
Growth Stage Specific Lighting Spectra Affect Photosynthetic Performance, Growth and Mineral Element Contents in Tomato



Abstract
:1. Introduction
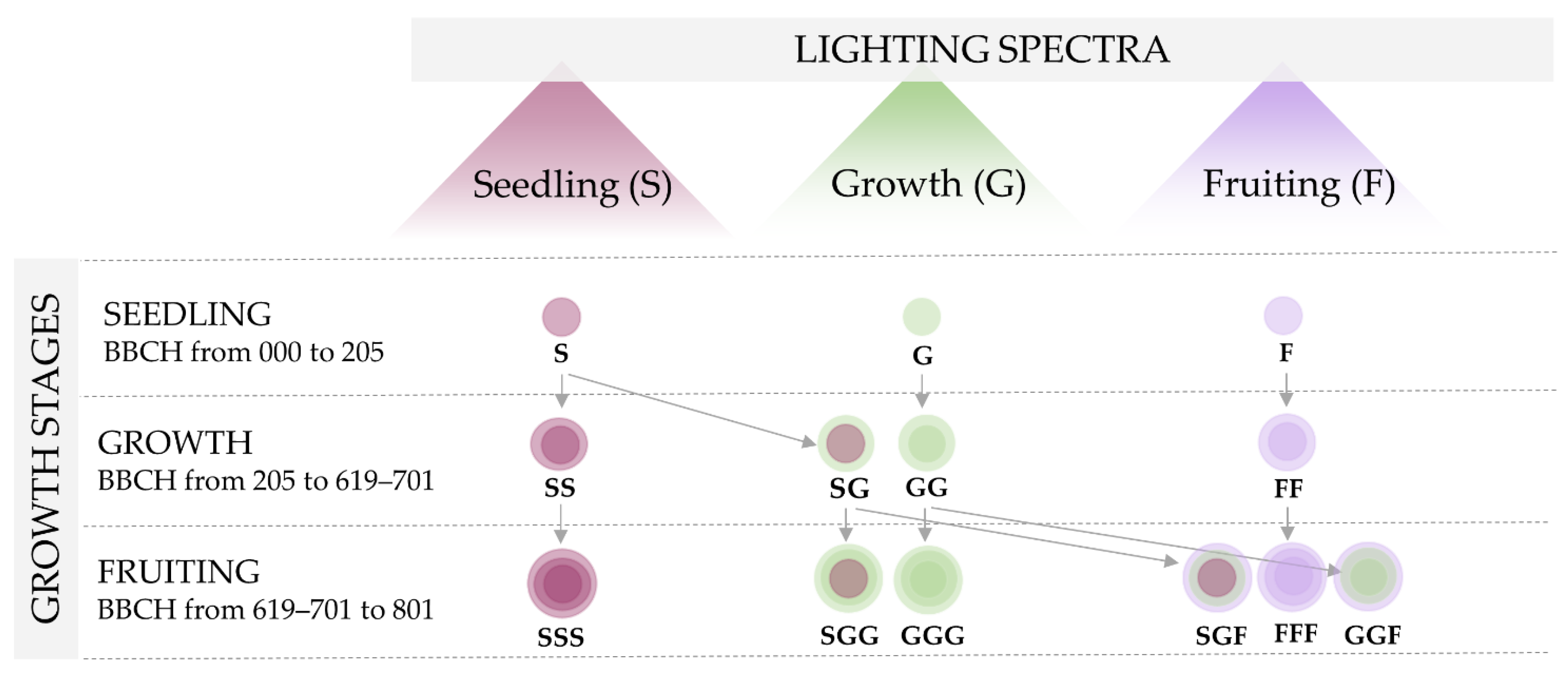
2. Materials and Methods
3. Results
3.1. Photosynthetic Performance
3.2. Growth and Yield Parameters
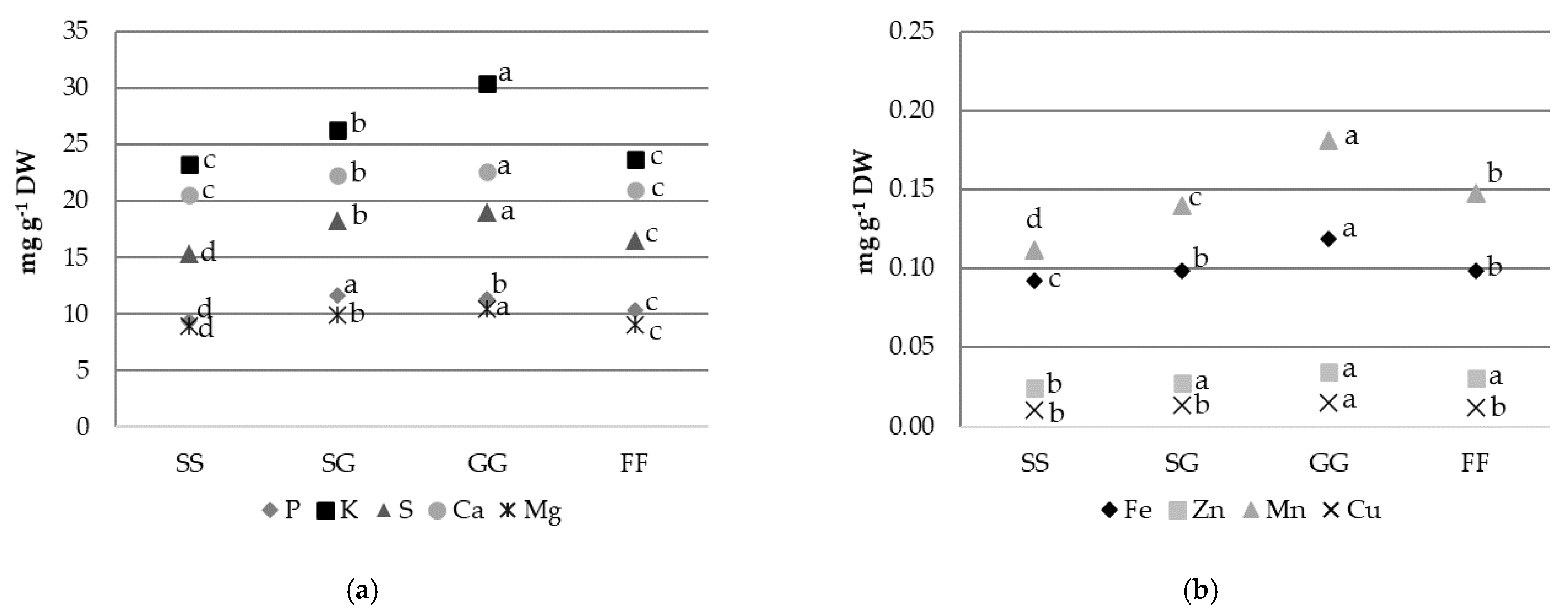
3.3. Mineral Element Contents
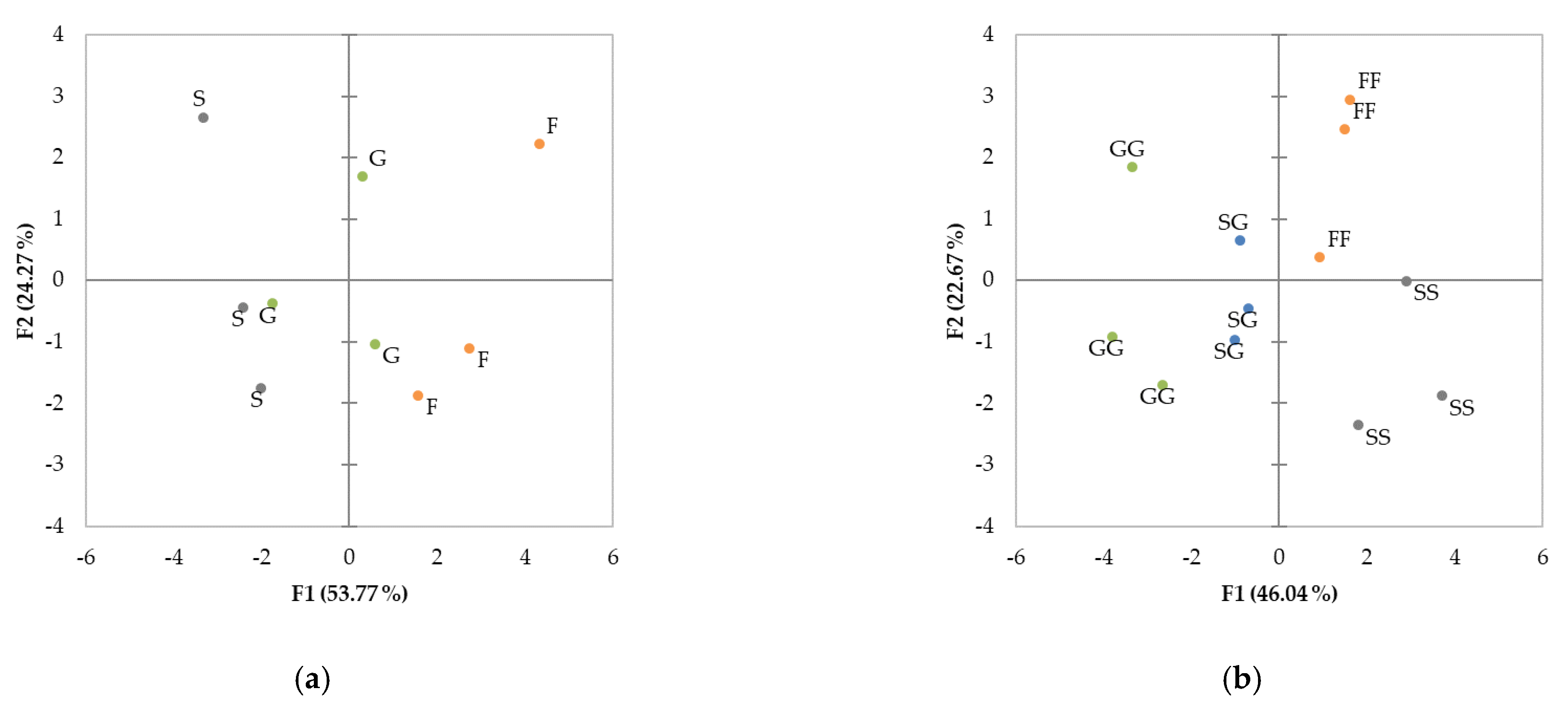
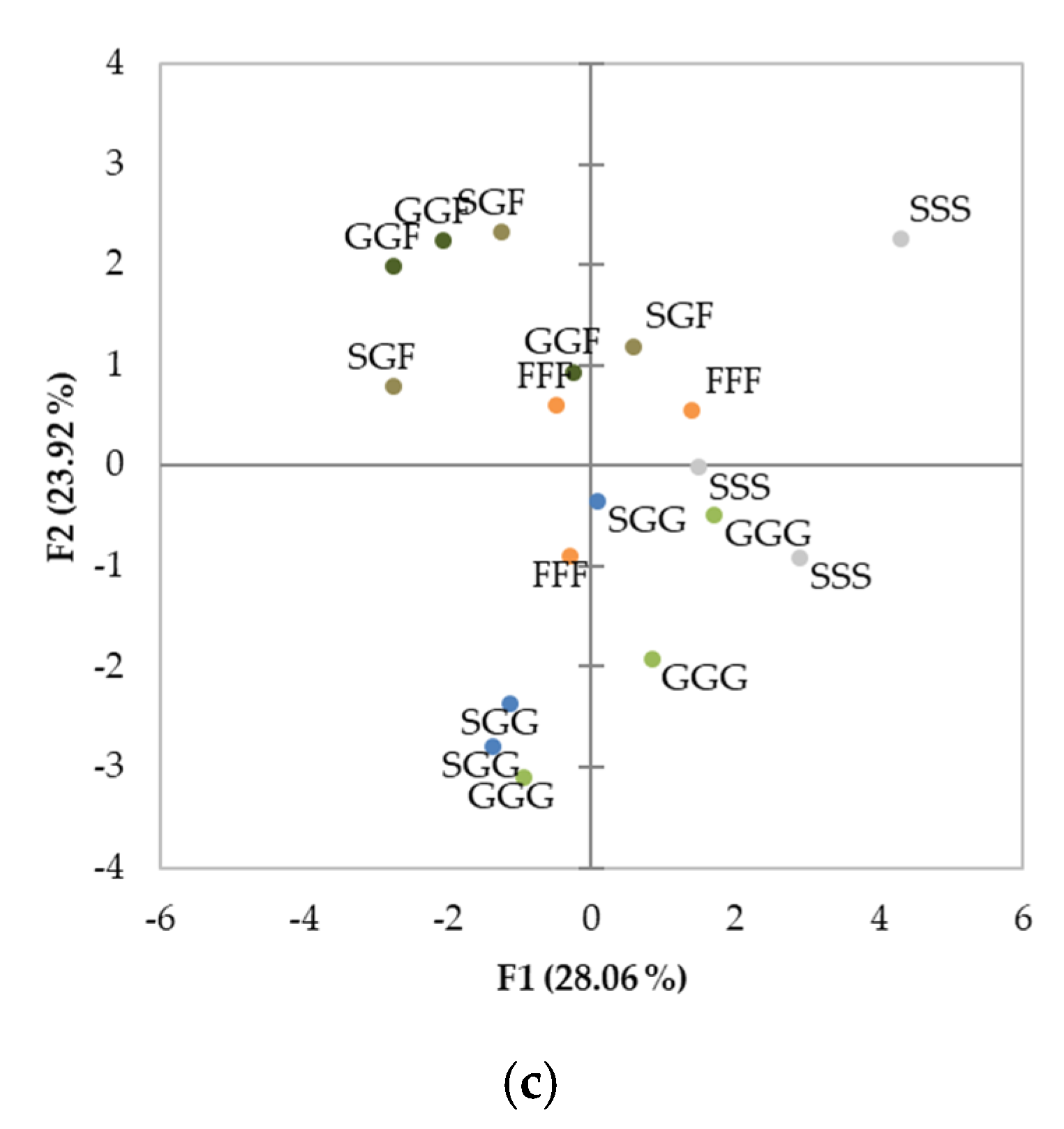
3.4. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, P.A.; Burns, C. Photobiology in protected horticulture. Food Energy Secur. 2016, 5, 223–238. [Google Scholar] [CrossRef]
- Cocetta, G.; Casciani, D.; Bulgari, R.; Musante, F.; Kołton, A.; Rossi, M.; Ferrante, A. Light use efficiency for vegetables production in protected and indoor environments. Eur. Phys. J. Plus 2017, 132, 132. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Vaštakaitė, V.; Gupta, S.D. Light-Emitting Diodes (LEDs) for Improved Nutritional Quality. In Light Emitting Diodes for Agriculture; Springer Science and Business Media LLC: Berlin, Germany, 2017; pp. 149–190. [Google Scholar]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Gautier, H.; Měch, R.; Prusinkiewicz, P.; Varlet-Grancher, C. 3D Architectural Modelling of Aerial Photomorphogenesis in White Clover (Trifolium repens L.) using L-systems. Ann. Bot. 2000, 85, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.D.; Barman, M.; Mitra, A. Photosynthetic apparatus plays a central role in photosensitive physiological acclimations affecting spinach (Spinacia oleracea L.) growth in response to blue and red photon flux ratios. Environ. Exp. Bot. 2018, 156, 170–182. [Google Scholar] [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of Seven Diverse Species to Blue and Green Light: Interactions with Photon Flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef]
- Samuolienė, G.; Viršilė, A.; Haimi, P.; Miliauskienė, J. Photoresponse to different lighting strategies during red leaf lettuce growth. J. Photochem. Photobiol. B Biol. 2020, 202, 111726. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Viršilė, A.; Samuolienė, G.; Vaštakaitė-Kairienė, V.; Jankauskienė, J.; Miliauskienė, J.; Novičkovas, A.; Duchovskis, P. Response of Mustard Microgreens to Different Wavelengths and Durations of UV-A LEDs. Front. Plant. Sci. 2019, 10, 1153. [Google Scholar] [CrossRef] [Green Version]
- Huché-Thélier, L.; Crespel, L.; Le Gourrierec, J.; Morel, P.; Sakr, S.; Leduc, N. Light signaling and plant responses to blue and UV radiations—Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Verdaguer, D.; Jansen, M.A.; Llorens, L.; Morales, L.O.; Neugart, S. UV-A radiation effects on higher plants: Exploring the known unknown. Plant Sci. 2017, 255, 72–81. [Google Scholar] [CrossRef]
- Štroch, M.; Materová, Z.; Vrábl, D.; Karlický, V.; Šigut, L.; Nezval, J.; Špunda, V. Protective effect of UV-A radiation during acclimation of the photosynthetic apparatus to UV-B treatment. Plant Physiol. Biochem. 2015, 96, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Allorent, G.; Petroutsos, D. Photoreceptor-dependent regulation of photoprotection. Curr. Opin. Plant Biol. 2017, 37, 102–108. [Google Scholar] [CrossRef]
- Yoshida, H.; Mizuta, D.; Fukuda, N.; Hikosaka, S.; Goto, E. Effects of varying light quality from single-peak blue and red light-emitting diodes during nursery period on flowering, photosynthesis, growth, and fruit yield of everbearing strawberry. Plant Biotechnol. 2016, 33, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Xu, H.; Shao, L.; Li, T.; Wang, Y.; Wang, R. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Env. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Schwarz, D.; Thompson, A.J.; Klãring, H.-P. Guidelines to use tomato in experiments with a controlled environment. Front. Plant Sci. 2014, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Higashi, T.; Tanigaki, Y.; Takayama, K.; Nagano, A.J.; Honjo, M.N.; Fukuda, H. Detection of Diurnal Variation of Tomato Transcriptome through the Molecular Timetable Method in a Sunlight-Type Plant Factory. Front. Plant Sci. 2016, 7, 87. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.; An, S.; Lee, B.; Chun, C. Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulture 2020, 6, 109. [Google Scholar] [CrossRef]
- Kreuger, M.; Meeuws, L.; Meeuws, G. Applications for Breeding and High-Wire Tomato Production in Plant Factory. In Smart Plant Factory; Kozai, T., Ed.; Springer: Singapore, 2018; pp. 289–299. ISBN 9789811310645. [Google Scholar]
- Izzo, L.G.; Mele, B.H.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Hernández, R.; Eguchi, T.; Deveci, M.; Kubota, C. Tomato seedling physiological responses under different percentages of blue and red photon flux ratios using LEDs and cool white fluorescent lamps. Sci. Hortic. 2016, 213, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, E.; Weerheim, K.; Schipper, R.; Dieleman, J.A. Partial replacement of red and blue by green light increases biomass and yield in tomato. Sci. Hortic. 2019, 249, 271–279. [Google Scholar] [CrossRef]
- Zhang, Y.; Kaiser, E.; Zhang, Y.; Zou, J.; Bian, Z.; Yang, Q.; Li, T. UVA radiation promotes tomato growth through morphological adaptation leading to increased light interception. Environ. Exp. Bot. 2020, 176, 104073. [Google Scholar] [CrossRef]
- Spalholz, H.; Perkins-Veazie, P.; Hernández, R. Impact of sun-simulated white light and varied blue:red spectrums on the growth, morphology, development, and phytochemical content of green- and red-leaf lettuce at different growth stages. Sci. Hortic. 2020, 264, 109195. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Duchovskis, P.; Urbonavičiūtė, A.; Samuolienė, G.; Jankauskienė, J.; Sakalauskaitė, J.; Šabajevienė, G.; Sirtautas, R.; Novičkovas, A. The effect of light-emitting diodes lighting on the growth of tomato transplants. Zemdir byste Agric. 2010, 97, 89–98. [Google Scholar]
- Şenilă, M.; Şenilă, L.; Roman, C. Evaluation of performance parameters for trace elements analysis in perennial plants using ICP-OES technique. J Plant Develop. 2011, 18, 87–93. [Google Scholar]
- Viršilė, A.; Miliauskienė, J.; Haimi, P.J.; Laužikė, K.; Samuolienė, G. The Comparison of Constant and Dynamic Red and Blue Light Irradiation Effects on Red and Green Leaf Lettuce. Agronomy 2020, 10, 1802. [Google Scholar] [CrossRef]
- LaNoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of Light Quality and Intensity on Diurnal Patterns and Rates of Photo-Assimilate Translocation and Transpiration in Tomato Leaves. Front. Plant Sci. 2018, 9, 756. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding Blue to Red Supplemental Light Increases Biomass and Yield of Greenhouse-Grown Tomatoes, but Only to an Optimum. Front. Plant Sci. 2019, 9, 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ieperen, W.; Savvides, A.; Fanourakis, D. Red and blue light effects during growth on hydraulic and stomatal conductance in leaves of young cucumber plants. Acta Hortic. 2012, 956, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Leperen, W. Plasticity of photosynthesis after the ‘red light syndrome’ in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Kalaitzoglou, P.; Van Ieperen, W.; Harbinson, J.; Van Der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.C.S.; Marcelis, L.F.M. Effects of Continuous or End-of-Day Far-Red Light on Tomato Plant Growth, Morphology, Light Absorption, and Fruit Production. Front. Plant Sci. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amoozgar, A.; Mohammadi, A.; Sabzalian, M.R. Impact of light-emitting diode irradiation on photosynthesis, phytochemical composition and mineral element content of lettuce cv. Grizzly. Photosynthetica 2017, 55, 85–95. [Google Scholar] [CrossRef]
- Pinho, P.; Jokinen, K.; Halonen, L. The influence of the LED light spectrum on the growth and nutrient uptake of hydroponically grown lettuce. Light. Res. Technol. 2016, 49, 866–881. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Viršilė, A.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Duchovskis, P. Nutrient Levels in Brassicaceae Microgreens Increase Under Tailored Light-Emitting Diode Spectra. Front. Plant Sci. 2019, 10, 1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lighting Spectra | Blue 447 nm µmol·m−2s−1 | Red 660 nm µmol·m−2s−1 | Far Red 740 nm µmol·m−2s−1 | Green 523 nm µmol·m−2s−1 | UV-A 385 nm mW·cm−2 |
|---|---|---|---|---|---|
| Seedling (S) | 75 | 165 | 10 | - | - |
| Growth (G) | 50 | 165 | 10 | 25 | - |
| Fruiting (F) | 60 | 180 | 10 | - | 1.6 |
| Treatment | Pr, µmol CO2 m−2·s−1 | Tr, mmol H2O m−2·s−1 | gs, mol H2O m−2·s−1 | WUE, µmol CO2 mmol−1 H2O | LUE, mol CO2 mol−1 Photons | Fv/Fm | ФPSII | PRI |
|---|---|---|---|---|---|---|---|---|
| Seedling: BBCH from 000 to 205 | ||||||||
| S | 23.6 a | 4.50 a | 0.82 a | 31.4 b | 0.019 a | 0.81 a | 0.75 b | 0.08 ab |
| G | 21.0 ab | 3.56 b | 0.51 b | 43.1 a | 0.017 ab | 0.82 a | 0.76 b | 0.09 a |
| F | 20.6 b | 3.29 b | 0.56 b | 41.4 a | 0.017 b | 0.73 b | 0.79 a | 0.07 b |
| Growth: BBCH from 205 to 619–701 | ||||||||
| SS | 16.6 a | 2.57 a | 0.11 a | 155.8 a | 0.07 a | 0.83 a | 0.77 b | 0.08 b |
| SG | 12.6 c | 1.44 bc | 0.06 bc | 207.0 a | 0.05 c | 0.83 a | 0.78 ab | 0.08 b |
| GG | 5.8 d | 1.13 c | 0.05 c | 155.2 a | 0.02 d | 0.81 b | 0.79 a | 0.08 b |
| FF | 14.6 b | 1.78 b | 0.08 b | 196.3 a | 0.06 b | 0.83 a | 0.78 ab | 0.09 a |
| Fruiting: BBCH from 619–701 to 801 | ||||||||
| SSS | 12.6 ab | 1.81 a | 0.05 a | 233.4 c | 0.05 ab | 0.81 ab | 0.08 a | 0.08 a |
| SGG | 10.6 b | 1.08 b | 0.04 ab | 297.6 bc | 0.04 b | 0.81 ab | 0.08 a | 0.08 a |
| GGG | 10.7 b | 1.27 ab | 0.04 ab | 273.0 bc | 0.04 b | 0.81 ab | 0.08 a | 0.08 a |
| SGF | 15.6 a | 1.14 b | 0.04 ab | 427.4 ab | 0.06 a | 0.79 b | 0.07 a | 0.07 a |
| FFF | 15.4 a | 1.39 ab | 0.05 a | 323.9 bc | 0.06 a | 0.82 a | 0.07 a | 0.07 a |
| GGF | 15.2 a | 0.92 b | 0.03 b | 565.3 a | 0.06 a | 0.81 ab | 0.07 a | 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samuolienė, G.; Miliauskienė, J.; Kazlauskas, A.; Viršilė, A. Growth Stage Specific Lighting Spectra Affect Photosynthetic Performance, Growth and Mineral Element Contents in Tomato. Agronomy 2021, 11, 901. https://doi.org/10.3390/agronomy11050901
Samuolienė G, Miliauskienė J, Kazlauskas A, Viršilė A. Growth Stage Specific Lighting Spectra Affect Photosynthetic Performance, Growth and Mineral Element Contents in Tomato. Agronomy. 2021; 11(5):901. https://doi.org/10.3390/agronomy11050901
Chicago/Turabian StyleSamuolienė, Giedrė, Jurga Miliauskienė, Algirdas Kazlauskas, and Akvilė Viršilė. 2021. "Growth Stage Specific Lighting Spectra Affect Photosynthetic Performance, Growth and Mineral Element Contents in Tomato" Agronomy 11, no. 5: 901. https://doi.org/10.3390/agronomy11050901
APA StyleSamuolienė, G., Miliauskienė, J., Kazlauskas, A., & Viršilė, A. (2021). Growth Stage Specific Lighting Spectra Affect Photosynthetic Performance, Growth and Mineral Element Contents in Tomato. Agronomy, 11(5), 901. https://doi.org/10.3390/agronomy11050901