
Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants



{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
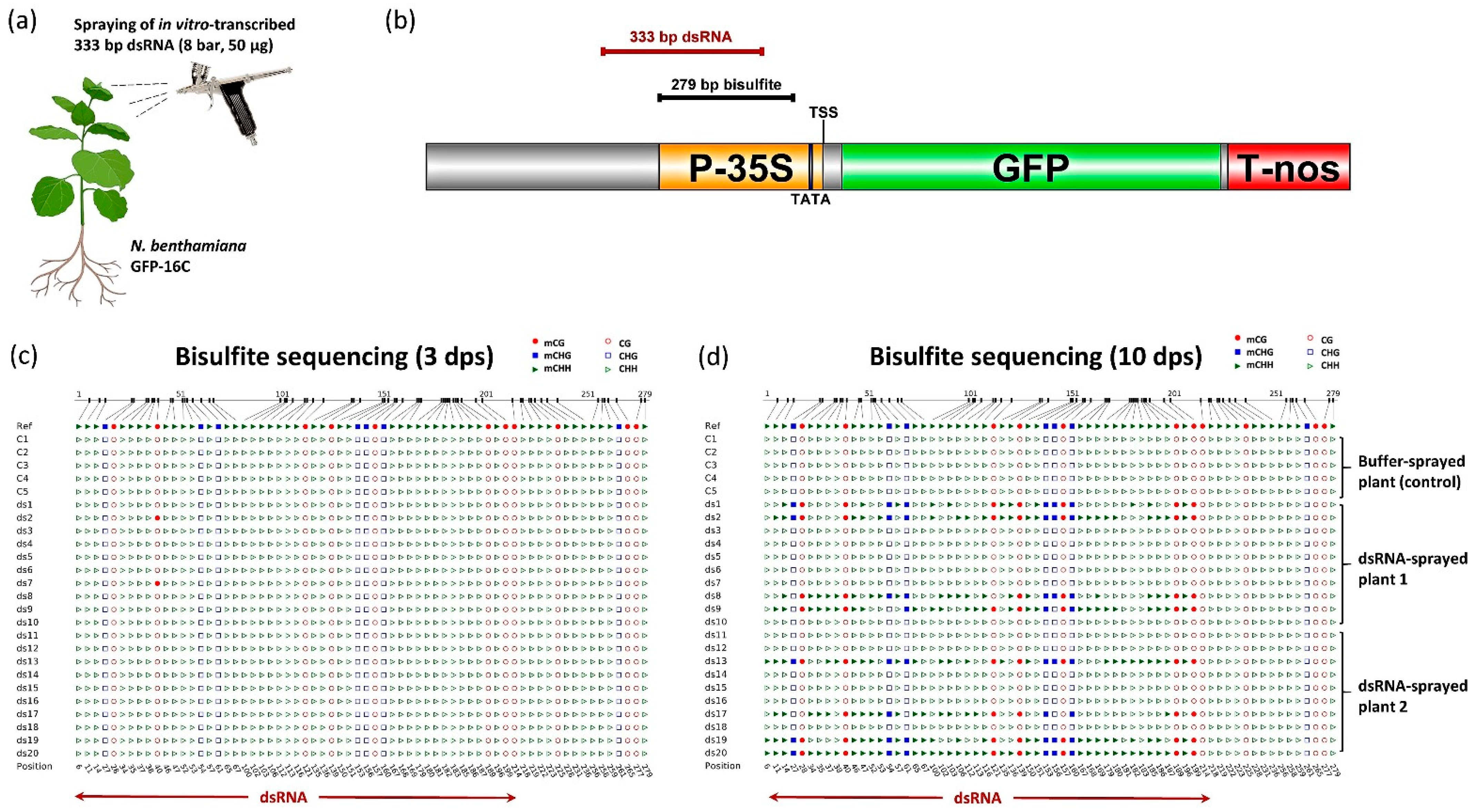
2.1. In Vitro Transcription of dsRNA
2.2. High-Pressure Spraying of dsRNA
2.3. Genomic DNA Extraction
2.4. Bisulfite Sequencing
3. Results and Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Wassenegger, M.; Heimes, S.; Riedel, L.; Sänger, H.L. RNA-directed de novo methylation of genomic sequences in plants. Cell 1994, 76, 567–576. [Google Scholar] [CrossRef]
- Jones, L.; Hamilton, A.J.; Voinnet, O.; Thomas, C.L.; Maule, A.J.; Baulcombe, D.C. RNA–DNA Interactions and DNA Methylation in Post-Transcriptional Gene Silencing. Plant Cell 1999, 11, 2291–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sijen, T.; Vijn, I.; Rebocho, A.; van Blokland, R.; Roelofs, D.; Mol, J.N.; Kooter, J.M. Transcriptional and posttranscriptional gene silencing are mechanistically related. Curr. Biol. 2001, 11, 436–440. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, A.F.; Matthew, L.; Smith, N.; Curtin, S.J.; Dedic-Hagan, J.; Ellacott, G.; Watson, J.M.; Wang, M.; Brosnan, C.; Carroll, B.J.; et al. RNA interference-inducing hairpin RNAs in plants act through the viral defence pathway. EMBO Rep. 2006, 7, 1168–1175. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [Green Version]
- Baumberger, N.; Baulcombe, D.C. Arabidopsis ARGONAUTE1 is an RNA Slicer that selectively recruits microRNAs and short interfering RNAs. Proc. Natl. Acad. Sci. USA 2005, 102, 11928–11933. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.W.-L.; Zilberman, D.; Xie, Z.; Johansen, L.K.; Carrington, J.C.; Jacobsen, S.E. RNA Silencing Genes Control de Novo DNA Methylation. Science 2004, 303, 1336. [Google Scholar] [CrossRef]
- Zilberman, D.; Cao, X.; Jacobsen, S.E. ARGONAUTE4 Control of Locus-Specific siRNA Accumulation and DNA and Histone Methylation. Science 2003, 299, 716–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, M.A.; Kanno, T.; Matzke, A.J. RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants. Annu. Rev. Plant Biol. 2015, 66, 243–267. [Google Scholar] [CrossRef]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nat. Rev. Genet. 2014, 15, 394–408. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J. DNA methylation in plants: Mechanisms and tools for targeted manipulation. New Phytol. 2020, 227, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Aufsatz, W.; Zilberman, D.; Mette, M.; Huang, M.S.; Matzke, M.; Jacobsen, S.E. Role of the DRM and CMT3 Methyltransferases in RNA-Directed DNA Methylation. Curr. Biol. 2003, 13, 2212–2217. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-W.; Shao, C.-R.; Zhang, C.-J.; Zhou, J.-X.; Zhang, S.-W.; Li, L.; Chen, S.; Huang, H.-W.; Cai, T.; He, X.-J. The SET Domain Proteins SUVH2 and SUVH9 Are Required for Pol V Occupancy at RNA-Directed DNA Methylation Loci. PLoS Genet. 2014, 10, e1003948. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.-L.; Zhang, G.; Tang, K.; Li, J.; Yang, L.; Huang, H.; Zhang, H.; Zhu, J.-K. Dicer-independent RNA-directed DNA methylation in Arabidopsis. Cell Res. 2016, 26, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalakouras, A.; Papadopoulou, K.K. Epigenetic Modifications: An Unexplored Facet of Exogenous RNA Application in Plants. Plants 2020, 9, 673. [Google Scholar] [CrossRef]
- Dalakouras, A.; Wassenegger, M. Revisiting RNA-directed DNA methylation. RNA Biol. 2013, 10, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, J.; Bischof, S.; Wang, H.; Feng, S.; Lee, T.-F.; Teng, C.; Chen, X.; Park, S.Y.; Liu, L.; Gallego-Bartolome, J.; et al. A One Precursor One siRNA Model for Pol IV-Dependent siRNA Biogenesis. Cell 2015, 163, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelissier, T.; Thalmeir, S.; Kempe, D.; Sanger, H.L.; Wassenegger, M. Heavy de novo methylation at symmetrical and non-symmetrical sites is a hallmark of RNA-directed DNA methylation. Nucleic Acids Res. 1999, 27, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aufsatz, W.; Mette, M.F.; Matzke, A.J.M.; Matzke, M. The role of MET1 in RNA-directed de novoand maintenance methylation of CG dinucleotides. Plant Mol. Biol. 2004, 54, 793–804. [Google Scholar] [CrossRef]
- Dadami, E.; Dalakouras, A.; Zwiebel, M.; Krczal, G.; Wassenegger, M. An endogene-resembling transgene is resistant to DNA methylation and systemic silencing. RNA Biol. 2014, 11, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Křížová, K.; Depicker, A.; Kovařík, A. Epigenetic switches of tobacco transgenes associate with transient redistribution of histone marks in callus culture. Epigenetics 2013, 8, 666–676. [Google Scholar] [CrossRef] [Green Version]
- Okano, Y.; Miki, D.; Shimamoto, K. Small interfering RNA (siRNA) targeting of endogenous promoters induces DNA methylation, but not necessarily gene silencing, in rice. Plant J. 2008, 53, 65–77. [Google Scholar] [CrossRef]
- Zilberman, D.; Cao, X.; Johansen, L.K.; Xie, Z.; Carrington, J.C.; Jacobsen, S.E. Role of Arabidopsis ARGONAUTE4 in RNA-Directed DNA Methylation Triggered by Inverted Repeats. Curr. Biol. 2004, 14, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Mette, M.F.; Aufsatz, W.; Van Der Winden, J.; Matzke, M.A.; Matzke, A.J.M. Transcriptional silencing and promoter methylation triggered by double-stranded RNA. EMBO J. 2000, 19, 5194–5201. [Google Scholar] [CrossRef] [PubMed]
- Mette, M.; Van Der Winden, J.; Matzke, M.; Matzke, A. Production of aberrant promoter transcripts contributes to methylation and silencing of unlinked homologous promoters in trans. EMBO J. 1999, 18, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalakouras, A.; Moser, M.; Zwiebel, M.; Krczal, G.; Hell, R.; Wassenegger, M. A hairpin RNA construct residing in an intron efficiently triggered RNA-directed DNA methylation in tobacco. Plant J. 2009, 60, 840–851. [Google Scholar] [CrossRef]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.L.; Papadopoulou, K. Genetically Modified Organism-Free RNA Interference: Exogenous Application of RNA Molecules in Plants. Plant Physiol. 2020, 182, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Aleynova, O.A.; Kalachev, A.V.; Suprun, A.R.; Ogneva, Z.V.; Kiselev, K.V. Induction of Transgene Suppression in Plants via External Application of Synthetic dsRNA. Int. J. Mol. Sci. 2019, 20, 1585. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A Perspective on RNAi-Based Biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Taning, C.N.T.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Mezzetti, B.; Sabbadini, S.; Sorteberg, H.; Sweet, J.; Ventura, V.; et al. RNA-based biocontrol compounds: Current status and perspectives to reach the market. Pest Manag. Sci. 2020, 76, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.J.; Statham, A.; Stirzaker, C.; Molloy, P.L.; Frommer, M. DNA methylation: Bisulphite modification and analysis. Nat. Protoc. 2006, 1, 2353–2364. [Google Scholar] [CrossRef] [PubMed]
- Hetzl, J.; Foerster, A.M.; Raidl, G.; Scheid, O.M. CyMATE: A new tool for methylation analysis of plant genomic DNA after bisulphite sequencing. Plant J. 2007, 51, 526–536. [Google Scholar] [CrossRef]
- Voinnet, O.; Baulcombe, D.C. Systemic signalling in gene silencing. Nat. Cell Biol. 1997, 389, 553. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Wassenegger, M.; McMillan, J.N.; Cardoza, V.; Maegele, I.; Dadami, E.; Runne, M.; Krczal, G.; Wassenegger, M. Induction of Silencing in Plants by High-Pressure Spraying of In vitro-Synthesized Small RNAs. Front. Plant Sci. 2016, 7, 1327. [Google Scholar] [CrossRef] [Green Version]
- Philips, J.G.; Naim, F.; Lorenc, M.T.; Dudley, K.J.; Hellens, R.P.; Waterhouse, P.M. The widely used Nicotiana benthamiana 16c line has an unusual T-DNA integration pattern including a transposon sequence. PLoS ONE 2017, 12, e0171311. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, K.V.; Suprun, A.R.; Aleynova, O.A.; Ogneva, Z.V.; Dubrovina, A.S. Physiological Conditions and dsRNA Application Approaches for Exogenously induced RNA Interference in Arabidopsis thaliana. Plants 2021, 10, 264. [Google Scholar] [CrossRef]
- Uslu, V.V.; Bassler, A.; Krczal, G.; Wassenegger, M. High-Pressure-Sprayed Double Stranded RNA Does Not Induce RNA Interference of a Reporter Gene. Front. Plant Sci. 2020, 11, 534391. [Google Scholar] [CrossRef]
- Daxinger, L.; Kanno, T.; Bucher, E.; Van Der Winden, J.; Naumann, U.; Matzke, A.J.M.; Matzke, M. A stepwise pathway for biogenesis of 24-nt secondary siRNAs and spreading of DNA methylation. EMBO J. 2008, 28, 48–57. [Google Scholar] [CrossRef]
- Pontes, O.; Vitins, A.; Ream, T.S.; Hong, E.; Pikaard, C.S.; Costa-Nunes, P. Intersection of Small RNA Pathways in Arabidopsis thaliana Sub-Nuclear Domains. PLoS ONE 2013, 8, e65652. [Google Scholar] [CrossRef] [Green Version]
- Boutla, A.; Kalantidis, K.; Tavernarakis, N.; Tsagris, M.; Tabler, M. Induction of RNA interference in Caenorhabditis elegans by RNAs derived from plants exhibiting post-transcriptional gene silencing. Nucleic Acids Res. 2002, 30, 1688–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirnaz, S.; Batley, J. Epigenetics: Potentials and Challenges in Crop Breeding. Mol. Plant 2019, 12, 1309–1311. [Google Scholar] [CrossRef]
- Varotto, S.; Tani, E.; Abraham, E.; Krugman, T.; Kapazoglou, A.; Melzer, R.; Radanović, A.; Miladinović, D. Epigenetics: Possible applications in climate-smart crop breeding. J. Exp. Bot. 2020, 71, 5223–5236. [Google Scholar] [CrossRef]
- Matzke, M.; Aufsatz, W.; Kanno, T.; Daxinger, L.; Papp, I.; Mette, M.; Matzke, A.J. Genetic analysis of RNA-mediated transcriptional gene silencing. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2004, 1677, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M. The Role of the RNAi Machinery in Heterochromatin Formation. Cell 2005, 122, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Fischer, U.; Kuhlmann, M.; Pecinka, A.; Schmidt, R.; Mette, M.F. Local DNA features affect RNA-directed transcriptional gene silencing and DNA methylation. Plant J. 2008, 53, 1–10. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.M.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Samarsky, D.A.; Fournier, M.J.; Singer, R.H.; Bertrand, E. The snoRNA box C/D motif directs nucleolar targeting and also couples snoRNA synthesis and localization. EMBO J. 1998, 17, 3747–3757. [Google Scholar] [CrossRef]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for Plant Improvement: Current Knowledge and Modeling Avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R. Breeding wheat for increased potential yield: Contrasting ideas from Donald and Fasoulas, and the case for early generation selection under nil competition. Field Crop. Res. 2020, 252, 107782. [Google Scholar] [CrossRef]
- Eamens, A.; Wang, M.-B.; Smith, N.A.; Waterhouse, P.M. RNA Silencing in Plants: Yesterday, Today, and Tomorrow. Plant Physiol. 2008, 147, 456–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigan, A.M.; Haug-Collet, K.; Clapp, J. Transcriptional silencing of heterologous anther promoters in maize: A genetic method to replace detasseling for seed production. Plant Reprod. 2014, 27, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Cigan, A.M.; Unger-Wallace, E.; Haug-Collet, K. Transcriptional gene silencing as a tool for uncovering gene function in maize. Plant J. 2005, 43, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Heilersig, B.H.J.B.; Loonen, A.E.H.M.; Janssen, E.M.; Wolters, A.-M.A.; Visser, R.G.F. Efficiency of transcriptional gene silencing of GBSSI in potato depends on the promoter region that is used in an inverted repeat. Mol. Genet. Genom. 2006, 275, 437–449. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalakouras, A.; Ganopoulos, I. Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants. Agronomy 2021, 11, 789. https://doi.org/10.3390/agronomy11040789
Dalakouras A, Ganopoulos I. Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants. Agronomy. 2021; 11(4):789. https://doi.org/10.3390/agronomy11040789
Chicago/Turabian StyleDalakouras, Athanasios, and Ioannis Ganopoulos. 2021. "Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants" Agronomy 11, no. 4: 789. https://doi.org/10.3390/agronomy11040789
APA StyleDalakouras, A., & Ganopoulos, I. (2021). Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants. Agronomy, 11(4), 789. https://doi.org/10.3390/agronomy11040789