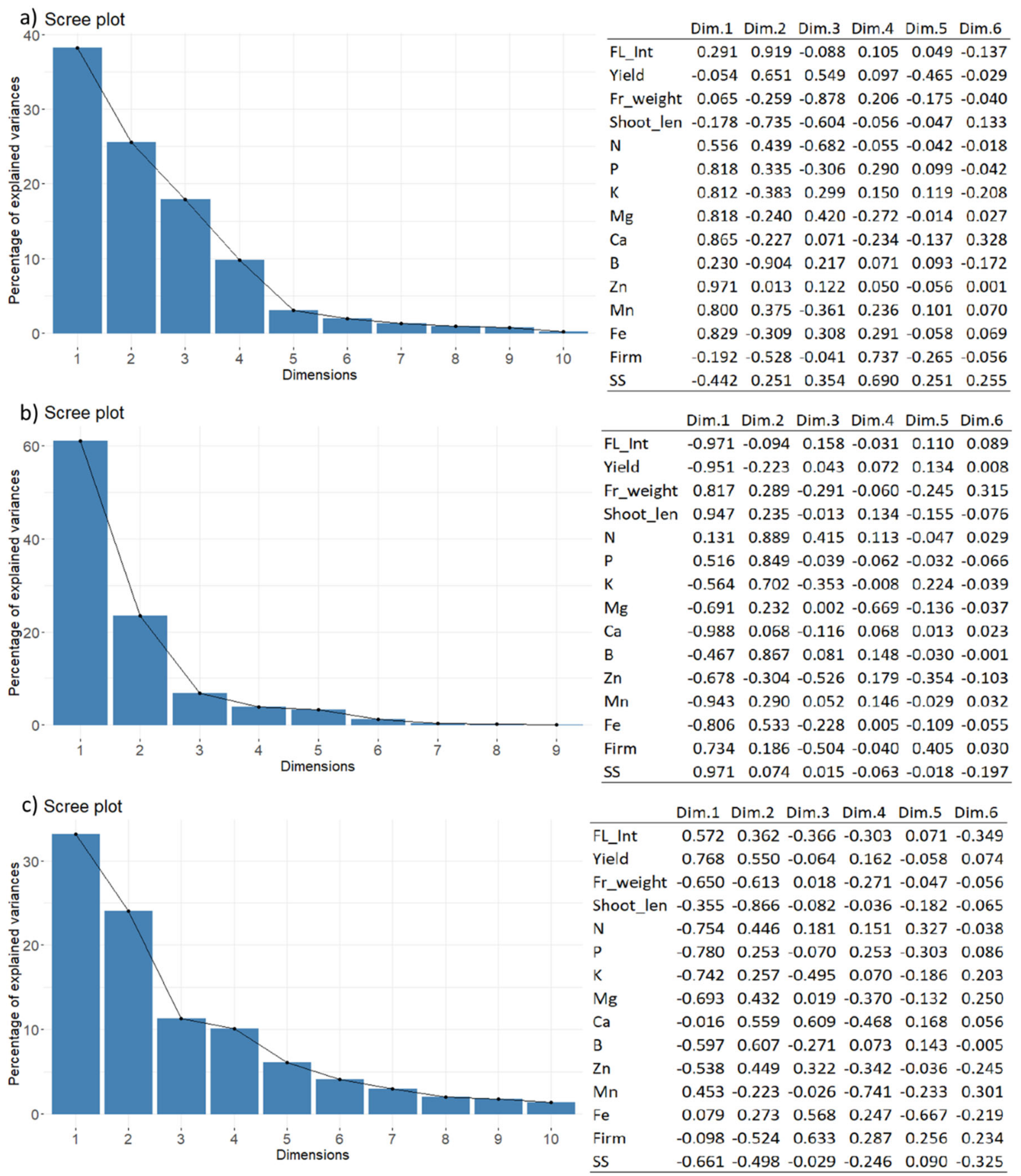
3. Results
3.1. Shoot Growth, Fruit Production, and Quality
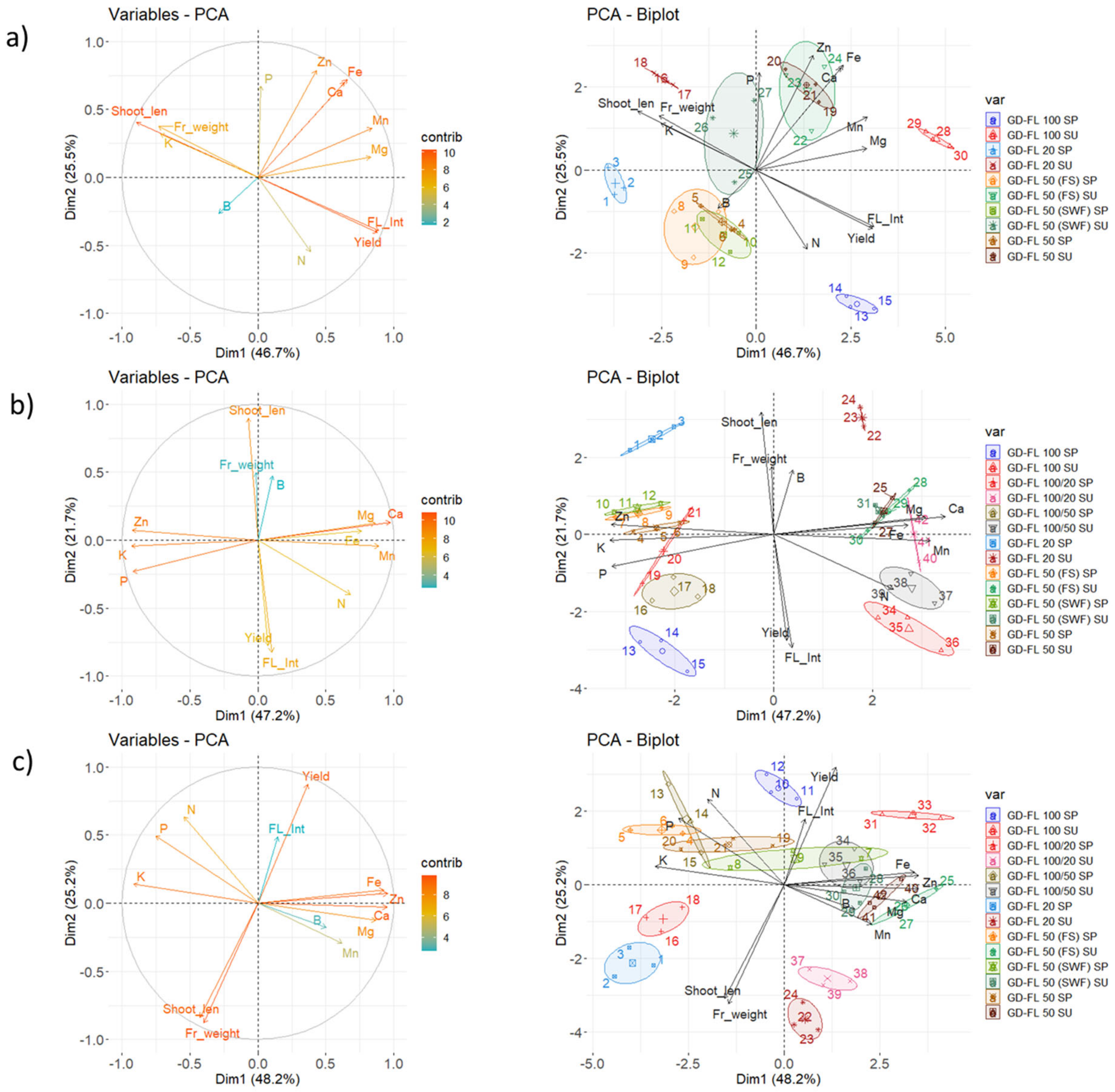
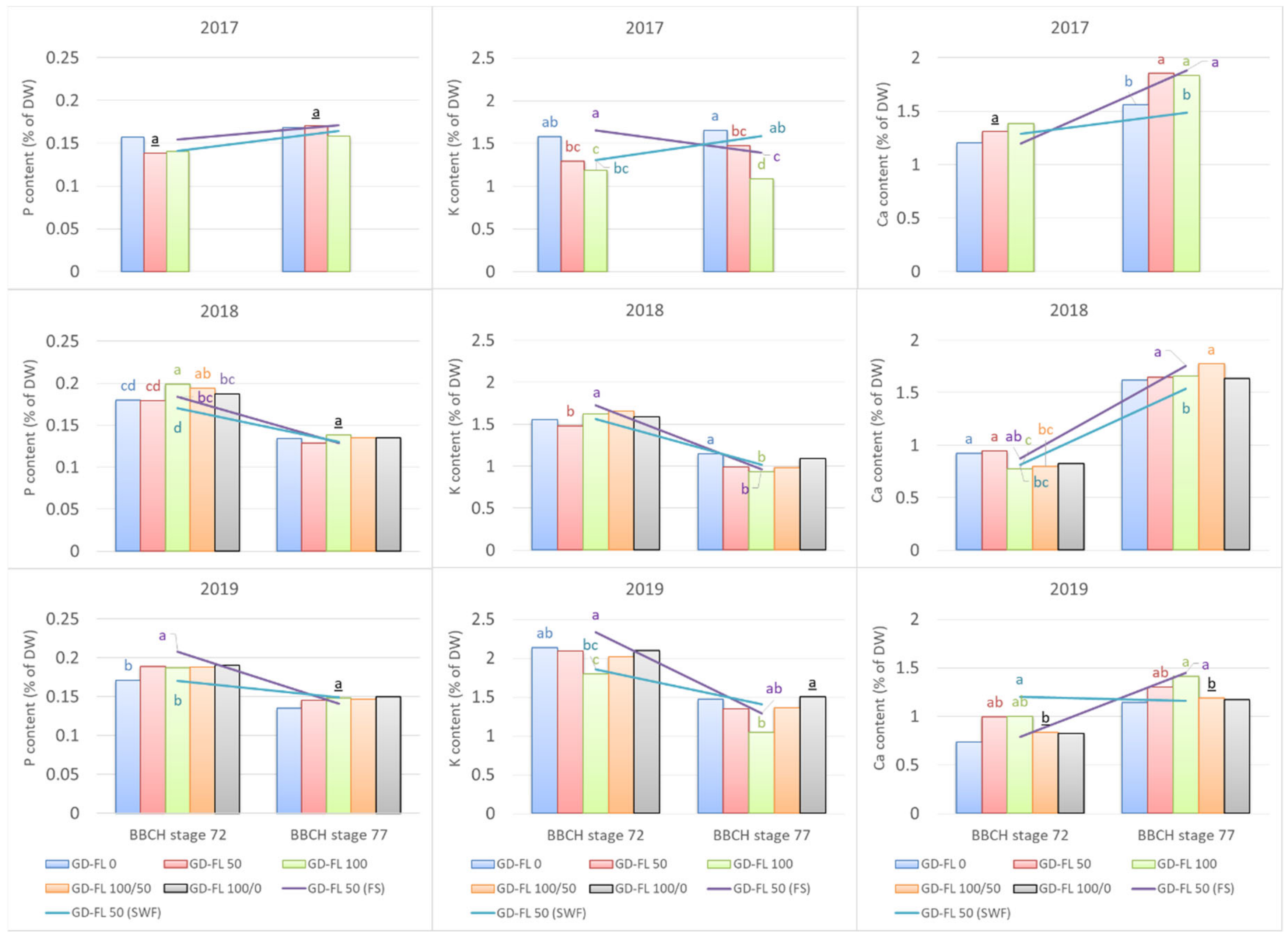
3.2. Mineral Content in Leaf Dry Matter
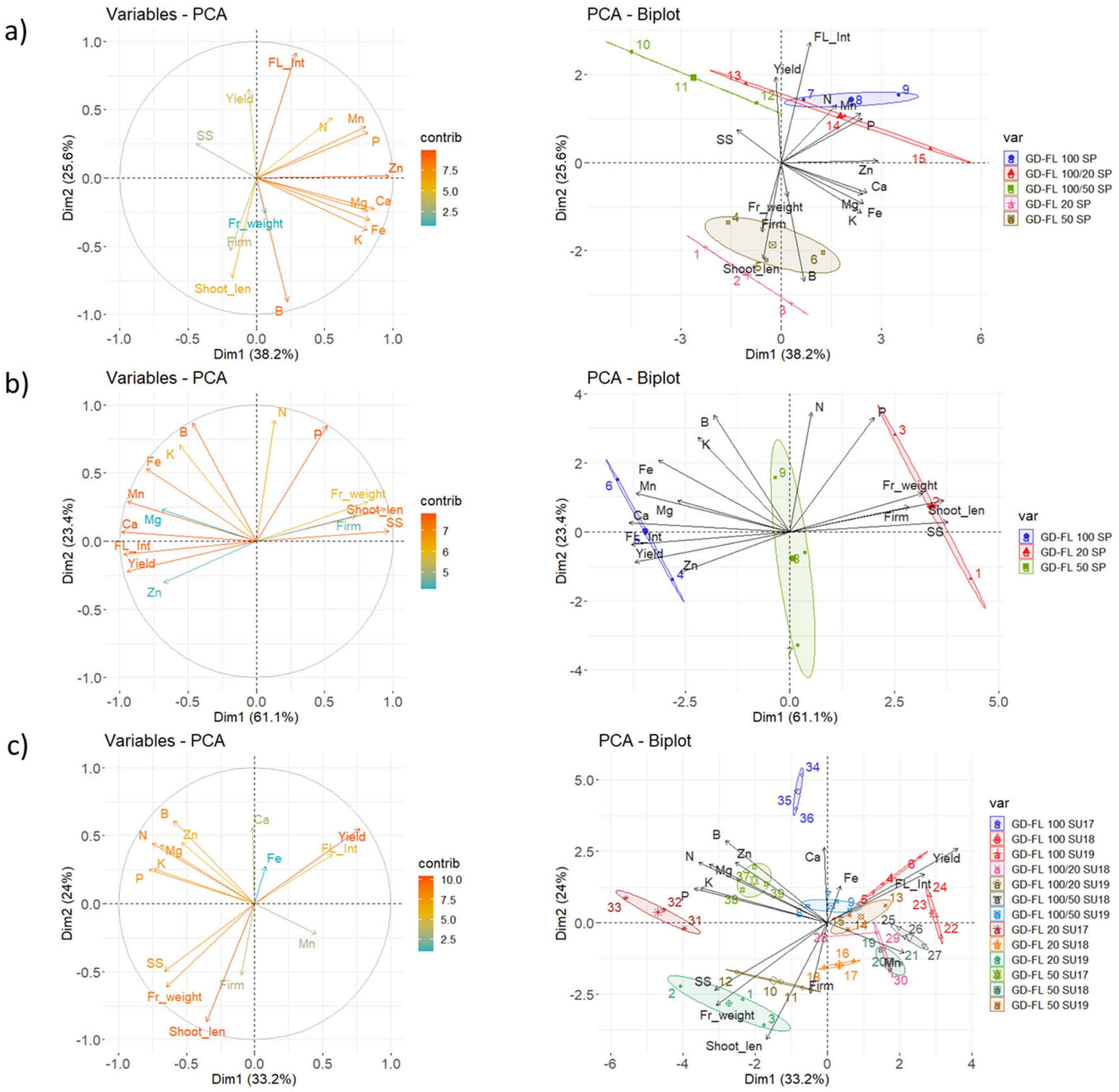
3.3. Mineral Content in Fruit Dry Matter of Apple Trees
4. Discussion
4.1. Nutritional Status of Apple Tree Leaves According to the Fruit Load and Shoot Growth Intensity
4.2. Nutritional Status of Apple Tree Fruit According to the Fruit Load and Shoot Growth Intensity
4.3. Relation of the Fruit Quality to Crop Load and Nutritional Status of Apple Tree
4.4. Interpretation of the Relationship of the Crop Load and Nutritional Status of Fruit Trees
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tromp, J. Mineral absorption and distribution in young apple trees under various environmental conditions. In Mineral Nutrition of Fruit Trees; Atkinson, D., Jackson, J.E., Sharples, R.O., Waller, W.M., Eds.; Butterworths: London, UK, 1980; pp. 173–182. [Google Scholar]
- Tromp, J. Mineral nutrition. In Fundamentals of Temperate Zone Tree Fruit Production; Tromp, J., Webster, A.D., Wertheim, S.J., Eds.; Backhuys Publishers: Leiden, Poland, 2005; pp. 55–64. [Google Scholar]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root development and nutrient uptake. Crit. Rev. Plant Sci. 2006, 25, 279–301. [Google Scholar] [CrossRef]
- Hansen, P. The effect of cropping on the distribution of growth in apple trees. Tidsskr Planteavl 1971, 75, 119–127. [Google Scholar]
- Oliveira, C.M.; Priestley, C.A. Carbohydrate reserves in deciduous fruit trees. Hortic. Rev. 1988, 10, 403–430. [Google Scholar] [CrossRef]
- Grossman, Y.L.; DeJong, T.M. Maximum vegetative growth potential and seasonal patterns of resource dynamics during peach growth. Ann. Bot. 1995, 76, 473–482. Available online: https://www.jstor.org/stable/42765149 (accessed on 20 November 2020). [CrossRef]
- Lakso, A.N.; Goffinet, M.C. Apple fruit growth. N. Y. Fruit Quart 2013, 21, 11–14. [Google Scholar]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How can we make plants grow faster? A source–sink perspective on growth rate. J. Exp. Bot. 2015, 67, 31–45. [Google Scholar] [CrossRef]
- Millard, P. Ecophysiology of the internal cycling of nitrogen for tree growth. J. Plant. Nutr. Soil Sci. 1996, 159, 1–10. [Google Scholar] [CrossRef]
- Neilsen, G.H.; Neilsen, D. Nutritional requirements of apple. In Apples: Botany, Production and Uses; Ferree, D.C., Warrington, I.J., Eds.; CAB International: Wernstein, Germany, 2003; pp. 267–302. [Google Scholar]
- Aichner, M.; Stimpfl, E. Seasonal pattern and interpretation of mineral nutrient concentrations in apple leaves. Acta Hortic. 2002, 594, 377–382. [Google Scholar] [CrossRef]
- Nachtigall, G.R.; Dechen, A.R. Seasonality of nutrients in leaves and fruits of apple trees. Sci. Agric. 2006, 63, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Čonka, P.; Bělíková, H.; Mészáros, M.; Kurešová, G.; Náměstek, J. Evaluation of seasonal variation in mineral composition of leaves and fruits of Malus domestica Borkh. ’Golden Delicious´ throughout growing season. Vědecké Práce Ovocnářské 2017, 25, 89–102. [Google Scholar]
- Fernández, V.; Sotiropoulos, T.; Brown, P.H. Foliar Fertilization: Scientific Principles and Field Practices; International Fertilizer Industry Association (IFA): Paris, France, 2013; Available online: https://1lib.eu/book/2977644/93baa2?regionChanged=&redirect=28394966 (accessed on 20 November 2020).
- Briat, J.-F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Socha, A.L.; Eguerinot, M.L. Mneuvering manganese: The role of transporter gene family members in manganese uptake and mobilization in plants. Front. Plant Sci. 2014, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Kalcsits, L.; Van Der Heijden, G.; Reid, M.; Mullin, K. Calcium absorption during fruit development in ‘honeycrisp’ apple measured using 44ca as a stable isotope tracer. HortScience 2017, 52, 1804–1809. [Google Scholar] [CrossRef] [Green Version]
- Blanke, M.M.; Lenz, F. Fruit photosynthesis. Plant. Cell Environ. 1989, 12, 31–46. [Google Scholar] [CrossRef]
- Palmer, J.W. Effects of varying crop load on photosynthesis, dry matter production and partitioning of Crispin/M.27 apple trees. Tree Physiol. 1992, 11, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P. The effect of cropping on uptake, contents and distribution of nutrients in apple trees. Tidsskr. Planteavl 1971, 75, 615–625. [Google Scholar]
- Xia, G.; Cheng, L.; Lakso, A.; Goffinet, M. Effects of nitrogen supply on source-sink balance and fruit size of ‘gala’ apple trees. J. Am. Soc. Hortic. Sci. 2009, 134, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Weinbaum, S.A.; Niederholzer, F.J.A.; Ponchner, S.; Rosecrance, R.C.; Carlson, R.M.; Whittlesey, A.C.; Muraoka, T.T. Nutrient uptake by cropping and defruited field-grown ‘Frendi’ prune trees. J. Am. Soc. Hortic. Sci. 1994, 119, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Samuolienė, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between apple tree rootstock, crop-load, plant nutritional status and yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Krasniqi, A.-L.; Blanke, M.; Kunz, A.; Damerow, L.; Lakso, A.; Meland, M. Alternate bearing in fruit tree crops: Past, present and future. Acta Hortic. 2017, 1177, 241–248. [Google Scholar] [CrossRef]
- Tromp, J. lower-bud formation in pome fruits as affected by fruit thinning. Plant. Growth Regul. 2000, 31, 27–34. [Google Scholar] [CrossRef]
- Link, H. Significance of flower and fruit thinning on fruit quality. Plant. Growth Regul. 2000, 31, 17–26. [Google Scholar] [CrossRef]
- Schröder, M.; Link, H. Calcium content in apple fruits after thinning treatments in relation to crop load, fruit size and leaf area. Acta Hortic. 2002, 594, 541–545. [Google Scholar] [CrossRef]
- Fallahi, E.; Fallahi, B.; Retamales, J.; Valdes, C.; Tabatabaei, S. Prediction of apple fruit quality using preharvest mineral nutrients. Acta Hortic. 2006, 721, 259–264. [Google Scholar] [CrossRef]
- Neilsen, D.; Neilsen, G.; Herbert, L.; Guak, S. Effect of irrigation and crop load management on fruit nutrition and quality for ambrosia/m.9 apple. Acta Hortic. 2010, 868, 63–72. [Google Scholar] [CrossRef]
- Fallahi, E.; Conway, W.S.; Hickey, K.; Sams, C.E. The role of calcium and nitrogen in postharvest quality and disease resistance of apples. HortScience 1997, 32, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, W. Ernährungsstörungen bei Kulturpflanzen; VEB Gustav Fischer Verlag: Jena, Germany, 1988; p. 762. [Google Scholar]
- Da Silva, G.G.C.; Neves, J.C.L.; Alvarez , V.V.H.; Leite, F.P. Nutritional diagnosis for eucalypt by DRIS, M-DRIS, and CND. Sci. Agric. 2004, 61, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Filho, F.D.A.A. DRIS: Concepts and applications on nutritional diagnosis in fruit crops. Sci. Agric. 2004, 61, 550–560. [Google Scholar] [CrossRef]
- Nachtigall, G.R.; Dechen, A.R. DRIS norms for evaluating the nutritional state of apple tree. Sci. Agric. 2007, 64, 282–287. [Google Scholar] [CrossRef] [Green Version]
- De Wit, J. ‘Klik’ pruning. Sad Nowocz. 2008, 34, 10–12. [Google Scholar]
- Meier, U. Growth stages of mono-and dicotyledonous plants. In BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlín, Germany, 2001. [Google Scholar] [CrossRef]
- Kon, T.M.; Schupp, J.R. Thinning tall spindle apple based on estimations made with a hand-thinning gauge. HortTechnology 2013, 23, 830–835. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-1. 2019. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 20 November 2020).
- Kassambara, A. Factoextra: Practical Guide to Principal Component Methods in R. R Package Version 1.0.7. 2017. Available online: https://cran.r-project.org/web/packages/factoextra/index.html (accessed on 20 November 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Flowering Rate and Fruit Load |
|---|---|
| GD-FL 20 | control with trees at a low flowering rate of 2–3 points and a fruit set of up to 20% of optimal fruit set. |
| GD-FL 50 | trees at moderate flowering rate of 4–6 points and a moderate fruit set of 50% of the optimal fruit set. |
| GD-FL 100 | high flowering rate of about 8–9 points and a full fruit set at 100% of the of optimal fruit set (five fruit per cm2 of the branch cross-sectional area). |
| GD-FL 100/50 | high flowering rate of about 8–9 points and reduced fruit set at 50% of the optimal fruit set. |
| GD-FL 100/20 | high flowering rate of 8–9 points and fruit set reduced to 20% of the of optimal fruit set. |
| Year | Group | Flowering (1–9) | Yield (kg/tree) | Fruit Weight (g) | Firmness kg/cm2 | Soluble Solids (° brix) | Extension Shoot Length (mm) |
|---|---|---|---|---|---|---|---|
| 2017 | GD-FL 20 | 1.9 c | 10.58 c | 211.8 a | 7.682 a | 16.21 a | 714 a |
| GD-FL 50 | 4.4 b | 31.44 b | 188.5 a | 7.673 a | 14.24 b | 623 b | |
| GD-FL 100 | 8.9 a | 52.92 a | 144.6 b | 7.177 a | 12.20 c | 538 c | |
| 2018 | GD-FL 20 | 2.4 d | 34.16 b | 181.0 ab | 7.735 a | 12.60 b | 795 a |
| GD-FL 50 | 4.7 c | 40.20 ab | 162.7 bc | 7.971 a | 13.48 ab | 708 bc | |
| GD-FL 100 | 8.6 ab | 49.36 a | 138.8 c | 7.318 b | 12.76 b | 618 d | |
| GD-FL 100/50 | 7.8 b | 48.68 a | 160.0 bc | 7.724 a | 14.29 a | 659 cd | |
| GD-FL 100/20 | 9.0 a | 38.26 ab | 197.2 a | 7.645 ab | 12.66 b | 728 ab | |
| 2019 | GD-FL 20 | 2.7 c | 10.52 b | 251.0 a | 7.779 a | 16.70 a | 876 a |
| GD-FL 50 | 4.9 b | 39.87 a | 177.6 b | 7.326 b | 13.81 b | 671 bc | |
| GD-FL 100 | 8.7 a | 48.47 a | 134.9 c | 7.184 bc | 11.96 c | 589 c | |
| GD-FL 100/50 | 7.9 a | 44.10 a | 172.2 b | 6.897 c | 14.45 b | 703 b | |
| GD-FL 100/20 | 8.3 a | 11.12 b | 242.8 a | 7.426 ab | 16.85 a | 835 a | |
| Year effect 1 | ns. | *** | *** | ** | *** | * | |
| Group effect 1 | *** | *** | *** | *** | *** | *** | |
| Group × Year 1 | ns. | *** | *** | *** | *** | ** | |
| Year effect 2 | * | ns. | ns. | ns. | ns. | ns. | |
| Group effect 2 | *** | *** | *** | *** | *** | *** | |
| Group × Year 2 | * | ns. | * | ns. | ns. | ns. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mészáros, M.; Hnátková, H.; Čonka, P.; Náměstek, J. Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple. Agronomy 2021, 11, 506. https://doi.org/10.3390/agronomy11030506
Mészáros M, Hnátková H, Čonka P, Náměstek J. Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple. Agronomy. 2021; 11(3):506. https://doi.org/10.3390/agronomy11030506
Chicago/Turabian StyleMészáros, Martin, Hana Hnátková, Patrik Čonka, and Jan Náměstek. 2021. "Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple" Agronomy 11, no. 3: 506. https://doi.org/10.3390/agronomy11030506
APA StyleMészáros, M., Hnátková, H., Čonka, P., & Náměstek, J. (2021). Linking Mineral Nutrition and Fruit Quality to Growth Intensity and Crop Load in Apple. Agronomy, 11(3), 506. https://doi.org/10.3390/agronomy11030506