
Characterization of Volatile Organic Compounds in ‘Rossa di Tropea’ Onion by Means of Headspace Solid-Phase Microextraction Gas Chromatography–Mass Spectrometry (HS/SPME GC–MS) and Sensory Analysis




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Setup
2.2. Phytosanitary Status Assessment
2.3. VOCs Analysis by HS/SPME GC-MS
2.4. Statistical Analysis
2.5. Sensory Analysis
3. Results and Discussion
3.1. VOCs Analysis by HS/SPME GC-MS
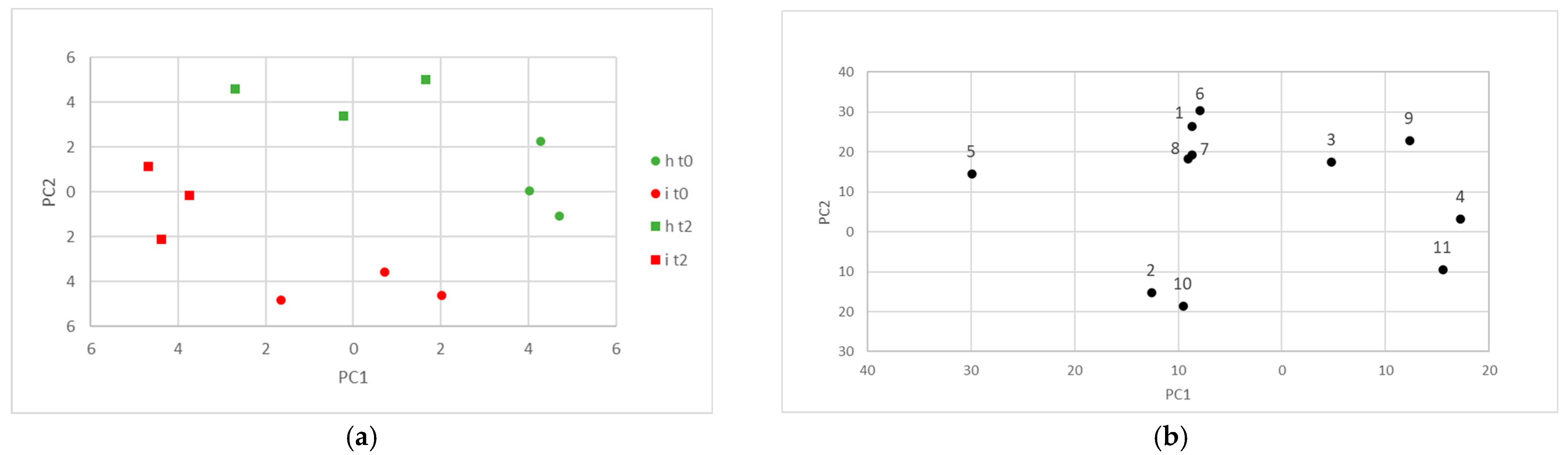
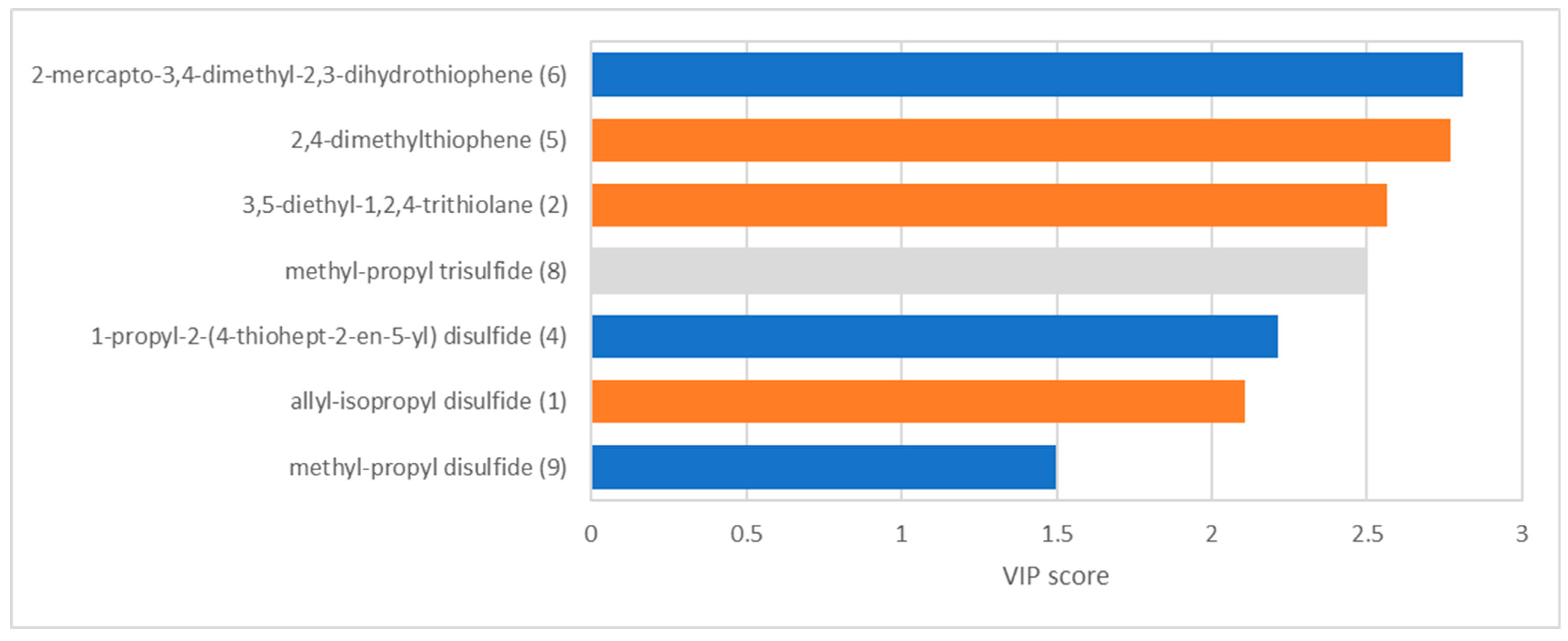
3.2. Statistical Analysis
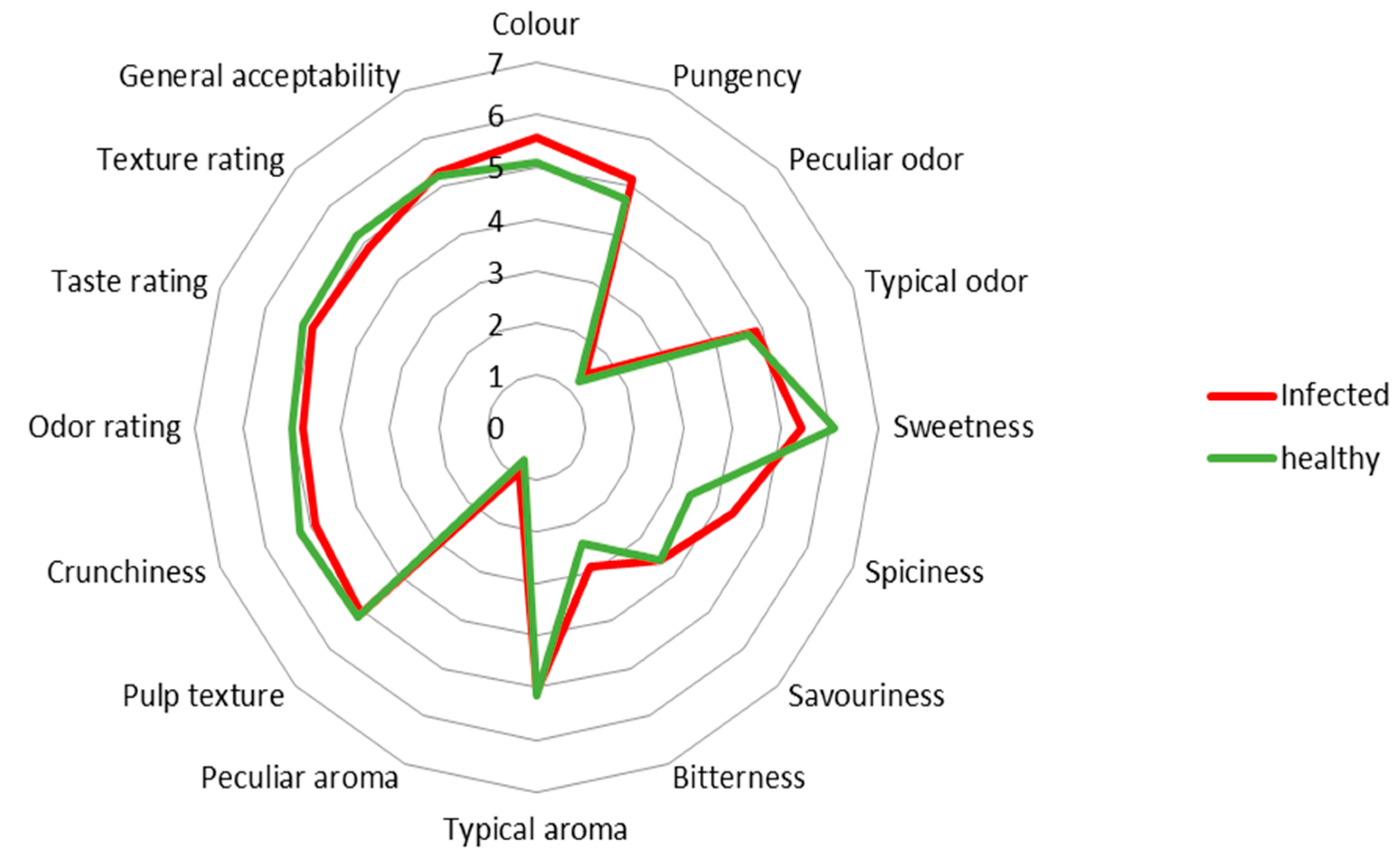
3.3. Sensory Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an Evolved Concept of Landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Wendin, K.; Mustafa, A.; Ortman, T.; Gerhardt, K. Consumer Awareness, Attitudes and Preferences towards Heritage Cereals. Foods 2020, 9, 742. [Google Scholar] [CrossRef] [PubMed]
- Giupponi, L.; Pedrali, D.; Leoni, V.; Rodari, A.; Giorgi, A. The Analysis of Italian Plant Agrobiodiversity Databases Reveals That Hilly and Sub-Mountain Areas Are Hotspots of Herbaceous Landraces. Diversity 2021, 13, 70. [Google Scholar] [CrossRef]
- Teuber, R.; Dolgopolova, I.; Nordström, J. Some like it organic, some like it purple and some like it ancient: Consumer preferences and WTP for value-added attributes in whole grain bread. Food Qual. Prefer. 2016, 52, 244–254. [Google Scholar] [CrossRef]
- Longin, C.F.H.; Würschum, T. Back to the Future—Tapping into Ancient Grains for Food Diversity. Trends Plant Sci. 2016, 21, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Amodeo, V.; Statti, G.; Conforti, F. Biological Properties and Bioactive Components of Allium cepa L.: Focus on Potential Benefits in the Treatment of Obesity and Related Comorbidities. Molecules 2018, 24, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Luca, A.; Edelenbos, M. Emission of volatile organic compounds from yellow onion (Allium cepa L.) bulbs during storage. J. Food Sci. Technol. 2019, 56, 2940–2948. [Google Scholar] [CrossRef]
- Mondy, N.; Duplat, D.; Christides, J.; Arnault, I.; Auger, J. Aroma analysis of fresh and preserved onions and leek by dual solid-phase microextraction–liquid extraction and gas chromatography–mass spectrometry. J. Chromatogr. A 2002, 963, 89–93. [Google Scholar] [CrossRef]
- Rose, P.; Whiteman, M.; Moore, P.K.; Zhu, Y.Z. Bioactive S-alk(en)yl cysteine sulfoxide metabolites in the genus Allium: The chemistry of potential therapeutic agents. Nat. Prod. Rep. 2005, 22, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Tsuge, N.; Tomotake, M.; Nagatome, Y.; Sawada, H.; Nagata, T.; Kumagai, H. An onion enzyme that makes the eyes water. Nat. Cell Biol. 2002, 419, 685. [Google Scholar] [CrossRef]
- Keusgen, M. Volatile Compounds of the Genus Allium L. (Onions). In Volatile Sulfur Compounds in Food; American Chemical Society: Washington, DC, USA, 2011; Volume 1068, pp. 183–214. [Google Scholar]
- Gîtin, L.; Dinică, R.; Neagu, C.; Dumitrascu, L. Sulfur compounds identification and quantification from Allium spp. fresh leaves. J. Food Drug Anal. 2014, 22, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Kusano, M.; Kobayashi, M.; Iizuka, Y.; Fukushima, A.; Saito, K. Unbiased profiling of volatile organic compounds in the headspace of Allium plants using an in-tube extraction device. BMC Res. Notes 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Parkin, K.L. In vitro biogeneration of pure thiosulfinates and propanethial-S-oxide. J. Agric. Food Chem. 2000, 48, 6254–6260. [Google Scholar] [CrossRef] [PubMed]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saviano, G.; Paris, D.; Melck, D.; Fantasma, F.; Motta, A.; Iorizzi, M. Metabolite variation in three edible Italian Allium cepa L. by NMR-based metabolomics: A comparative study in fresh and stored bulbs. Metabolomics 2019, 15, 105. [Google Scholar] [CrossRef]
- Kallio, H.; Salorinne, L. Comparison of onion varieties by headspace gas chromatography-mass spectrometry. J. Agric. Food Chem. 1990, 38, 1560–1564. [Google Scholar] [CrossRef]
- Liguori, L.; Adiletta, G.; Nazzaro, F.; Fratianni, F.; di Matteo, M.; Albanese, D. Biochemical, Antioxidant Properties and Antimicrobial Activity of Different Onion Varieties in the Mediterranean Area. J. Food Meas. Charact. 2019, 13, 1232–1241. [Google Scholar]
- Cranshaw, W.; Crowe, F.; Davis, R.; Dugan, F. Infections/Biotic Diseases. In Compendium of Onion and Garlic Diseases and Pests; Schwartz, H., Mohan, S., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2007. [Google Scholar]
- Manglli, A.; Tomassoli, L.; Tiberini, A.; Agosteo, G.E.; Fontana, A.; Pappu, H.R.; Albanese, G. A survey on the infection of Onion yellow dwarf virus and Iris yellow spot tospovirus in seed and bulb productions systems of onion in Calabria, Italy. Eur. J. Plant Pathol. 2020, 156, 767–778. [Google Scholar] [CrossRef]
- Mishra, J.; Srivastava, R.; Trivedi, P.K.; Verma, P.C. Effect of virus infection on the secondary metabolite production and phytohormone biosynthesis in plants. 3 Biotech 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Tungadi, T.; Groen, S.C.; Murphy, A.M.; Pate, A.E.; Iqbal, J.; Bruce, T.J.A.; Cunniffe, N.J.; Carr, J.P. Cucumber mosaic virus and its 2b protein alter emission of host volatile organic compounds but not aphid vector settling in tobacco. Virol. J. 2017, 14, 91. [Google Scholar] [CrossRef] [Green Version]
- Mwando, N.L.; Tamiru, A.; Nyasani, J.O.; Obonyo, M.A.O.; Caulfield, J.C.; Bruce, T.J.A.; Subramanian, S. Maize Chlorotic Mottle Virus Induces Changes in Host Plant Volatiles that Attract Vector Thrips Species. J. Chem. Ecol. 2018, 44, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Fleming-Jones, M.E.; Smith, R.E. Volatile Organic Compounds in Foods: A Five Year Study. J. Agric. Food Chem. 2003, 51, 8120–8127. [Google Scholar] [CrossRef] [PubMed]
- Gouinguené, S.P.; Turlings, T.C. The Effects of Abiotic Factors on Induced Volatile Emissions in Corn Plants. Plant Physiol. 2002, 129, 1296–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Schmidt, N.E.; Gitaitis, R. Detection of onion postharvest diseases by analyses of headspace volatiles using a gas sensor array and GC-MS. LWT 2011, 44, 1019–1025. [Google Scholar] [CrossRef]
- De Lange, E.S.; Rodriguez-Saona, C. Does enhanced nutrient availability increase volatile emissions in cranberry? Plant Signal. Behav. 2019, 14, 1616517. [Google Scholar] [CrossRef] [PubMed]
- Villière, A.; Le Roy, S.; Fillonneau, C.; Guillet, F.; Falquerho, H.; Boussely, S.; Prost, C. Evaluation of aroma profile differences between sué, sautéed, and pan-fried onions using an innovative olfactometric approach. Flavour 2015, 4, 24. [Google Scholar] [CrossRef]
- Choi, S.M.; Lee, N.-J.; Kim, J.-Y.; Lim, S.-T. Volatile composition and sensory characteristics of onion powders prepared by convective drying. Food Chem. 2017, 231, 386–392. [Google Scholar] [CrossRef]
- Prithiviraj, B.; Vikram, A.; Kushalappa, A.; Yaylayan, V. Volatile Metabolite Profiling for the Discrimination of Onion Bulbs Infected by Erwinia carotovora ssp. carotovora, Fusarium oxysporum and Botrytis allii. Eur. J. Plant Pathol. 2004, 110, 371–377. [Google Scholar] [CrossRef]
- Vikram, A.; Hamzehzarghani, H.; Kushalappa, A. Volatile metabolites from the headspace of onion bulbs inoculated with postharvest pathogens as a tool for disease discrimination. Can. J. Plant Pathol. 2005, 27, 194–203. [Google Scholar] [CrossRef]
- Wang, A.; Islam, N.; Johansen, A.; Haapalainen, M.; Latvala, S.; Edelenbos, M. Pathogenic Fusarium oxysporum f. sp. cepae growing inside onion bulbs emits volatile organic compounds that correlate with the extent of infection. Postharvest Biol. Technol. 2019, 152, 19–28. [Google Scholar] [CrossRef]
- Tiberini, A.; Mangano, R.; Micali, G.; Leo, G.; Manglli, A.; Tomassoli, L.; Albanese, G. Onion yellow dwarf virus ∆∆Ct-based relative quantification obtained by using real-time polymerase chain reaction in ‘Rossa di Tropea’ onion. Eur. J. Plant Pathol. 2018, 153, 251–264. [Google Scholar] [CrossRef]
- Araniti, F.; Pantò, S.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Chemical Characterization of Volatile Organic Compounds (VOCs) Through Headspace Solid Phase Micro Extraction (SPME). In Advances in Plant Ecophysiology Techniques; Metzler, J.B., Ed.; Springer: Cham, Switzerland, 2018; pp. 401–417. [Google Scholar]
- Araniti, F.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Allelopatic Potential of Dittrichia viscosa (L.) W. Greuter Mediated by VOCs: A Physiological and Metabolomic Approach. PLoS ONE 2017, 12, e0170161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Szymańska, E.; Saccenti, E.; Smilde, A.K.; Westerhuis, J.A. Double-check: Validation of diagnostic statistics for PLS-DA models in metabolomics studies. Metabolomics 2011, 8, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Brereton, R.G. Consequences of sample size, variable selection, and model validation and optimisation, for predicting classification ability from analytical data. TrAC Trends Anal. Chem. 2006, 25, 1103–1111. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; Van Velzen, E.J.J.; Van Duijnhoven, J.P.M.; Van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Brereton, R.G. Pattern recognition in chemometrics. Chemom. Intell. Lab. Syst. 2015, 149, 90–96. [Google Scholar] [CrossRef]
- Lee, I.; Yang, J. Common Clustering Algorithms. In Comprehensive Chemometrics; Elsevier: Amsterdam, The Nederlands, 2009; pp. 577–618. [Google Scholar]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.1–14.10.91. [Google Scholar] [CrossRef]
- ISO 8586:2012 Sensory Analysis—General Guidelines for the Selection, Training and Monitoring of Selected Assessors and Expert Sensory Assessors; ISO: Geneva, Switzerland, 2012.
- Meilgaard, M.C.; Carr, B.T. Sensory Evaluation Techniques; Chapman and Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Block, E. The Organosulfur Chemistry of the Genus Allium—Implications for the Organic Chemistry of Sulfur. Angew. Chem. Int. Ed. 1992, 31, 1135–1178. [Google Scholar] [CrossRef]
- McCallum, J.; Porter, N.; Searle, B.; Shaw, M.; Bettjeman, B.; McManus, M. Sulfur and nitrogen fertility affects flavour of field-grown onions. Plant Soil 2005, 269, 151–158. [Google Scholar] [CrossRef]
- Wang, A.; Casadei, F.; Johansen, A.; Bukman, H.; Edelenbos, M. Emission of volatile organic compounds from healthy and diseased onions. Acta Hortic. 2016, 1144, 333–340. [Google Scholar] [CrossRef]
- Fernández-Bedmar, Z.; Demyda-Peyrás, S.; Merinas-Amo, T.; Del Río-Celestino, M. Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation. Foods 2019, 8, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekara, A.; Pokluda, R.; Del Vacchio, L.; Somma, S.; Caruso, G.; Agnieszka, S.; Robert, P.; Del, V.L.; Silvano, S.; Gianluca, C. Interactions among genotype, environment and agronomic practices on production and quality of storage onion (Allium cepa L.)—A review. Hortic. Sci. 2017, 44, 21–42. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, R.A. Comparative Distribution of Volatile Aliphatic Disulfides Derived from Fresh and Dehydrated Onions. J. Food Sci. 1968, 33, 298–304. [Google Scholar] [CrossRef]
- Järvenpää, E.P.; Zhang, Z.; Huopalahti, R.; King, J.W. Determination of fresh onion (Allium cepa L.) volatiles by solid phase microextraction combined with gas chromatography-mass spectrometry. Z. Lebensm. Forsch. A 1998, 207, 39–43. [Google Scholar] [CrossRef]
- Løkke, M.M.; Edelenbos, M.; Larsen, E.; Feilberg, A. Investigation of Volatiles Emitted from Freshly Cut Onions (Allium cepa L.) by Real Time Proton-Transfer Reaction-Mass Spectrometry (PTR-MS). Sensors 2012, 12, 16060–16076. [Google Scholar] [CrossRef] [Green Version]
- Block, E.; Naganathan, S.; Putman, D.; Zhao, S.H. Allium chemistry: HPLC analysis of thiosulfinates from onion, garlic, wild garlic (ramsoms), leek, scallion, shallot, elephant (great-headed) garlic, chive, and Chinese chive. Uniquely high allyl to methyl ratios in some garlic samples. J. Agric. Food Chem. 1992, 40, 2418–2430. [Google Scholar] [CrossRef]
- Berger, R. Flavours and Fragrances, Chemistry, Bioprocessing and Sustainability; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Boelens, M.; De Valois, P.J.; Wobben, H.J.; Van der Gen, A. Volatile flavor compounds from onion. J. Agric. Food Chem. 1971, 19, 984–991. [Google Scholar] [CrossRef]
- Cecchi, L.; Ieri, F.; Vignolini, P.; Mulinacci, N.; Romani, A. Characterization of Volatile and Flavonoid Composition of Different Cuts of Dried Onion (Allium cepa L.) by HS-SPME-GC-MS, HS-SPME-GC×GC-TOF and HPLC-DAD. Molecules 2020, 25, 408. [Google Scholar] [CrossRef] [Green Version]
- Block, E.; Shu, H.Z. Onion essential oil chemistry. Cis-and trans-2-mercapto-3,4-dimethyl 2,3-dihydrothiophene from pyrolysis of bis(1-propenyl) disulfide. Tetrahedron Lett. 1990, 31, 4999–5002. [Google Scholar] [CrossRef]
- Eady, C.C.; Kamoi, T.; Kato, M.; Porter, N.G.; Davis, S.; Shaw, M.; Kamoi, A.; Imai, S. Silencing Onion Lachrymatory Factor Synthase Causes a Significant Change in the Sulfur Secondary Metabolite Profile. Plant Physiol. 2008, 147, 2096–2106. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhang, J.; Zhou, Z.; Hua, Z.; Wan, H.; Xie, Y.; Wang, Z.; Deng, L. Analysis of Volatile Compounds and Identification of Characteristic Aroma Components of Toona sinensis (A. Juss.) Roem. Using GC-MS and GC-O. Food Nutr. Sci. 2013, 4, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Ntatsi, G.; Ferreira, I.C.F.R. Long-term storage of onion and the factors that affect its quality: A critical review. Food Rev. Int. 2016, 33, 62–83. [Google Scholar] [CrossRef]
- Sharma, K.; Assefa, A.D.; Ko, E.Y.; Lee, E.T.; Park, S.W. Quantitative analysis of flavonoids, sugars, phenylalanine and tryptophan in onion scales during storage under ambient conditions. J. Food Sci. Technol. 2013, 52, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Lesemann, D.-E. Specific Cytological Alterations in Virus-Infected Plant Cells. In Electron Microscopy of Plant Pathogens; Springer: Berlin/Heidelberg, Germany, 1991; pp. 147–159. [Google Scholar]
- Elena, S.F.; Rodrigo, G. Towards an integrated molecular model of plant–virus interactions. Curr. Opin. Virol. 2012, 2, 719–724. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | OYDV | Diameter t0 (mm) | Diameter t2 (mm) | Weight t0 (g) | Weight t2 (g) |
|---|---|---|---|---|---|
| B2 | absent | 56.8 | n.d. | 239.4 | n.d. |
| C7 | absent | 55.1 | n.d. | 232.2 | n.d. |
| F5 | absent | 55.9 | n.d. | 236.6 | n.d. |
| B21 | absent | 56.2 | 53.9 | 234.6 | 185.1 |
| C29 | absent | 57.8 | 54.2 | 242.1 | 190.2 |
| F22 | absent | 56.4 | 53.2 | 235.8 | 185.3 |
| A3 | present | 58.8 | n.d. | 250.8 | n.d. |
| D6 | present | 57.7 | n.d. | 244.6 | n.d. |
| E4 | present | 58.5 | n.d. | 249.1 | n.d. |
| A23 | present | 57.9 | 52.5 | 244.9 | 156.2 |
| D25 | present | 58.2 | 53.1 | 248.7 | 158.7 |
| E24 | present | 58.0 | 52.9 | 248.9 | 158.8 |
| Entry | Compound | CAS n. |
|---|---|---|
| 1 | allyl-isopropyl disulfide | 67421-85-6 |
| 2 | 3,5-diethyl-1,2,4-trithiolane | 54644-28-9 |
| 3 | 1-nitro-2-prop-2-enylsulfanylpropane | 127865-37-6 |
| 4 | 1-propyl-2-(4-thiohept-2-en-5-yl) disulfide | 143193-11-7 |
| 5 | 2,4-dimethylthiophene | 638-00-6 |
| 6 | 2-mercapto-3,4-dimethyl-2,3-dihydrothiophene | n.d. |
| 7 | acetic acid | 64-19-7 |
| 8 | methyl-propyl trisulfide | 17619-36-2 |
| 9 | methyl-propyl disulfide | 2179-60-4 |
| 10 | Propanethiol | 107-03-9 |
| 11 | propyl disulfide | 629-19-6 |
| Entry | Sampling Time | t0 | t2 | |||
|---|---|---|---|---|---|---|
| Compound | Control | OYDV | Control | OYDV | VOC Ratio | |
| 1 | allyl-isopropyl disulfide | 5.67 ± 0.52 | 6.83 ± 1.09 | 33.15 ± 7.05 ** | 7.93 ± 0.32 ** | 0.38 |
| 2 | 3,5-diethyl 1,2,4-trithiolane | 0.71 ± 0.30 | 1.12 ± 0.77 | 0.27 ± 0.03 ** | 1.07 ± 0.08 ** | 2.24 |
| 3 | 1-nitro-2-prop-2-enylsulfanylpropane | 0.31 ± 0.17 | 0.13 ± 0.06 | 0.33 ± 0.23 | 0.28 ± 0.05 | 0.63 |
| 4 | 1-propyl 2,4-thiohept-2-en-5-yl disulfide | 1.12 ± 0.63 | 0.57 ± 0.30 | 0.37 ± 0.03 ** | 0.12 ± 0.02 ** | 0.46 |
| 5 | 2,4-dimethyl thiophene | 0.05 ± 0.02 | 0.07 ± 0.03 | 0.25 ± 0.01 ** | 0.37 ± 0.03 ** | 1.48 |
| 6 | 2-mercapto-3,4-dimethyl 2,3-dihydrothiophene | 0.27 ± 0.19 | 0.27 ± 0.03 | 1.10 ± 0.04 ** | 0.44 ± 0.05 ** | 0.52 |
| 8 | methyl-propyl trisulfide | 0.29 ± 0.22 | 0.11 ± 0.05 | 2.34 ± 1.94 | 0.24 ± 0.06 | 0.13 |
| 9 | methyl-propyl disulfide | 1.57 ± 0.11 ** | 0.78 ± 0.11 ** | 3.59 ± 1.54 | 0.58 ± 0.09 | 0.26 |
| 10 | propanethiol | 10.97 ± 1.67 * | 16.15 ± 1.63 * | 4.76 ± 2.00 | 15.44 ± 5.49 | 2.01 |
| 11 | propyl-disulfide | 67.39 ± 3.48 | 60.13 ± 6.57 | 51.59 ± 9.94 | 61.11 ± 3.49 | 1.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taglienti, A.; Araniti, F.; Piscopo, A.; Tiberini, A. Characterization of Volatile Organic Compounds in ‘Rossa di Tropea’ Onion by Means of Headspace Solid-Phase Microextraction Gas Chromatography–Mass Spectrometry (HS/SPME GC–MS) and Sensory Analysis. Agronomy 2021, 11, 874. https://doi.org/10.3390/agronomy11050874
Taglienti A, Araniti F, Piscopo A, Tiberini A. Characterization of Volatile Organic Compounds in ‘Rossa di Tropea’ Onion by Means of Headspace Solid-Phase Microextraction Gas Chromatography–Mass Spectrometry (HS/SPME GC–MS) and Sensory Analysis. Agronomy. 2021; 11(5):874. https://doi.org/10.3390/agronomy11050874
Chicago/Turabian StyleTaglienti, Anna, Fabrizio Araniti, Amalia Piscopo, and Antonio Tiberini. 2021. "Characterization of Volatile Organic Compounds in ‘Rossa di Tropea’ Onion by Means of Headspace Solid-Phase Microextraction Gas Chromatography–Mass Spectrometry (HS/SPME GC–MS) and Sensory Analysis" Agronomy 11, no. 5: 874. https://doi.org/10.3390/agronomy11050874
APA StyleTaglienti, A., Araniti, F., Piscopo, A., & Tiberini, A. (2021). Characterization of Volatile Organic Compounds in ‘Rossa di Tropea’ Onion by Means of Headspace Solid-Phase Microextraction Gas Chromatography–Mass Spectrometry (HS/SPME GC–MS) and Sensory Analysis. Agronomy, 11(5), 874. https://doi.org/10.3390/agronomy11050874