
Temporal Variability in the Rhizosphere Bacterial and Fungal Community Structure in the Melon Crop Grown in a Closed Hydroponic System

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Analytical Methods
2.3. DNA Extraction and Molecular Analysis
2.4. Statistical Analysis
3. Results
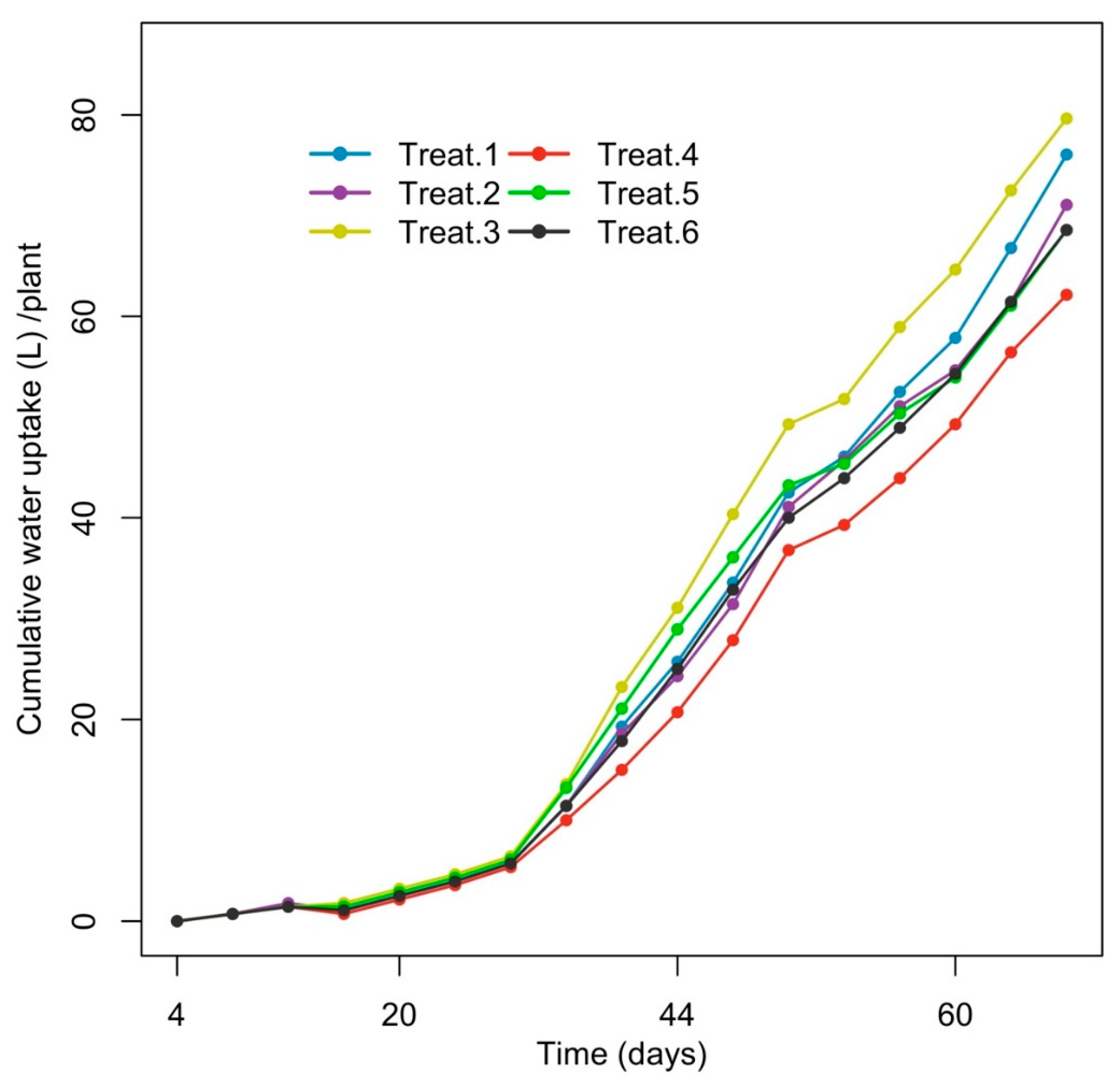
3.1. Cumulative Water Uptake
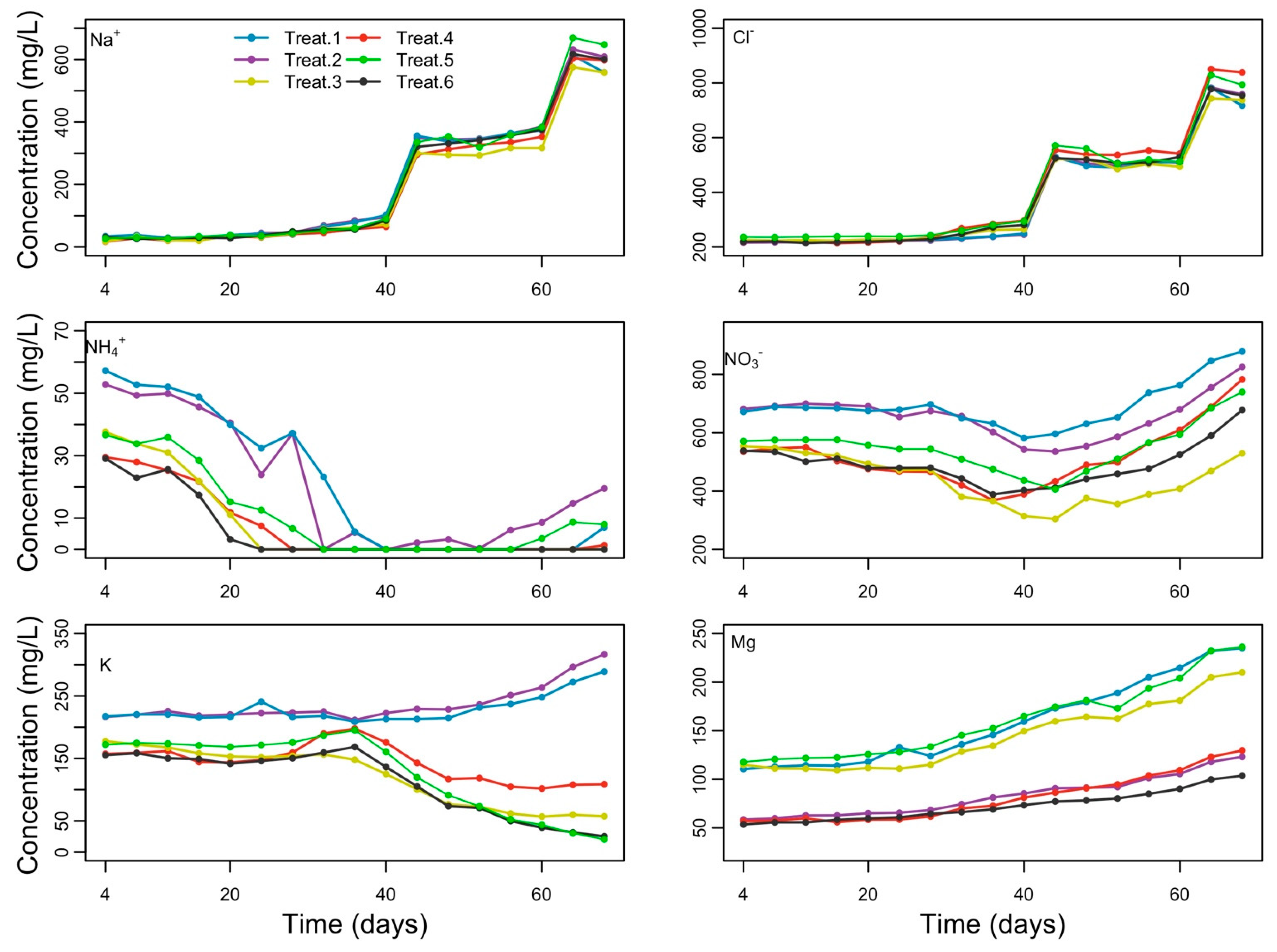
3.2. Nutrient Accumulation in the Root Zone
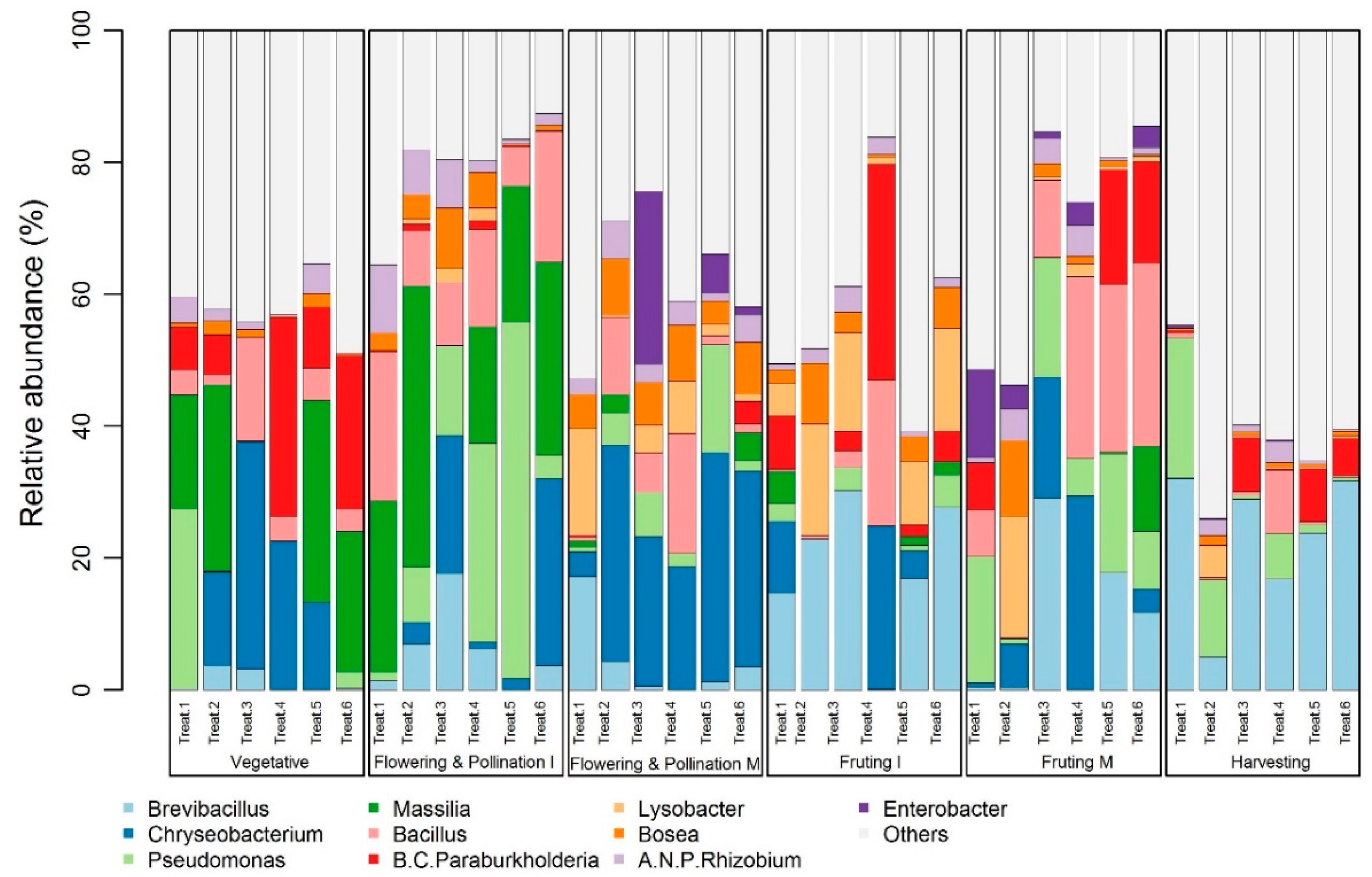
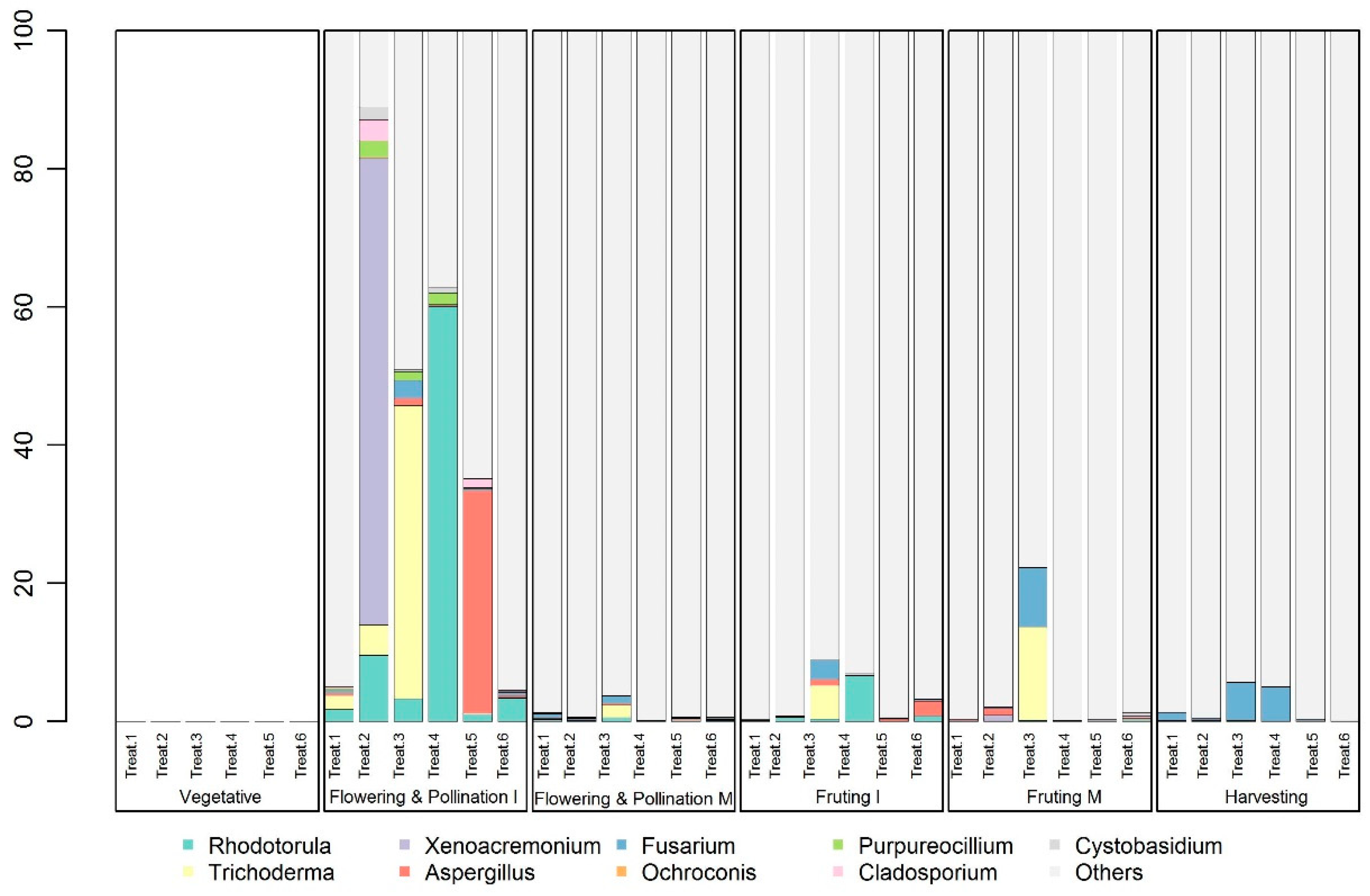
3.3. Shift in Microbial Community Composition in the Melon Plant Rhizosphere
3.4. Shifts in the Relative Abundances of Specific Microbial Taxa
3.5. Melon Yield and Fruit Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rouphael, Y.; Colla, G. Growth, yield, fruit quality and nutrient uptake of hydroponically cultivated zucchini squash as affected by irrigation systems and growing seasons. Sci. Hortic. (Amst.) 2005, 105, 177–195. [Google Scholar] [CrossRef]
- Savvas, D.; Pappa, V.A.; Kotsiras, A.; Gizas, G. NaCl accumulation in a cucumber crop grown in a completely closed hydroponic system as influenced by NaCl concentration in irrigation water. Eur. J. Hortic. Sci. 2005, 70, 217. [Google Scholar]
- Shaaban, M.; Van Zwieten, L.; Bashir, S.; Younas, A.; Nunez-Delgado, A.; Chhajro, M.A.; Kubar, K.A.; Ali, U.; Rana, M.S.; Mehmood, M.A. A concise review of biochar application to agricultural soils to improve soil conditions and fight pollution. J. Environ. Manag. 2018, 228, 429–440. [Google Scholar] [CrossRef]
- Reis, S.; Bekunda, M.; Howard, C.M.; Karanja, N.; Winiwarter, W.; Yan, X.; Bleeker, A.; Sutton, M.A. Synthesis and review: Tackling the nitrogen management challenge: From global to local scales. Environ. Res. Lett. 2016, 11, 120205. [Google Scholar] [CrossRef]
- Savvas, D.; Passam, I. Hydroponic Production of Vegetables and Ornamentals; Savvas, D., Passam, H., Eds.; Embryo Publications: Athens, Greece, 2002; ISBN 9608002125. [Google Scholar]
- Raviv, M.; Krasnovsky, A.; Medina, S.; Reuveni, R. Assessment of various control strategies for recirculation of greenhouse effluents under semi-arid conditions. J. Hortic. Sci. Biotechnol. 1998, 73, 485–491. [Google Scholar] [CrossRef]
- Neocleous, D.; Savvas, D. NaCl accumulation and macronutrient uptake by a melon crop in a closed hydroponic system in relation to water uptake. Agric. Water Manag. 2016, 165, 22–32. [Google Scholar] [CrossRef]
- Neocleous, D.; Savvas, D. Simulating NaCl accumulation in a closed hydroponic crop of zucchini: Impact on macronutrient uptake, growth, yield, and photosynthesis. J. Plant Nutr. Soil Sci. 2017, 180, 283–293. [Google Scholar] [CrossRef]
- Neocleous, D.; Vasilakakis, M. Effects of NaCl stress on red raspberry (Rubus idaeus L. ‘Autumn Bliss’). Sci. Hortic. (Amst.) 2007, 112, 282–289. [Google Scholar] [CrossRef]
- Läuchli, A.; Grattan, S.R. Plant growth and development under salinity stress. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–32. [Google Scholar]
- Stewart-Wade, S.M. Plant pathogens in recycled irrigation water in commercial plant nurseries and greenhouses: Their detection and management. Irrig. Sci. 2011, 29, 267–297. [Google Scholar] [CrossRef]
- Dankwa, A.S.; Machado, R.M.; Perry, J.J. Sources of food contamination in a closed hydroponic system. Lett. Appl. Microbiol. 2020, 70, 55–62. [Google Scholar] [CrossRef]
- Mukhtar, H.; Lin, C.-M.; Wunderlich, R.F.; Cheng, L.-C.; Ko, M.-C.; Lin, Y.-P. Climate and land cover shape the fungal community structure in topsoil. Sci. Total Environ. 2020, 751, 141721. [Google Scholar] [CrossRef]
- Lin, Y.-P.; Wunderlich, R.F.; Lin, C.-M.; Uphoff, N.; Schmeller, D.S.; Shipin, O.V.; Watanabe, T.; Mukhtar, H. Topsoil Microbial Community Structure Responds to Land Cover Type and Environmental Zone in the Western Pacific Region. Sci. Total Environ. 2020, 764, 144349. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Bakker, P.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Kim, M.-S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.-M.; Allen, R.; Melo, I.S. Rhizobacterial volatile emissions regulate auxin homeostasis and cell expansion in Arabidopsis. Planta 2007, 226, 839. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef] [PubMed]
- Rosier, A.; Medeiros, F.H.V.; Bais, H.P. Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant Soil 2018, 428, 35–55. [Google Scholar] [CrossRef] [Green Version]
- Ortíz-Castro, R.; Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; López-Bucio, J. The role of microbial signals in plant growth and development. Plant Signal. Behav. 2009, 4, 701–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, J. Beneficial bacteria and fungi in hydroponic systems: Types and characteristics of hydroponic food production methods. Sci. Hortic. (Amst.) 2015, 195, 206–215. [Google Scholar] [CrossRef]
- Sheridan, C.; Depuydt, P.; De Ro, M.; Petit, C.; Van Gysegem, E.; Delaere, P.; Dixon, M.; Stasiak, M.; Aciksöz, S.B.; Frossard, E. Microbial community dynamics and response to plant growth-promoting microorganisms in the rhizosphere of four common food crops cultivated in hydroponics. Microb. Ecol. 2017, 73, 378–393. [Google Scholar] [CrossRef]
- Mukhtar, H.; Lin, Y.-P.; Anthony, J. Ammonia Oxidizing Archaea and Bacteria in East Asian Paddy Soils—A Mini Review. Environment 2017, 4, 84. [Google Scholar]
- Mukhtar, H.; Lin, Y.-P.; Shipin, O.V.; Petway, J.R. Modeling Nitrogen Dynamics in a Waste Stabilization Pond System Using Flexible Modeling Environment with MCMC. Int. J. Environ. Res. Public Health 2017, 14, 765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhtar, H.; Lin, Y.-P.; Lin, C.-M.; Petway, J.R. Assessing Thermodynamic Parameter Sensitivity for Simulating Temperature Response of Soil Nitrification. Environ. Sci. Process Impacts 2019, 21, 1596–1608. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233. [Google Scholar] [CrossRef]
- Neocleous, D.; Savvas, D. Effect of different macronutrient cation ratios on macronutrient and water uptake by melon (Cucumis melo) grown in recirculating nutrient solution. J. Plant. Nutr. Soil Sci. 2015, 178, 320–332. [Google Scholar] [CrossRef]
- Rath, K.M.; Maheshwari, A.; Rousk, J. Linking microbial community structure to trait distributions and functions using salinity as an environmental filter. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, S.K.; Williams, R.J.; Hofmockel, K.S. Environmental filtering of microbial communities in agricultural soil shifts with crop growth. PLoS ONE 2015, 10, e0134345. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Burger, Y.; Horev, C.; Porat, A.; Edelstein, M. Performance of Galia-type melons grafted on to Cucurbita rootstock in Monosporascus cannonballus-infested and non-infested soils. Ann. Appl. Biol. 2005, 146, 381–387. [Google Scholar] [CrossRef]
- Tzay-Fa, S. Effects of NO3/NH4 Ratio and Depth of Nutrient Solution on the Growth of Greenhouse Muskmelon. J. Agric. Res. China 1989, 38, 1. (In Chinese) [Google Scholar]
- Lin, J.-T. Effect of nutrient temperature on growth and development of deep flow technique hydroponic netted melon. Master’s Thesis, National Taiwan University, Taipei, Taiwan, July 2018. [Google Scholar]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant. 2008, 133, 692–704. [Google Scholar] [CrossRef]
- Senbayram, M.; Gransee, A.; Wahle, V.; Thiel, H. Role of magnesium fertilisers in agriculture: Plant–soil continuum. Crop Pasture Sci. 2016, 66, 1219–1229. [Google Scholar] [CrossRef]
- Hermans, C.; Johnson, G.N.; Strasser, R.J.; Verbruggen, N. Physiological characterisation of magnesium deficiency in sugar beet: Acclimation to low magnesium differentially affects photosystems I and II. Planta 2004, 220, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardossi, A.; Falossi, F.; Malorgio, F.; Incrocci, L.; Bellocchi, G. Empirical models of macronutrient uptake in melon plants grown in recirculating nutrient solution culture. J. Plant. Nutr. 2005, 27, 1261–1280. [Google Scholar] [CrossRef]
- Usherwood, N.R. The role of potassium in crop quality. Potassium Agric. 1985, 489–513. [Google Scholar]
- Ashley, M.K.; Grant, M.; Grabov, A. Plant responses to potassium deficiencies: A role for potassium transport proteins. J. Exp. Bot. 2006, 57, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton, K.; Cook, R.L. Potassium in plant nutrition. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1954; Volume 6, pp. 253–303. ISBN 0065-2113. [Google Scholar]
- Karimi, B.; Terrat, S.; Dequiedt, S.; Saby, N.P.A.; Horrigue, W.; Lelièvre, M.; Nowak, V.; Jolivet, C.; Arrouays, D.; Wincker, P. Biogeography of soil bacteria and archaea across France. Sci. Adv. 2018, 4, eaat1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of root colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, F.S.; Herrera, P.G.; Chiquito, G.M.; Morales, M.F.; Castro, A.P.; Paulo, P.L. Relationship between microbial community and environmental conditions in a constructed wetland system treating greywater. Ecol. Eng. 2019, 139, 105581. [Google Scholar] [CrossRef]
- Choi, M.-J.; Bae, J.-Y.; Kim, K.-Y.; Kang, H.; Cha, C.-J. Brevibacillus fluminis sp. nov., isolated from sediment of estuarine wetland. Int. J. Syst. Evol. Microbiol. 2010, 60, 1595–1599. [Google Scholar] [CrossRef]
- You, C.; Zhang, C.; Kong, F.; Feng, C.; Wang, J. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl–mancozeb on bacterial communities in tobacco rhizospheric soil. Ecol. Eng. 2016, 91, 119–125. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- An, P.; Inanaga, S.; Lux, A.; Li, X.J.; Ali, M.E.K.; Matsui, T.; Sugimoto, Y. Effects of salinity and relative humidity on two melon cultivars differing in salt tolerance. Biol. Plant. 2002, 45, 409–415. [Google Scholar] [CrossRef]
- Aulakh, M.S.; Wassmann, R.; Bueno, C.; Kreuzwieser, J.; Rennenberg, H. Characterization of root exudates at different growth stages of ten rice (Oryza sativa L.) cultivars. Plant Biol. 2001, 3, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Growth Stage | Treat.1 | Treat.2 | Treat.3 | Treat.4 | Treat.5 | Treat.6 | |
|---|---|---|---|---|---|---|---|---|
| 1 | Vegetative | 36 | 36 | 27 | 27 | 27 | 27 | |
| 620 | 620 | 465 | 465 | 465 | 465 | |||
| K | 195 | 195 | 146 | 146 | 146 | 146 | ||
| Mg | 50 | 25 | 50 | 25 | 50 | 25 | ||
| 2 | Flowering and pollination I + flowering and pollination M | 36 | 36 | 27 | 27 | 27 | 27 | |
| 620 | 620 | 465 | 465 | 465 | 465 | |||
| K | 195 | 195 | 244 | 244 | 244 | 244 | ||
| Mg | 50 | 25 | 50 | 25 | 50 | 25 | ||
| 3 | fruiting I + fruiting M + Harvesting | 36 | 36 | 36 | 36 | 36 | 36 | |
| 620 | 620 | 620 | 620 | 620 | 620 | |||
| K | 195 | 195 | 48 | 48 | 0 | 0 | ||
| Mg | 50 | 25 | 50 | 25 | 50 | 25 | ||
| All measurements are in mg/L | ||||||||
| Treatments | Weight | Shape Index | Flesh Thickness | Sugar Content | Ascorbic Acid |
|---|---|---|---|---|---|
| (kg) | (cm) | (Brix) | (mg·L−1) | ||
| Treat. 1 | 1.56 ± 0.06 | 0.92 ± 0.06 | 3.92 ± 0.24 | 11.21 ± 1.63 * | 155 ± 32.06 |
| Treat. 2 | 1.84 ± 0.27 * | 0.98 ± 0.02 | 4.05 ± 0.31 | 9.11 ± 0.67 * | 91.50 ± 39.98 |
| Treat. 3 | 1.85 ± 0.27 * | 1.01 ± 0.06 | 4.19 ± 0.21 | 11.35 ± 1.88 | 112.50 ± 44.37 |
| Treat. 4 | 1.79 ± 0.13 | 1.04 ± 0.03 | 4.10 ± 0.15 | 10.95 ± 0.77 | 136.00 ± 46.04 |
| Treat. 5 | 1.63 ± 0.20 | 1.00 ± 0.05 | 4.00 ± 0.16 | 10.82 ± 0.60 | 120.33 ± 39.95 |
| Treat. 6 | 1.51 ± 0.09 * | 1.00 ± 0.04 | 4.00 ± 0.17 | 11.00 ± 1.38 | 132.17 ± 21.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-P.; Lin, C.-M.; Mukhtar, H.; Lo, H.-F.; Ko, M.-C.; Wang, S.-J. Temporal Variability in the Rhizosphere Bacterial and Fungal Community Structure in the Melon Crop Grown in a Closed Hydroponic System. Agronomy 2021, 11, 719. https://doi.org/10.3390/agronomy11040719
Lin Y-P, Lin C-M, Mukhtar H, Lo H-F, Ko M-C, Wang S-J. Temporal Variability in the Rhizosphere Bacterial and Fungal Community Structure in the Melon Crop Grown in a Closed Hydroponic System. Agronomy. 2021; 11(4):719. https://doi.org/10.3390/agronomy11040719
Chicago/Turabian StyleLin, Yu-Pin, Chiao-Ming Lin, Hussnain Mukhtar, Hsiao-Feng Lo, Min-Chun Ko, and Shu-Jen Wang. 2021. "Temporal Variability in the Rhizosphere Bacterial and Fungal Community Structure in the Melon Crop Grown in a Closed Hydroponic System" Agronomy 11, no. 4: 719. https://doi.org/10.3390/agronomy11040719
APA StyleLin, Y. -P., Lin, C. -M., Mukhtar, H., Lo, H. -F., Ko, M. -C., & Wang, S. -J. (2021). Temporal Variability in the Rhizosphere Bacterial and Fungal Community Structure in the Melon Crop Grown in a Closed Hydroponic System. Agronomy, 11(4), 719. https://doi.org/10.3390/agronomy11040719