
Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus




Abstract
:1. Introduction
- (1)
- Is it possible to fix a mathematical model for the biology of RPW in conditions of cold winter climates?
- (2)
- Once the above model has been fixed, is it possible to use an optimal control model to simulate the optimum time for application of chemical larval control?
2. Materials and Methods
2.1. Biological Data Used
2.2. Mathematical Model
2.3. Dynamic Model of Optimal Chemical Control
2.4. Software Used
3. Results
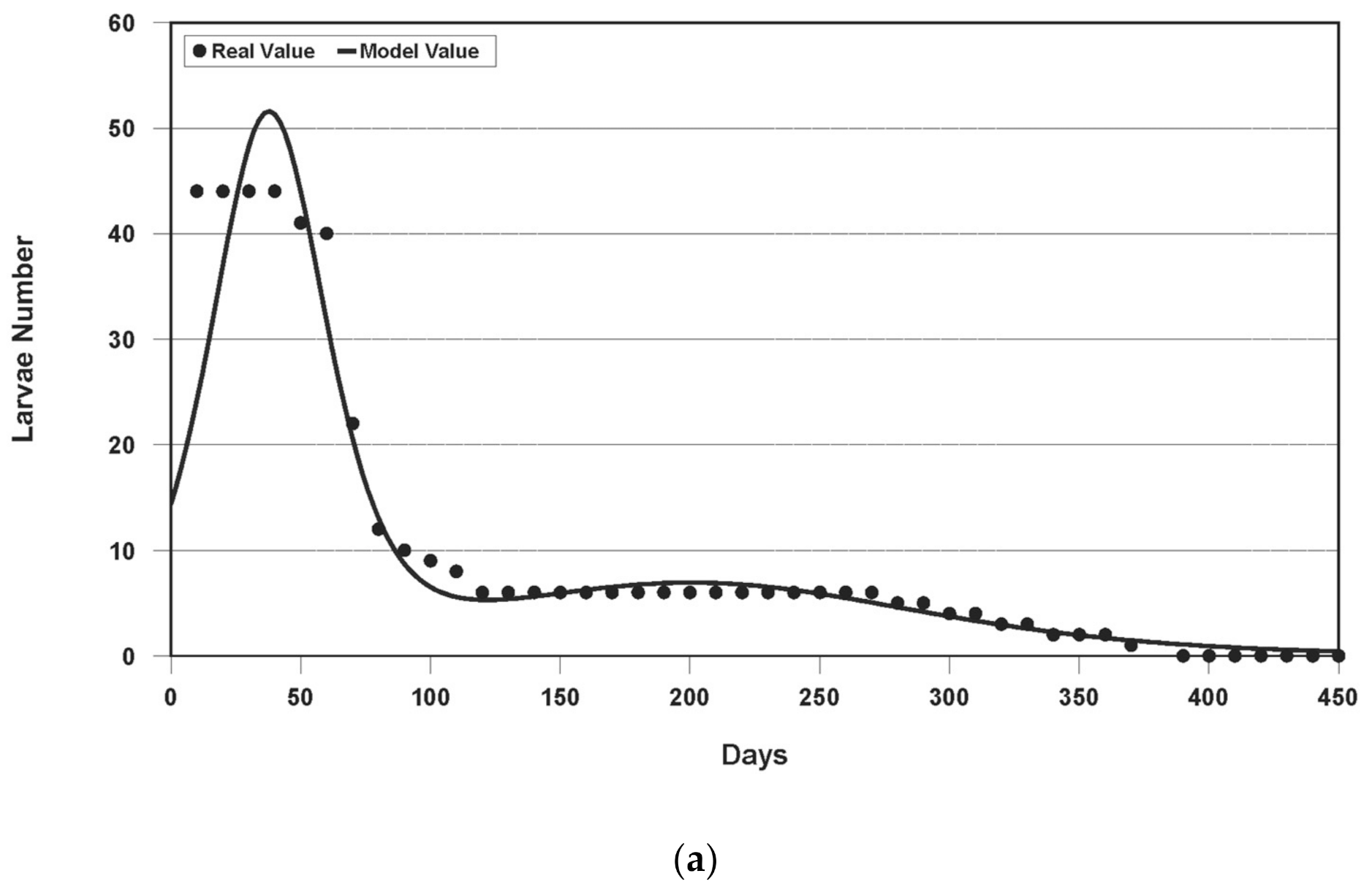
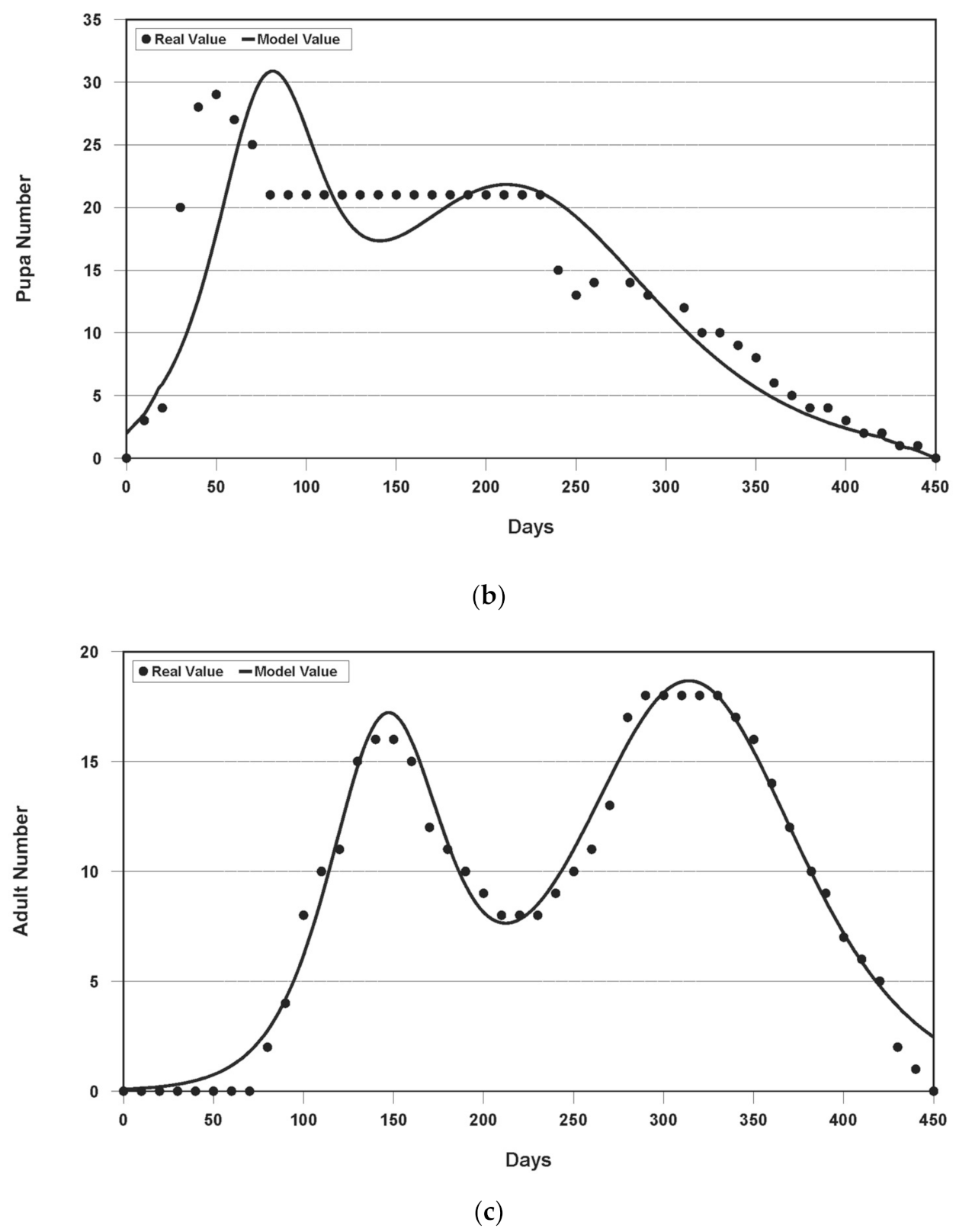
3.1. Results of Model Fitting
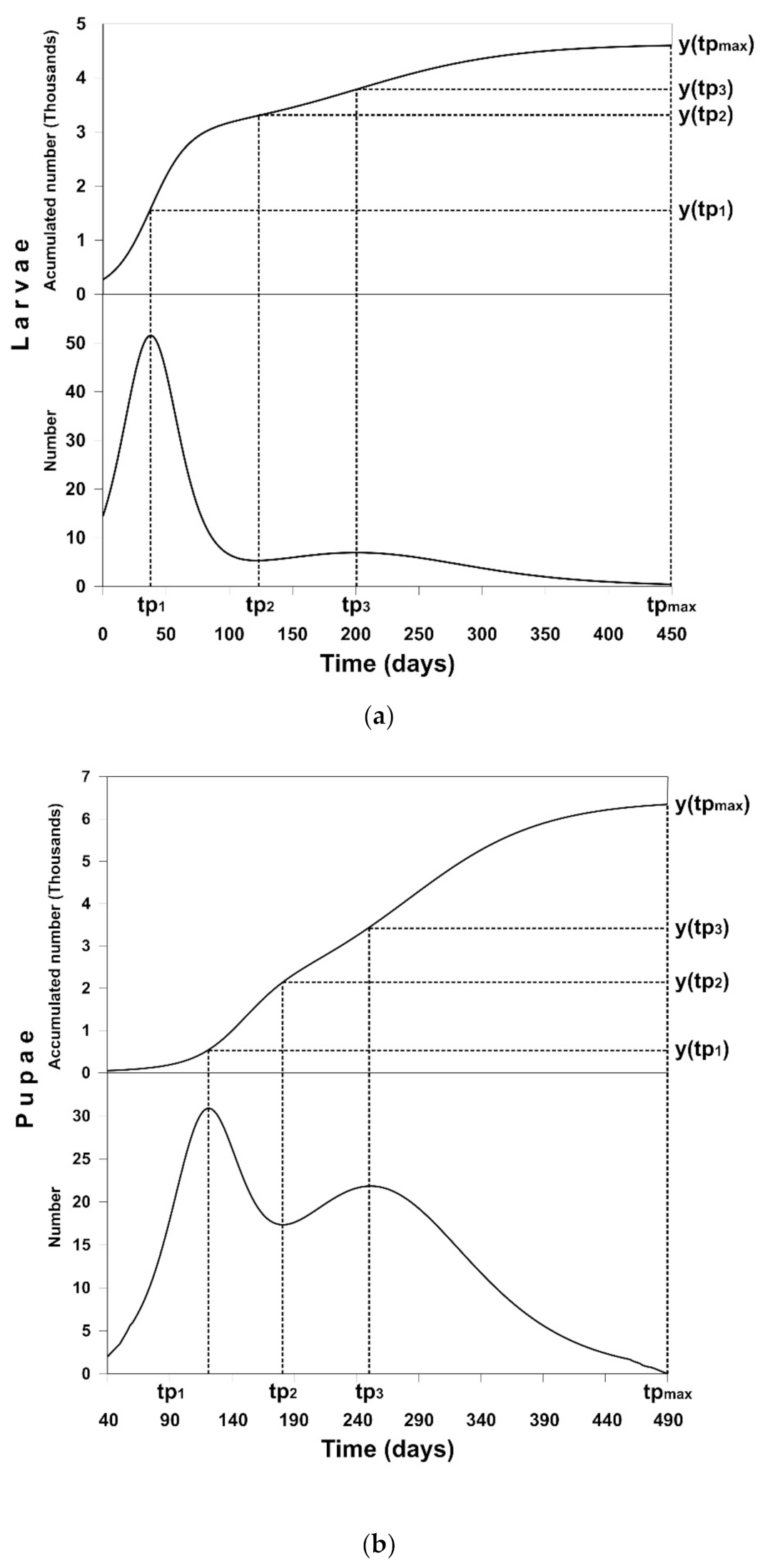
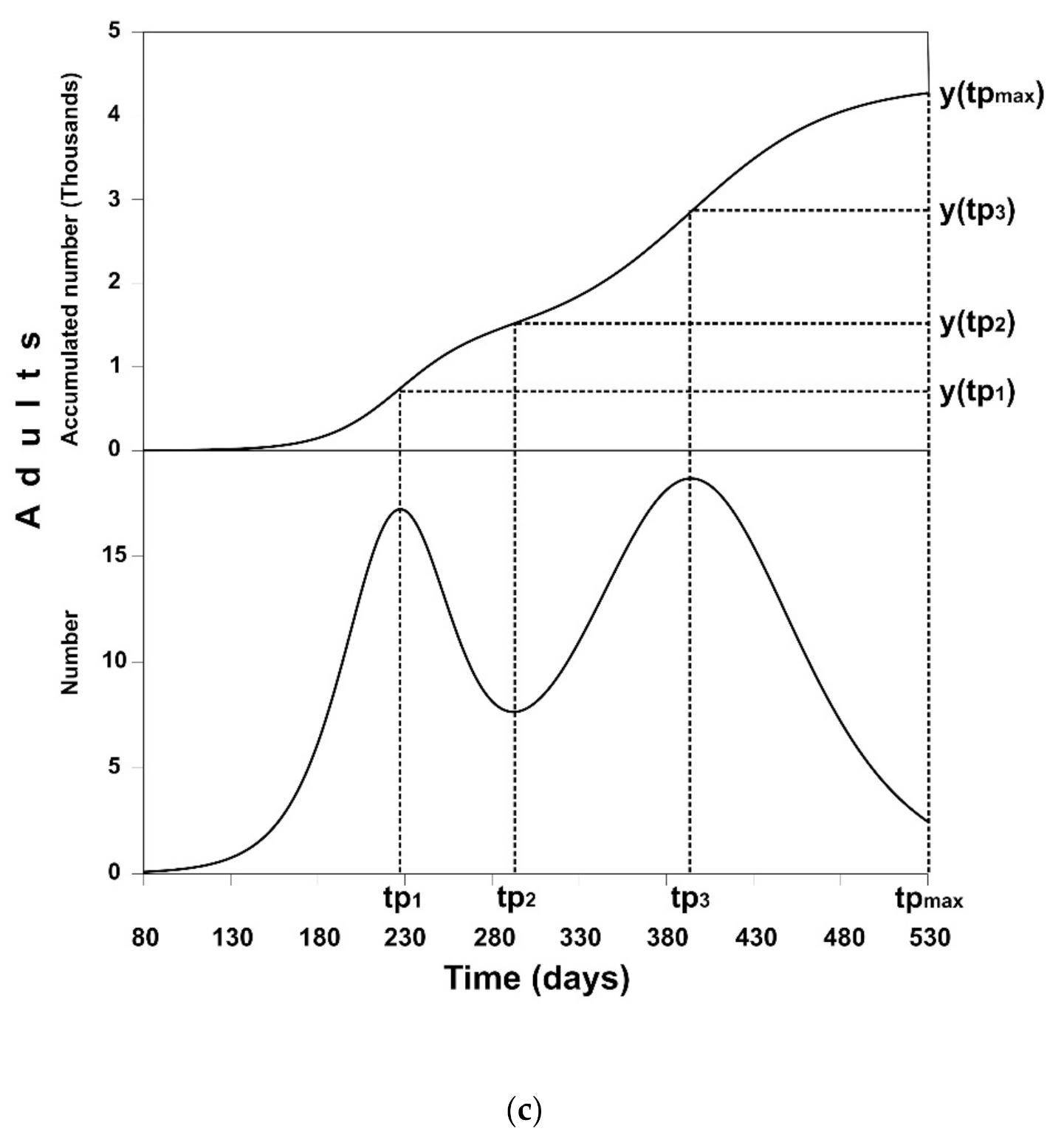
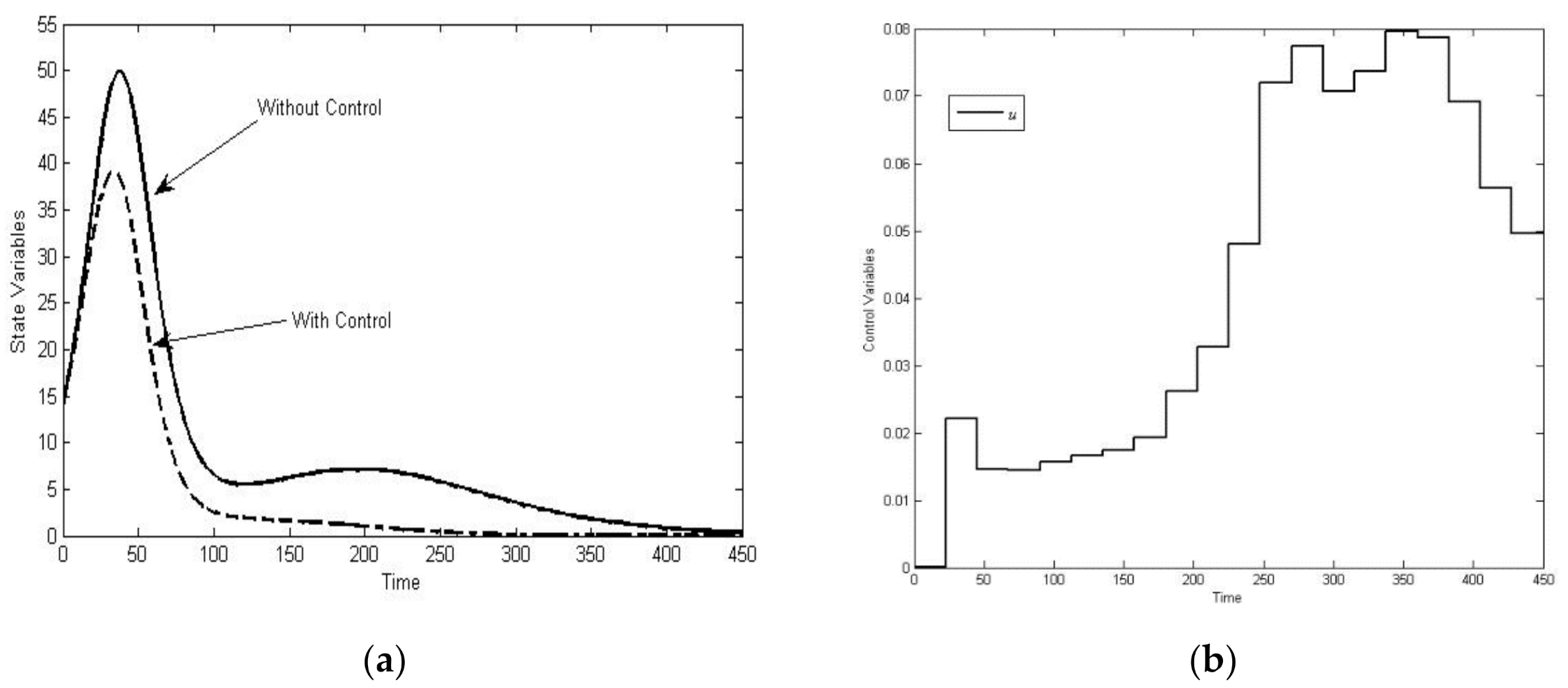
3.2. Solution of the Optimal Control Problem
4. Discussion
- (A)
- The model is sufficiently simple and flexible to allow its extension to this pest species in other geographical areas.
- (B)
- It saves some of the costs of experimentation on the effectiveness of the chemical control of RPW, as simulations can be carried out before expensive field trials, providing valuable information in advance.
- (C)
- With the introduction of optimisation into the model, the model could be used to calculate the ideal timing of chemical pest control, depending on climatic conditions and/or stage of infestation, with potential savings in treatment costs.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, M.M.; Cabello, T. Biología y Ecología del Curculiónido rojo de la Palmera, Rhynchophorus ferrugineus (Col.: Dryophthoridae); Universidad de Almeria: Almeria, Spain, 2005; pp. 1–202. [Google Scholar]
- Cabello, T. Rhynchophorus ferrugineus: Biología, dispersión y modo de acción. Phytoma 2012, 235, 36–38. [Google Scholar]
- CABI. Invasive species compendium: Rhynchophorus ferrugineus. Available online: https://www.cabi.org/isc/datasheet/47472 (accessed on 25 April 2021).
- Ferry, M.; Gómez, S. El picudo rojo de la palmera datilera: Gravedad de la plaga en España y necesidad de un cambio radical y urgente de estrategia en la lucha. Phytoma 2007, 184, 1–4. [Google Scholar]
- Ferry, M.; Gomez, S. El picudo rojo de la palmera datilera. Phytoma España 2007, 185, 42–46. [Google Scholar]
- Rochat, D.; Dembilio, O.; Jaques, J.A.; Suma, P.; Pergola, A.; Hamidi, R.; Kontodimas, D.; Soroker, V. Rhynchophorus ferrugineus: Taxonomy, distribution, biology, and life cicle. In Handbook of Major Palm Pests: Biology and Management; Soroker, V., Colazza, S., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 69–104. [Google Scholar]
- Peri, E.; Rochat, D.; Belusic, G.; Ilic, M.; Soroker, V.; Barkan, S.; Guarino, S.; Bue, P.L.; Colazza, S. Rhynchophorus ferrugineus: Behavior, ecology and communication. In Handbook of Major Palm Pests: Biology and Management; Soroker, V., Colazza, S., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 105–130. [Google Scholar]
- Milosavljevic, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewis, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive plam weevils, Rhynchophorus spp. J. Pest Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Giblin-Davis, R.M.; RomenoFaleiro, J.; Jacas, J.A.; Peña, J.E.; Vidyasagar, P.S.P.V. Biology and management of the red palm weevil, Rhynchophorus ferrugineus. In Potential Invasive Pests of Agricultural Crops; Peña, J.E., Ed.; CABI International: Walling-Ford, UK, 2013; pp. 1–34. [Google Scholar]
- Leon-Quinto, T.; Fimia, A.; Madrigal, R.; Serna, A. Morphological response of the red palm weevil, Rhynchophorus ferrugineus, to a transient low temperature analyzed by computer tomography and holographic microscopy. J. Therm. Biol. 2020, 94. [Google Scholar] [CrossRef]
- FCEC (Food Chain Evaluation Consortium). Quantification of Costs and Benefits of Amendments to the EU Plant Health Regime: Final Report. Available online: https://ec.europa.eu/food/sites/food/files/plant/docs/ph_biosec_rules_fcec_final_report_economic_study_plant_health_en.pdf (accessed on 9 November 2020).
- MacLeod, A.; Hussein, M. Economic and social impact of Rhynchophorus ferrugineus and Paysandisia archon on palms. In Handbook of Major Palm Pests: Biology and Management; Soroker, V., Colazza, S., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 54–68. [Google Scholar]
- Cobos-Suárez, J.M. Situation of R. ferrugineus in Spain. In Proceedings of the Red Palm Weevil Control Strategy for Europe: International Conference, Valencia, Spain, 5–6 May 2010. [Google Scholar]
- Jacas, J.A.; Dembilio, O.; Llácer, E. Research activities focused on management of red palm weevil at the UJI-IVIA Associated Unit (Region of Valencia, Spain). EPPO Bull. 2011, 41, 122–127. [Google Scholar] [CrossRef]
- Rao, P.V.S.; Subramaniam, T.R.; Abraham, E.V. Control of the red palm weevil on coconut. J. Plant. Crop. 1973, 1, 26–27. [Google Scholar]
- El-Ezaby, F.A. Injection as a method to control the Red Indian date palm weevil Rhynchophorus ferrugineus. Arab J. Plant Prot. 1997, 15, 31–38. [Google Scholar]
- Cabello, T.; De La Peña, J.; Barranco, P. Laboratory test of imidacloprid and oxamyl against Rhynchophorus ferrugineus, new pest of palms in Spain. Ann. Appl. Biol. 1997, 130, 2–3. [Google Scholar]
- Barranco, P.; de la Pena, J.; Martin, M.M.; Cabello, T. Efficiency of chemical control of the new palm pest Rhynchophorus ferrugineus (Col.: Curculionidae). Bol. Sanid. Veg. Plagas 1998, 24, 301–306. [Google Scholar]
- Hernandez-Marante, D.; Folk, F.; Sanchez, A.; Fernandez-Escobar, R. Control del curculiónido ferruginoso de las palmeras (Rhynchophorus ferrugineus) mediante inyecciones al tronco y pulverización foliar. Bol. Sanid. Veg. Plagas 2003, 29, 563–573. [Google Scholar]
- Faleiro, J.R. A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Col.: Rhynchophoridae) in coconut and date palm during the las one hundred years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar]
- Rafikov, M.; Balthazar, J.M. Optimal pest control problem in population dynamics. Comput. Appl. Math. 2005, 24, 65–81. [Google Scholar]
- Meng, X.; Song, Z.; Chen, L. A new mathematical model for optimal control strategies of integrated pest management. J. Biol. Syst. 2007, 15, 219–234. [Google Scholar] [CrossRef]
- Dabbs, K. Optimal Control in Discrete Pest Control Models. University of Tennessee Honors Thesis Projects. 2010. Available online: https://trace.tennessee.edu/utk_chanhonoproj/1375 (accessed on 6 May 2021).
- Amorim, L.; Bergamin-Filho, A.; Hau, B. Analysis of progress curves of sugarcane smut on different cultivars using functions of double sigmoid pattern. Phytopathology 1993, 83, 933–936. [Google Scholar] [CrossRef]
- Hau, B.; Amorim, L.; Bergamin-Filho, A. Mathematical functions to describe disease progress curves of double sigmoid pattern. Phytopathology 1993, 83, 928–932. [Google Scholar] [CrossRef]
- Carreño, R. 1996 Modelos logísticos. Aplicaciones a la Agronomía. Facultad de Ciencias Experimentales. Ph.D. Dissertation, Universidad de Almeria, Almeria, Spain, 1997; p. 168. [Google Scholar]
- Cabello, T.; Carreño, R. Métodos logísticos aplicados a la fenología de Noctuidos Plagas en el Sur de España (Lep.: Noctuidae). Bol. Sanid. Veg. Plagas 2002, 28, 226–319. [Google Scholar]
- Rafikov, M.; Angelelli, T. Optimization of biological pest control of sugarcane borer. In Proceedings of the 18th IEEE International Conference on Control Applications Part of 2009 IEEE Multi-Conference on Systems and Control, Saint Petersburg, Russia, 8–10 July 2009. [Google Scholar]
- Gallego, J.R.; López, I.; Gámez, M.; Cabello, T.; Varga, Z.; Garay, J. Simulation model applied to biological pest control by entomophagous species in commercial tomato greenhouses. Hung. Agric. Eng. 2013, 25, 67–70. [Google Scholar]
- Rossini, L.; Severini, M.; Contarini, M.; Speranza, S. A proposal of application in urban context to forecast pest insects’ life cycle. In Environmental and Territorial Modelling for Planning and Design; Leone, A., Gargiulo, C., Eds.; Federico II Open Access University Press: Napoli, Italy, 2018; pp. 169–178. [Google Scholar] [CrossRef]
- Hansen, E.M.; Bentz, B.J.; Powell, J.A.; Gray, D.R.; Vandygriff, J.C. Prepupal diapause and instar IV developmental rates of the spruce beetle, Dendroctonus rufipennis (Col.: Curculionidae). J. Insect Physiol. 2011, 57, 1347–1357. [Google Scholar] [CrossRef]
- Menu, F. Diapause development in the chestnut weevil Curculio elephas. Entomol. Exp. Appl. 1993, 69, 91–96. [Google Scholar] [CrossRef]
- Menu, F.; Debouzie, D. Larval development variation and adult emergence in the chestnut weevil Curculio elephans (Col., Curculionidae). J. Appl. Entomol. 1995, 119, 279–284. [Google Scholar] [CrossRef]
- Higaki, M. Effect of temperature on the termination of prolonged larval diapause in the chestnut weevil Curculio sikkimensis (Col.: Curculionidae). J. Insect Physiol. 2005, 51, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Prasifka, J.R.; Rinehart, J.P.; Yocum, G.D. Nonconstant thermal regimes enhance overwintering success and accelerate diapause development for Smicronyx fulvus (Col.: Curculionidae). J. Econ. Entomol. 2015, 108, 1804–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; Biological Survey of Canada: Monograph Series No. 1; National Museum of Natural Science: Ottawa, ON, Canada, 1987; pp. 1–439. [Google Scholar]
- Gordh, G.; Headrick, D.H. A Dictionary of Entomology; CABI Publishing: Wallingford, UK, 2001; pp. 1–1032. [Google Scholar]
- Rahalkar, G.W.; Harwalkar, M.R.; Rananavare, H.D. Development of red palm weevil, Rhynchophorus ferrugineus, on sugarcane. Indian J. Entomol. 1972, 34, 213–215. [Google Scholar]
- Rahalkar, G.W.; Harwalkar, M.R.; Rannavaer, H.D.; Tamhankar, A.J.; Shanthram, K. Rhynchophorus ferrugineus. In Handbook of Insect Rearing; Singh, P., Moore, R.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 279–286. [Google Scholar]
- Jandel Scientific. Table Curve 2D-User´s Manual. Version 5.0; Jandel Scientific: San Rafael, CA, USA, 1994. [Google Scholar]
- Bardsley, W.G. SIMFIT: Simulation, Fitting, Statistics and Plotting. Reference Manual; University of Manchester: Manchester, UK, 2010. [Google Scholar]
- Banga, J.R.; Balsa-Canto, E.; Moles, C.G.; Alonso, A.A. Dynamic optimization of bioprocesses: Efficient and robust numerical strategies. J. Biotechnol. 2005, 117, 407–419. [Google Scholar] [CrossRef]
- Menu, F.; Debouzie, D. Coin-flipping plasticity and prolonged diapause in insects: Example of the chestnut weevil Curculio elephas (Col.: Curculionidae). Oecologia 1993, 93, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Ushatinskaya, R.S. A critical review of the superdiapause in insects. Ann. Zool. 1984, 21, 3–30. [Google Scholar]
- Cabello, T. Population biology and dynamics of the Red Palm Weeevil, Rhynchophorus ferrugineus (Col.: Dryophothoridae) in Spain. In I Jornada Internacional sobre el Picudo Rojo de las Palmeras; Fundación Agrolimed, Generalitat Valenciana: Valencia, Spain, 2006; pp. 19–34. [Google Scholar]
- Potamitis, I.; Ganchev, T.; Kontodimas, D. On automatic bioacoustic detection of pests: The cases of Rhynchophorus ferrugineus and Sitophilus oryzae. J. Econ. Entomol. 2009, 102, 1681–1690. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Ruiz, V.; Moltó, E.; Tapia, G.; Téllez, M.M. Development of a bioacoustic sensor for the early detection of red palm weevil (Rhynchophorus ferrugineus Oliver). Crop Prot. 2010, 29, 671–676. [Google Scholar] [CrossRef]
- Centro Nacional de Información Geográfica. Instituto Geográfico Nacional. Atlas Nacional de España. Available online: http://centrodedescargas.cnig.es/CentroDescargas/busquedaRedirigida.do?ruta=PUBLICACION_CNIG_DATOS_VARIOS/aneTematico/Espana_Temperatura-media-de-las-maximas-de-enero_1981–2010_mapa_14670_spa.pdf# (accessed on 11 November 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Fitting Parameters (*) | Statistical Parmeters | |||||||
|---|---|---|---|---|---|---|---|---|---|
| a0 | a1 | a2 | a3 | a4 | a5 | d.f. | R2 adj. | p | |
| Larvae | 673.739 | 0.04410 | 3.56691 | 2117.26 | 0.01967 | 80.0043 | 38 | 0.97654 | <0.01 |
| Pupa | 74980.4 | 0.05752 | 792.265 | 10500.9 | 0.02003 | 115.495 | 40 | 0.97744 | <0.01 |
| Adult | 28372.7 | 0.04231 | 485.713 | 218719 | 0.02546 | 2901.76 | 40 | 0.95999 | <0.01 |
| Stage | Term (i) | Fitting Parameters | Statistical Parameters | ||||
|---|---|---|---|---|---|---|---|
| Ki | x0i | ri | d.f. | R2 adj. | p | ||
| Larvae | 1 | 2951.43 | 214.53 | 0.06771 | 37 | 0.99764 | <0.01 |
| 2 | 1673.39 | 57.6115 | 0.01661 | ||||
| Pupa | 1 | 1810.51 | 4.8024 | 0.05484 | 40 | 0.99872 | <0.01 |
| 2 | 4621.58 | 47.1067 | 0.01889 | ||||
| Adult | 1 | 1342.24 | 1.2492 | 0.04789 | 43 | 0.99951 | <0.01 |
| 2 | 3031.58 | 1.32482 | 0.02461 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solano-Rojas, Y.; Gámez, M.; López, I.; Garay, J.; Varga, Z.; Cabello, T. Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus. Agronomy 2021, 11, 920. https://doi.org/10.3390/agronomy11050920
Solano-Rojas Y, Gámez M, López I, Garay J, Varga Z, Cabello T. Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus. Agronomy. 2021; 11(5):920. https://doi.org/10.3390/agronomy11050920
Chicago/Turabian StyleSolano-Rojas, Y., M. Gámez, I. López, J. Garay, Z. Varga, and T. Cabello. 2021. "Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus" Agronomy 11, no. 5: 920. https://doi.org/10.3390/agronomy11050920
APA StyleSolano-Rojas, Y., Gámez, M., López, I., Garay, J., Varga, Z., & Cabello, T. (2021). Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus. Agronomy, 11(5), 920. https://doi.org/10.3390/agronomy11050920