
Effects on Crop Development, Yields and Chemical Composition of Celeriac (Apium graveolens L. var. rapaceum) Cultivated Underneath an Agrivoltaic System



Abstract
:1. Introduction
2. Material & Methods
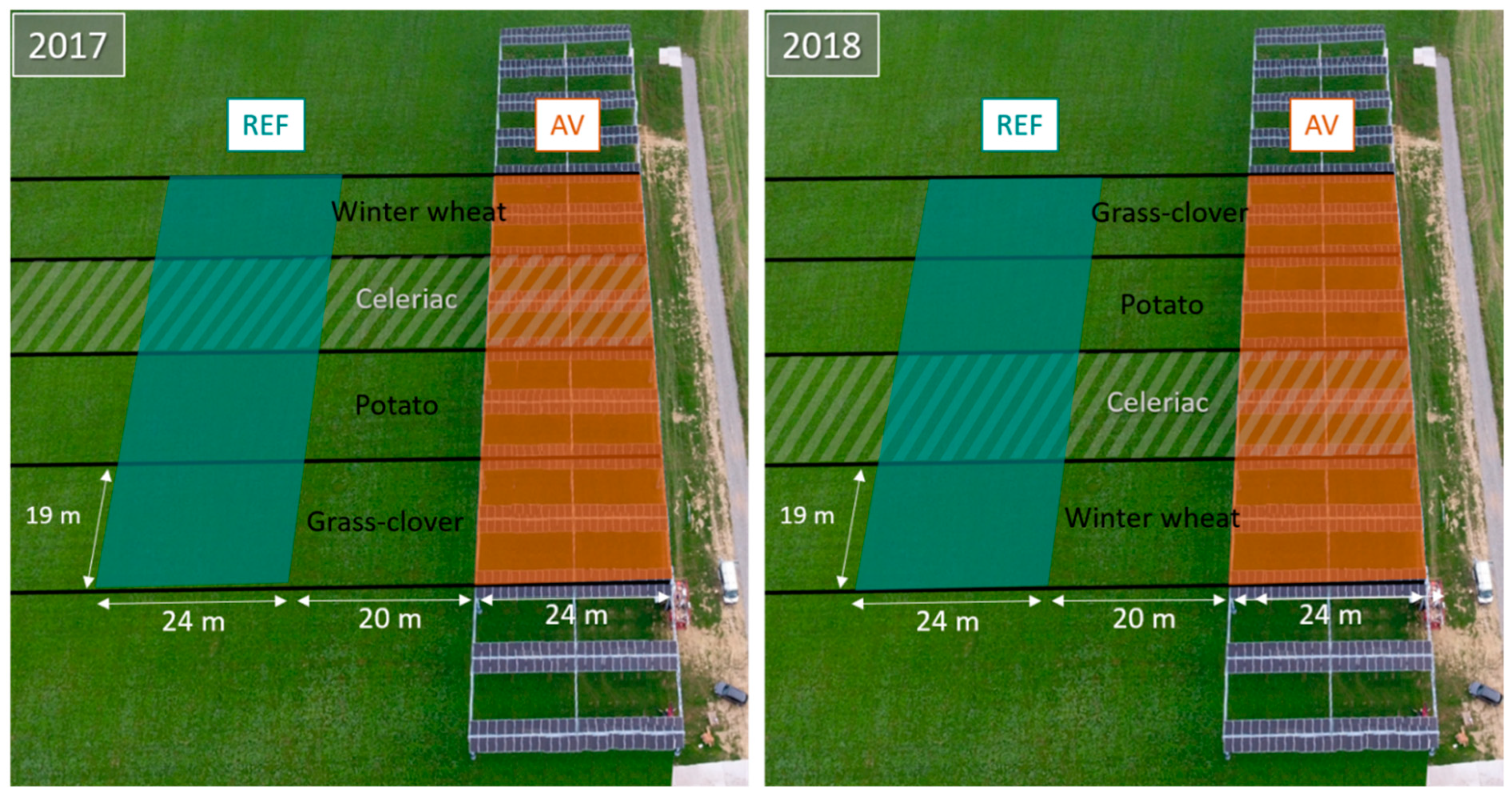
2.1. Site Description & Field Experiment
2.2. Microclimate
2.3. Crop Monitoring & Harvest
2.4. Analysis of Chemical Composition
2.5. Statistical Analysis
3. Results & Discussion
3.1. Microclimate
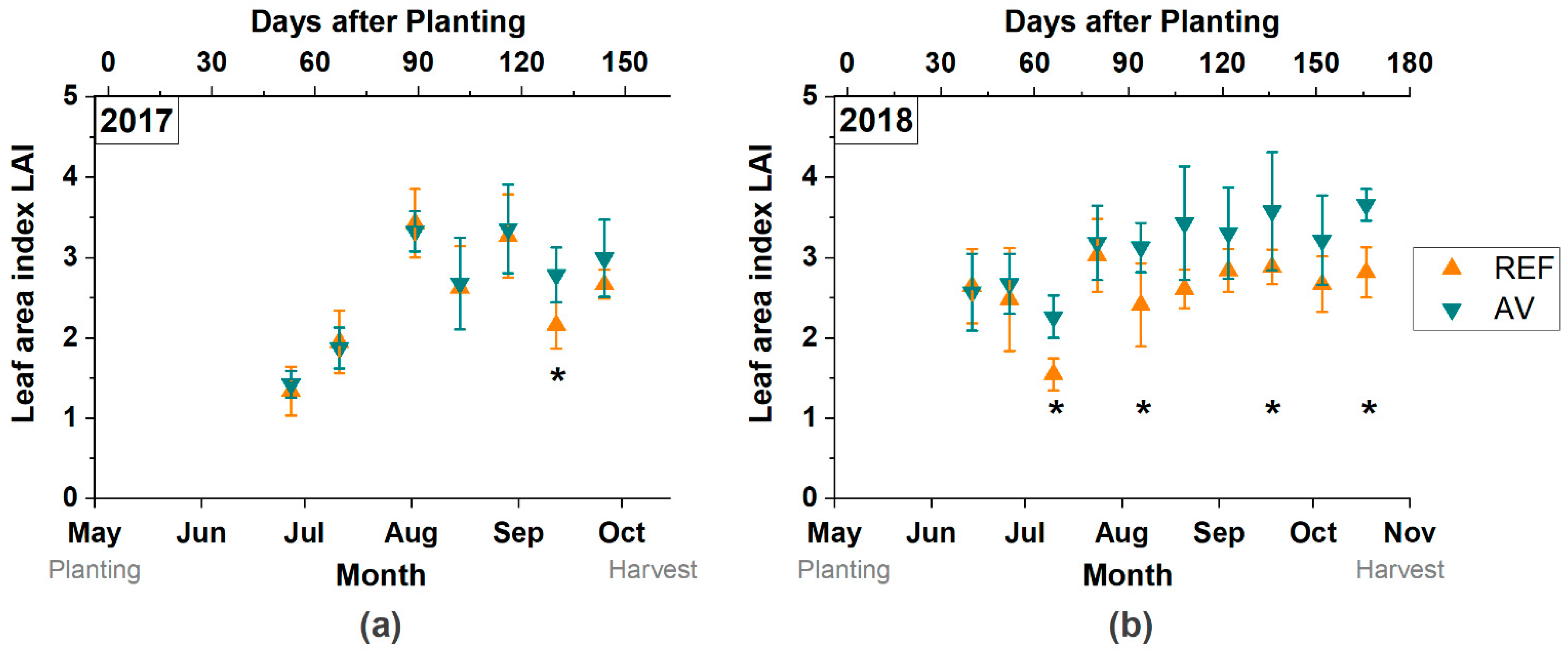
3.2. Crop Development
3.3. Bulb Yields and Yield Components
3.4. Chemical Composition
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schindele, S.; Trommsdorff, M.; Schlaak, A.; Obergfell, T.; Bopp, G.; Reise, C.; Braun, C.; Weselek, A.; Bauerle, A.; Högy, P.; et al. Implementation of agrophotovoltaics: Techno-economic analysis of the price-performance ratio and its policy implications. Appl. Energy 2020, 265, 114737. [Google Scholar] [CrossRef]
- Weselek, A.; Ehmann, A.; Zikeli, S.; Lewandowski, I.; Schindele, S.; Högy, P. Agrophotovoltaic systems: Applications, challenges, and opportunities. A review. Agron. Sustain. Dev. 2019, 39, 545. [Google Scholar] [CrossRef]
- Dupraz, C.; Marrou, H.; Talbot, G.; Dufour, L.; Nogier, A.; Ferard, Y. Combining solar photovoltaic panels and food crops for optimising land use: Towards new agrivoltaic schemes. Renew. Energy 2011, 36, 2725–2732. [Google Scholar] [CrossRef]
- Dinesh, H.; Pearce, J.M. The potential of agrivoltaic systems. Renew. Sustain. Energy Rev. 2016, 54, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Valle, B.; Simonneau, T.; Sourd, F.; Pechier, P.; Hamard, P.; Frisson, T.; Ryckewaert, M.; Christophe, A. Increasing the total productivity of a land by combining mobile photovoltaic panels and food crops. Appl. Energy 2017, 206, 1495–1507. [Google Scholar] [CrossRef]
- Gorjian, S.; Minaee, S.; M.Mirchegini, L.; Trommsdorff, M.; Shamshiri, R. Applications of solar PV systems in agricultural automation and robotics. In Photovoltaic Solar Energy Conversion; Academic Press: Cambridge, MA, USA, 2020; pp. 191–235. ISBN 978-0-12-819610-6. [Google Scholar]
- Barron-Gafford, G.A.; Pavao-Zuckerman, M.A.; Minor, R.L.; Sutter, L.F.; Barnett-Moreno, I.; Blackett, D.T.; Thompson, M.; Dimond, K.; Gerlak, A.K.; Nabhan, G.P.; et al. Agrivoltaics provide mutual benefits across the food–energy–water nexus in drylands. Nat. Sustain. 2019, 2, 848–855. [Google Scholar] [CrossRef]
- Marrou, H.; Wery, J.; Dufour, L.; Dupraz, C. Productivity and radiation use efficiency of lettuces grown in the partial shade of photovoltaic panels. Eur. J. Agron. 2013, 44, 54–66. [Google Scholar] [CrossRef]
- Weselek, A.; Bauerle, A.; Hartung, J.; Zikeli, S.; Lewandowski, I.; Högy, P. Microclimate, crop development and harvestable crop yield of grass-clover, potatoes and winter wheat in an agrivoltaic system under organic management in south-west Germany. Agron. Sustain. Dev. 2021. under review. [Google Scholar]
- Sekiyama, T.; Nagashima, A. Solar Sharing for Both Food and Clean Energy Production: Performance of Agrivoltaic Systems for Corn, A Typical Shade-Intolerant Crop. Environments 2019, 6, 65. [Google Scholar] [CrossRef] [Green Version]
- Homma, M.; Doi, T.; Yoshida, Y. A Field Experiment and the Simulation on Agrivoltaic-systems regarding to Rice in a Paddy Field. J. Jpn. Soc. Energy Resour. 2016, 37, 23–31. [Google Scholar] [CrossRef]
- Marrou, H.; Dufour, L.; Wery, J. How does a shelter of solar panels influence water flows in a soil–crop system? Eur. J. Agron. 2013, 50, 38–51. [Google Scholar] [CrossRef]
- International Federation of Organic Agriculture Movements. Principles of Organic Agriculture. Available online: https://www.ifoam.bio/sites/default/files/2020-03/poa_english_web.pdf (accessed on 6 February 2021).
- Trommsdorff, M.; Kang, J.; Reise, C.; Schindele, S.; Bopp, G.; Ehmann, A.; Weselek, A.; Högy, P.; Obergfell, T. Combining food and energy production: Design of an agrivoltaic system applied in arable and vegetable farming in Germany. Renew. Sustain. Energy Rev. 2021, 140, 110694. [Google Scholar] [CrossRef]
- Pascaris, A.S.; Schelly, C.; Pearce, J.M. A First Investigation of Agriculture Sector Perspectives on the Opportunities and Barriers for Agrivoltaics. Agronomy 2020, 10, 1885. [Google Scholar] [CrossRef]
- Eurostat. The Fruit and Vegetable Sector in the EU—A Statistical Overview. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php/The_fruit_and_vegetable_sector_in_the_EU_-_a_statistical_overview (accessed on 2 April 2021).
- Lim, T.K. Apium graveolens var. rapaceum. In Edible Medicinal and Non Medicinal Plants: Volume 9, Modified Stems, Roots, Bulbs; Lim, T.K., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 367–373. ISBN 978-94-017-9511-1. [Google Scholar]
- Hadley, P.; Fordham, R. Vegetables of Temperate Climates. Miscellaneous Root Crops. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 5948–5951. ISBN 978-0-12-227055-0. [Google Scholar]
- Destatis. Anbauflächen und Erntemengen von Ökologisch Angebautem Gemüse. Available online: https://www.destatis.de/DE/Themen/Branchen-Unternehmen/Landwirtschaft-Forstwirtschaft-Fischerei/Obst-Gemuese-Gartenbau/Tabellen/oekologisches-gemuese.html (accessed on 11 February 2020).
- Feller, C.; Bleiholder, H.; Buhr, L.; Hack, H.; Hess, M.; Klose, R. Phänologische Entwicklungsstadien von Gemüsepflanzen: II. Fruchtgemüse und Hülsenfrüchte. In Compendium of Growth Stage Identification Keys for Mono- and Dicotyledonous Plants, Extended BBCH Scale; German Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany, 1995; Volume 47, pp. 109–111. [Google Scholar]
- LTZ Augustenberg. Agrarmeterologie Baden-Württemberg. Available online: www.wetter-bw.de (accessed on 7 February 2021).
- VDLUFA. 10.8.1.2 Mikrowellenbeheizter Druckaufschluss, 8. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- European Committee for Standardization. Animal Feeding Stuffs—Methods of Sampling and Analysis—Determination of Calcium, Sodium, Phosphorus, Magnesium, Potassium, Sulphur, Iron, Zinc, Copper, Manganese and Cobalt after Pressure Digestion by ICP-AES; EN 15621:2017-10; European Committee for Standardization: Brussels, Belgium, 2017. [Google Scholar]
- VDLUFA. 17.9.1 Bestimmung von Ausgewählten Elementen in Pflanzen Sowie in Grund- und Mischfuttermitteln Mittels Messenspektromie mit Induktiv Gekoppeltem Plasma, 8. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- VDLUFA. 2.2.2.2 Bestimmung von Nitrat in Pflanzlichem Material Mittels Ionenchromatographie; Darmstadt, Band VII Umweltanalytik; VDLUFA: Darmstadt, Germany, 1983. [Google Scholar]
- VDLUFA. 2.2.2.3 Bestimmung des Gehaltes von Extrahierbarem Jod in Futtermitteln Mittels Induktiv Gekoppeltem Plasma und Massenspektrometrie (ICP-MS); Darmstadt, Band VII Umweltanalytik; VDLUFA: Darmstadt, Germany, 1983. [Google Scholar]
- VDLUFA. 4.1.2 Bestimmung von Rohprotein mittels DUMAS-Verbrennungsmethode, 5. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2004. [Google Scholar]
- Regulation (EC) No 152/2009. Laying down the Methods of Sampling and Analysis for the Official Control of Feed. Annex III: Methods of Analysis to Control the Composition of Feed Materials and Compounds Feed; CR (EC) No. 152/2009 III; European Union: Brussels, Belgium, 2009. [Google Scholar]
- VDLUFA. 6.5.1 Bestimmung der Neutral-Detergenzien-Faser nach Amylasebehandlung (aNDF) Sowie Nach Amylasebehandlung und Veraschung (aNDFom), 8. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- VDLUFA. 6.5.2 Bestimmung der Säure-Detergenzien-Faser (ADF) und der Säure-Detergenzien-Faser nach Veraschung (ADFom), 8. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- VDLUFA. 6.5.3 Bestimmung des Säure-Detergenzien-Lignins (ADL), 8. Ergänzung; Darmstadt, Band III Futtermittel; VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- Marrou, H.; Guilioni, L.; Dufour, L.; Dupraz, C.; Wery, J. Microclimate under agrivoltaic systems: Is crop growth rate affected in the partial shade of solar panels? Agric. For. Meteorol. 2013, 177, 117–132. [Google Scholar] [CrossRef]
- Amaducci, S.; Yin, X.; Colauzzi, M. Agrivoltaic systems to optimise land use for electric energy production. Appl. Energy 2018, 220, 545–561. [Google Scholar] [CrossRef]
- McMaster, G.S.; Morgan, J.A.; Willis, W.O. Effects of Shading on Winter Wheat Yield, Spike Characteristics, and Carbohydrate Allocation1. Crop Sci. 1987, 27, 967–973. [Google Scholar] [CrossRef]
- Li, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Effects of shading on morphology, physiology and grain yield of winter wheat. Eur. J. Agron. 2010, 33, 267–275. [Google Scholar] [CrossRef]
- Kuruppuarachchi, D.S.P. Intercropped potato (Solanum spp.) Effect of shade on growth and tuber yield in the northwestern regosol belt of Sri Lanka. Field Crops Res. 1990, 25, 61–72. [Google Scholar] [CrossRef]
- Gommers, C.M.M.; Visser, E.J.W.; Onge, K.R.S.; Voesenek, L.A.C.J.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant adaptation to dynamically changing environment: The shade avoidance response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- United Nations. UNECE Standard FFV-59 Concerning the Marketing and Commercial Quality Control of Root and Tubercle Vegetables. Available online: https://www.unece.org/fileadmin/DAM/trade/agr/standard/fresh/FFV-Std/English/59_RootandTubercleVeg.pdf (accessed on 13 December 2020).
- Hole, C.C.; Thomas, T.H.; Barnes, A.; Scott, P.A.; Rankin, W.E.F. Dry Matter Distribution between Shoot and Storage Root of Carrot, Parsnip, Radish and Red Beet. Ann. Bot. 1984, 53, 625–631. [Google Scholar] [CrossRef]
- Suojala, T. Growth of and partitioning between shoot and storage root of carrot in a northern climate. AFSci 2000, 9. [Google Scholar] [CrossRef]
- Hole, C.C.; Morris, G.E.L.; Cowper, A.S. Distribution of dry matter between shoot and storage root of field-grown carrots. I. Onset of differences between cultivars. J. Hortic. Sci. 1987, 62, 335–341. [Google Scholar] [CrossRef]
- Elamri, Y.; Cheviron, B.; Lopez, J.-M.; Dejean, C.; Belaud, G. Water budget and crop modelling for agrivoltaic systems: Application to irrigated lettuces. Agric. Water Manag. 2018, 208, 440–453. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, J.; Yang, Z.; Xin, G.; Yuan, J.; Xin, J.; Huang, C. Genotype variations in accumulation of cadmium and lead in celery (Apium graveolens L.) and screening for low Cd and Pb accumulative cultivars. Front. Environ. Sci. Eng. 2013, 7, 85–96. [Google Scholar] [CrossRef]
- Regulation (EC) 1881/2006. Setting Maximum Levels for Certain Contaminants in Foodstuffs; CR (EC) No 1881/2006; European Union: Brussels, Belgium, 2006. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of Drought on Nutrient Uptake and Assimilation in Vegetable Crops. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. ISBN 978-3-642-32653-0. [Google Scholar]
- Heath-Pagliuso, S.; Matlin, S.A.; Fang, N.; Thompson, R.H.; Rappaport, L. Stimulation of furanocoumarin accumulation in celery and celeriac tissues by Fusarium oxysporum f. sp. Apii. Phytochemistry 1992, 31, 2683–2688. [Google Scholar] [CrossRef]
- Schulzová, V.; Babička, L.; Hajšlová, J. Furanocoumarins in celeriac from different farming systems: A 3-year study. J. Sci. Food Agric. 2012, 92, 2849–2854. [Google Scholar] [CrossRef] [PubMed]
- Zobel, A.M.; Brown, S.A. Influence of low-intensity ultraviolet radiation on extrusion of furanocoumarins to the leaf surface. J. Chem. Ecol. 1993, 19, 939–952. [Google Scholar] [CrossRef]
- Popova, M.; Stoyanova, A.; Valyovska-Popova, N.; Bankova, V.; Peev, D. A new coumarin and total phenolic and flavonoids content of Bulgarian celeriac. Izvestiya Khimiya Bulgarska Akademiya Naukite 2014, 46, 88–93. [Google Scholar]
- Leclerc, J.; Miller, M.L.; Joliet, E.; Rocquelin, G. Vitamin and Mineral Contents of Carrot and Celeriac Grown under Mineral or Organic Fertilization. Biol. Agric. Hortic. 1991, 7, 339–348. [Google Scholar] [CrossRef]
- Kaiser, A.; Hartmann, K.I.; Kammerer, D.R.; Carle, R. Evaluation of the effects of thermal treatments on color, polyphenol stability, enzyme activities and antioxidant capacities of innovative pasty celeriac (Apium graveolens L. var. rapaceum (Mill.) DC.) products. Eur. Food Res. Technol. 2013, 237, 353–365. [Google Scholar] [CrossRef]
- MacLeod, G.; Ames, J.M. Volatile components of celery and celeriac. Phytochemistry 1989, 28, 1817–1824. [Google Scholar] [CrossRef]
- Maynard, D.N.; Barker, A.V.; Minotti, P.L.; Peck, N.H. Nitrate Accumulation in Vegetables. In Advances in Agronomy; Brady, N.C., Ed.; Academic Press: Cambridge, MA, USA, 1976; pp. 71–118. ISBN 0065-2113. [Google Scholar]
- Bruning-Fann, C.; Kaneene, J. The effects of nitrate, nitrite and N-nitroso compounds on human health: A review. Vet. Hum. Toxicol. 1994, 35, 521–538. [Google Scholar]
- Schuthan, W.; Bengtsson, B.; Bosund, I.; Hylmö, B. Nitrate accumulation in spinach. Plant Foods Hum. Nutr. 1967, 14, 317–330. [Google Scholar] [CrossRef]
- Wright, M.J.; Davison, K.L. Nitrate Accumulation In Crops And Nitrate Poisoning In Animals. In Advances in Agronomy; Norman, A.G., Ed.; Academic Press: Cambridge, MA, USA, 1964; pp. 197–247. ISBN 0065-2113. [Google Scholar]
- Wojciechowska, R.; Siwek, P. The effect of shading on nitrate metabolism in stalks and blades of celery leaves (Apium graveolens L. var. dulce). Folia Hortic. 2006, 18, 25–35. [Google Scholar]
- Derolez, J.; Vulsteke, G. Accumulation of nitrate: A cultivar-linked property with celeriac (Apium graveolens L.var. rapaceum). Plant Foods Hum. Nutr. 1985, 35, 375–378. [Google Scholar] [CrossRef]
- Dambrauskienė, E.; Maročkiene, N.; Viskelis, P.; Rubinskiene, M. Evaluation of productivity and biochemical composition of edible root celery. Acta Hortic. 2009, 830, 115–119. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
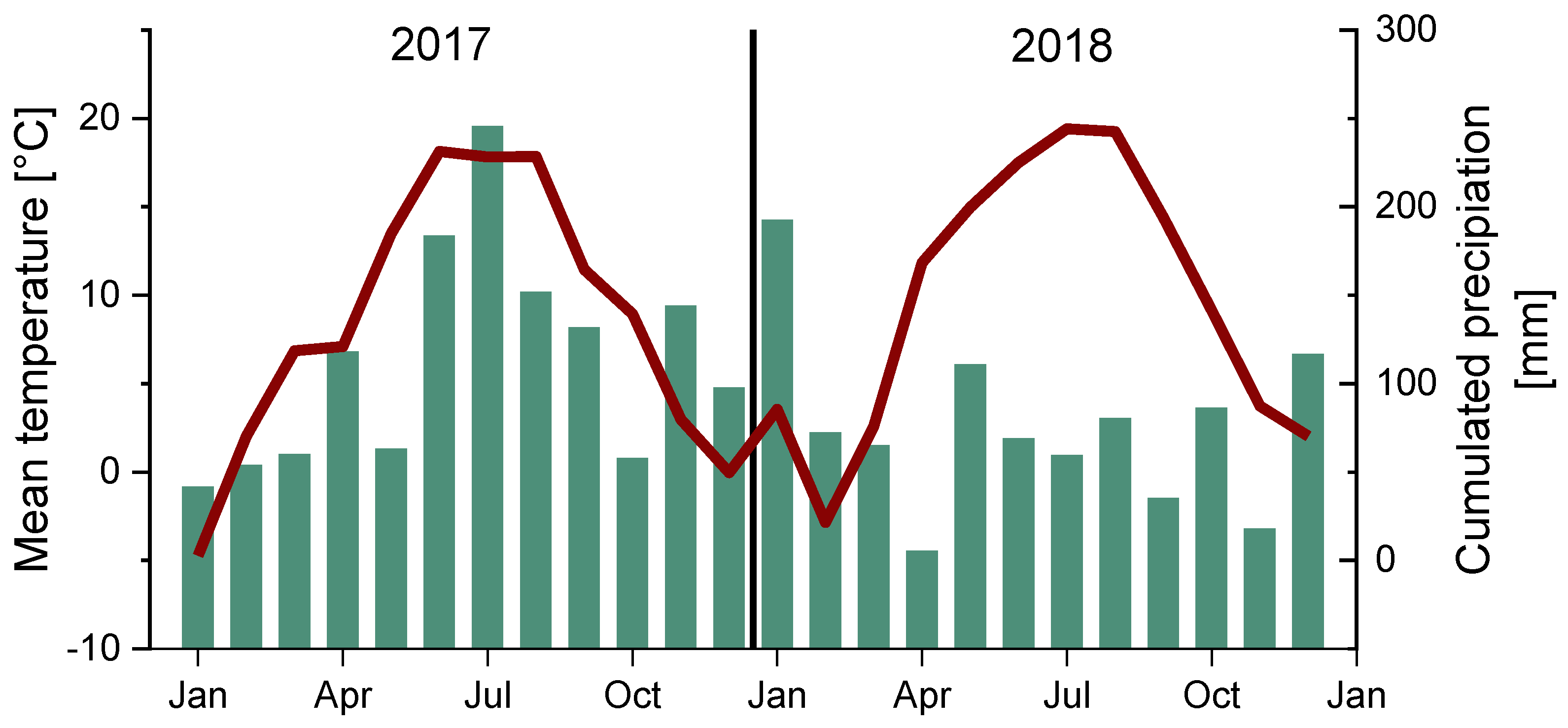{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Air Temperature [°C] | Humidity [%] | Soil Temperature [°C] | Soil Moisture [%] | PPFD [μmol/m²s] | ||
|---|---|---|---|---|---|---|
| 2017 | REF | 8.7 | 79.1 | 18.4 | 25.2 | 469.4 |
| AV | 8.7 | 81.9 | 17.2 | 27.1 | 336.7 | |
| 2018 | REF | 10.4 | 71.6 | 19.2 | 20.9 | 497.9 |
| AV | 10.4 | 74.4 | 17.8 | 17.8 | 344.5 |
| Treatment | [% DM] | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crude Protein | Crude Fat | aNDFom | ADFom | ADL | C | S | Ca | K | Mg | Na | P | Cl | |
| 2017 | |||||||||||||
| AV | 10a | 2.25a | - | - | - | 39.5a | 0.09a | 0.31a | 4.09a | 0.2 | 0.31a | 0.58a | 0.08a |
| REF | 9.4a | 2.18a | - | - | - | 40.1b | 0.09a | 0.34b | 3.9a | 0.21 | 0.31a | 0.59a | 0.08a |
| 2018 | |||||||||||||
| AV | 11.7b | 2.78b | 13.3 | 9.5a | 2.06 | 40.7c | 0.08b | 0.28c | 2.19b | 0.19 | 0.16b | 0.33b | 0.05b |
| REF | 12.1b | 2.83b | 16.1 | 10.5b | 3.0 | 41.1d | 0.08ab | 0.3ac | 2.25b | 0.18 | 0.22c | 0.3b | 0.06ab |
| SEM | 0.196 | 0.075 | 1.348 | 0.072 | 0.327 | 0.149 | 0.002 | 0.005 | 0.076 | 0.01 | 0.018 | 0.01 | 0.009 |
| p-value | |||||||||||||
| Year | <0.0001 | <0.0001 | - | - | - | <0.0001 | 0.0612 | 0.0001 | <0.0001 | 0.082 | <0.0001 | <0.0001 | 0.0152 |
| Trt | 0.6643 | 0.8713 | 0.1918 | 0.0002 | 0.0985 | 0.0048 | 1.0 | 0.0021 | 0.4225 | 0.9 | 0.0844 | 0.3628 | 0.7760 |
| Trt∗Year | 0.0397 | 0.4254 | - | - | - | 0.6843 | 0.3166 | 0.2009 | 0.1221 | 0.7071 | 0.1065 | 0.1832 | 0.2694 |
| Treatment | [ppm DM] | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Al | B | Ba | Cu | Fe | Mn | Zn | Si | Cd | Co | Cr | Ni | Mo | Pb | Se | I | |
| 2018 | ||||||||||||||||
| AV | 3.16a | 33a | 8.04a | 16a | 21.3a | 36.4a | 29.3a | <150 | 0.67a | <0.02 | 0.23a | 0.92a | 0.05a | 0.08a | 0.03 | <0.50 |
| REF | 3.02a | 30.8a | 11.1b | 17.4a | 21a | 60.2b | 31.2a | <150 | 1.33b | <0.02 | 0.08ab | 1.49b | 0.04a | 0.11b | 0.03 | <0.50 |
| 2017 | ||||||||||||||||
| AV | 1.28b | 27b | 2.47c | 14.1b | 30b | 42.1ac | 25.9b | <150 | 0.5a | <0.02 | 0.02b | 1.19a | <0.02b | 0.06c | 0.02 | <0.50 |
| REF | 2.7ab | 24.6b | 3.8c | 12.8b | 28.4b | 48.1c | 25.7b | <150 | 0.96c | <0.02 | 0.03b | 2.15c | <0.02b | 0.1ab | 0.02 | <0.50 |
| SEM | 0.53 | 1.02 | 0.876 | 0.515 | 1.404 | 2.462 | 1.069 | - | 0.079 | - | 0.059 | 0.089 | 0.003 | 0.006 | 0.007 | - |
| p-value | ||||||||||||||||
| Year | 0.0626 | 0.0001 | <0.0001 | <0.0001 | 0.0001 | 0.2261 | 0.0019 | - | 0.0064 | - | 0.0484 | 0.0004 | <0.0001 | 0.0029 | 0.163 | - |
| Trt | 0.2705 | 0.0467 | 0.0299 | 0.8685 | 0.6533 | 0.0001 | 0.4452 | - | <0.0001 | - | 0.2875 | <0.0001 | 0.2872 | 0.0002 | 0.9202 | - |
| Trt∗Year | 0.1818 | 0.8578 | 0.3487 | 0.0225 | 0.6657 | 0.0046 | 0.3385 | - | 0.231 | - | 0.1996 | 0.0521 | 0.2872 | 0.4191 | 0.7898 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weselek, A.; Bauerle, A.; Zikeli, S.; Lewandowski, I.; Högy, P. Effects on Crop Development, Yields and Chemical Composition of Celeriac (Apium graveolens L. var. rapaceum) Cultivated Underneath an Agrivoltaic System. Agronomy 2021, 11, 733. https://doi.org/10.3390/agronomy11040733
Weselek A, Bauerle A, Zikeli S, Lewandowski I, Högy P. Effects on Crop Development, Yields and Chemical Composition of Celeriac (Apium graveolens L. var. rapaceum) Cultivated Underneath an Agrivoltaic System. Agronomy. 2021; 11(4):733. https://doi.org/10.3390/agronomy11040733
Chicago/Turabian StyleWeselek, Axel, Andrea Bauerle, Sabine Zikeli, Iris Lewandowski, and Petra Högy. 2021. "Effects on Crop Development, Yields and Chemical Composition of Celeriac (Apium graveolens L. var. rapaceum) Cultivated Underneath an Agrivoltaic System" Agronomy 11, no. 4: 733. https://doi.org/10.3390/agronomy11040733
APA StyleWeselek, A., Bauerle, A., Zikeli, S., Lewandowski, I., & Högy, P. (2021). Effects on Crop Development, Yields and Chemical Composition of Celeriac (Apium graveolens L. var. rapaceum) Cultivated Underneath an Agrivoltaic System. Agronomy, 11(4), 733. https://doi.org/10.3390/agronomy11040733