
Open Field Simulating Nocturnal Warming on Summer Maize Performance in the North China Plain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Sampling and Measurement
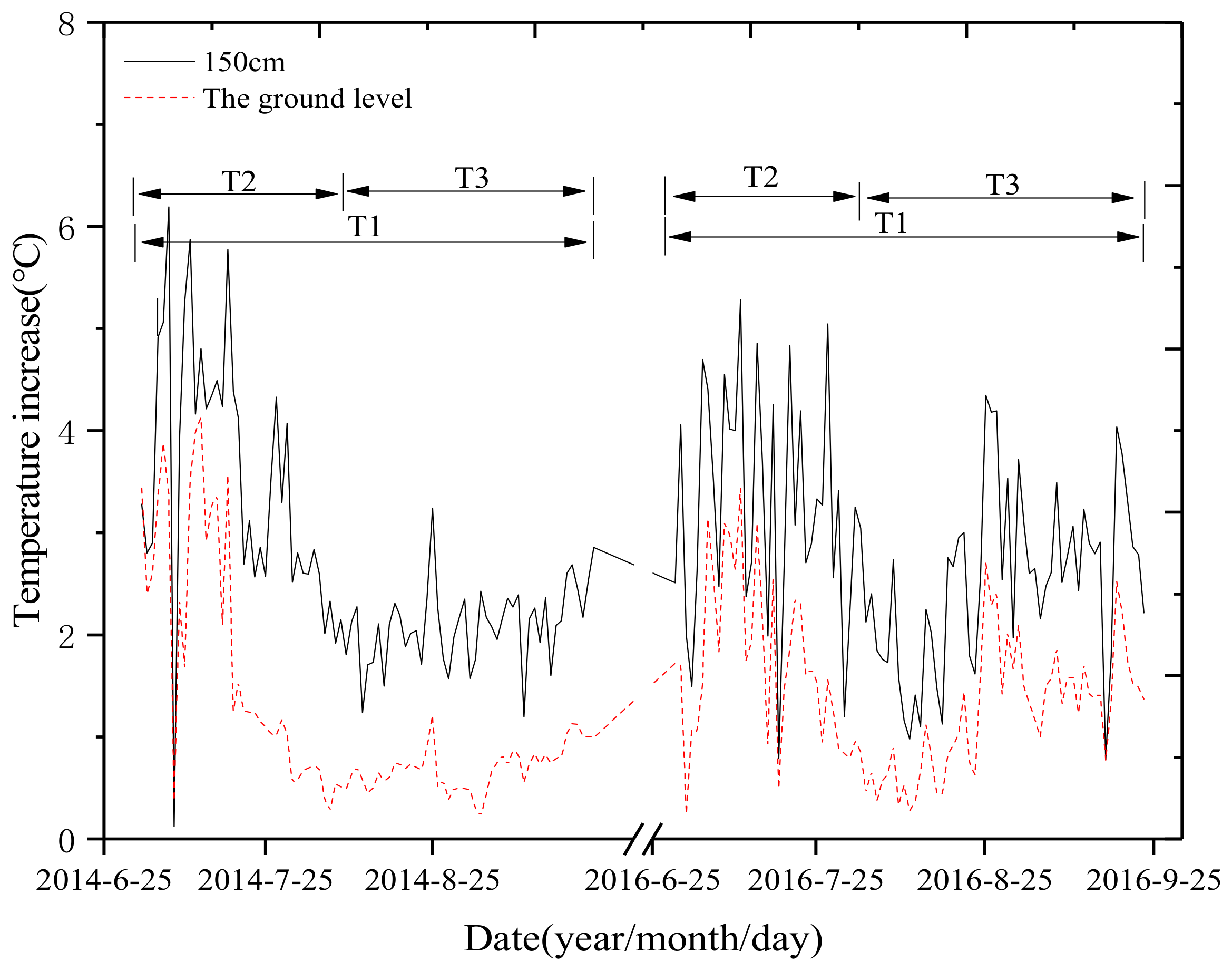
2.3.1. Temperature, Rainfall, and Soil Water Content
2.3.2. Crop Phenology, Biomass and Yield
2.3.3. Leaf Areas and Specific Ear Leaf Weight
2.3.4. Leaf Photosynthesis and Respiration Measurement
2.4. Statistical Analyses
3. Results
3.1. Weather Conditions and Temperature Changes under Nocturnal Warming
3.2. Nocturnal Warming Effects on the Soil Water Content
3.3. Crop Development under Nocturnal Warming
3.4. Leaf Area Index, Ear Leaf Area, Specific Leaf Weight, and Photosynthesis and Respiration Rates
3.5. Grain and Biomass Yield and Grain Yield Components
4. Discussion
4.1. Phenological Effects of NW
4.2. Photosynthesis and Respiration
4.3. Biomass Production and Harvest Index
4.4. Effects of NW during Different Stages on Maize Production
4.5. NW Effects under Different Soil Water Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014–Synthesis Report; IPCC: Geneva, Szwitzerland, 2014; p. 10. [Google Scholar]
- Easterling, D.R.; Horton, B.; Jones, P.D.; Peterson, T.C.; Karl, T.R.; Parker, D.E.; Salinger, M.J.; Razuvayev, V.; Plummer, N.; Jamason, P.; et al. Maximum and minimum temperature trends for the globe. Science 1997, 277, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, C.; Müller, C.; Wang, C.; Ciais, P.; Janssens, I.; Peñuelas, J.; Asseng, S.; Li, T.; Elliott, J.; et al. Emergent constraint on crop yield response to warmer temperature from field experiments. Nat. Sustain. 2020. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Lei, C.; Deng, A.; Qian, C.; Hoogmoed, W.; Zhang, W. Will higher minimum temperatures increase corn production in Northeast China? An analysis of historical data over 1965–2008. Agricb. For. Meteorol. 2011, 151, 1580–1588. [Google Scholar] [CrossRef]
- Meng, Q.; Hou, P.; Lobell, D.B.; Wang, H.; Cui, Z.; Zhang, F.; Chen, X. The benefits of recent warming for maize production in high latitude China. Clim. Chang. 2014, 122, 341–349. [Google Scholar] [CrossRef]
- Fan, Y.; Tian, M.; Jing, Q.; Tian, Z.; Han, H.; Jiang, D.; Cao, W.; Dai, T. Winter night warming improves pre-anthesis crop growth and post-anthesis photosynthesis involved in grain yield of winter wheat (Triticum aestivum L.). Field Crops Res. 2015, 178, 100–108. [Google Scholar] [CrossRef]
- Yang, H.; Lu, D.; Shen, X.; Cai, X.; Lu, W. Heat stress at different grain filling stages affects fresh waxy maize grain yield and quality. Cereal Chem. 2015, 92, 258–264. [Google Scholar] [CrossRef]
- Liu, S.; Mo, X.; Lin, Z.; Xu, Y.; Ji, J.; Wen, G.; Richey, J. Crop yield responses to climate change in the Huang-Huai-Hai plain of China. Agric. Water Manag. 2010, 97, 1195–1209. [Google Scholar] [CrossRef]
- Li, K.; Xiong, W.; Pan, J.; Lin, E.; Li, Y.; Han, X. Trend evaluation on changes of maize yield in china under global warming by 1.5 °C and 2.0 °C. Chin. J. Agrometeorol. 2018, 39, 765–777. [Google Scholar]
- Zheng, C.; Zhang, J.; Chen, J.; Chen, C.; Tian, Y.; Deng, A.; Song, Z.; Nawaz, M.M.; van Groenigen, K.J.; Zhang, W. Nighttime warming increases winter-sown wheat yield across major Chinese cropping regions. Field Crops Res. 2017, 214, 202–210. [Google Scholar] [CrossRef]
- Mohammed, A.R.; Tarpley, L. Effects of high night temperature and spikelet position on yield-related parameters of rice (Oryza sativa L.) plants. Eur. J. Agron. 2010, 33, 117–123. [Google Scholar] [CrossRef]
- Chen, J.; Chen, C.; Tian, Y.; Zhang, X.; Dong, W.; Zhang, B.; Zhang, J.; Zheng, C.; Deng, A.; Song, Z.; et al. Differences in the impacts of nighttime warming on crop growth of rice-based cropping systems under field conditions. Eur. J. Agron. 2017, 82, 80–92. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, R.; Wang, Y. Night-time warming affects N and P dynamics and productivity of winter wheat plants. Can. J. Plant. Sci. 2013, 93, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Cantarero, M.G.; Cirilo, A.G.; Andrade, F.H. Night temperature at silking affects kernel set in maize. Crop. Sci. 1999, 39, 703–710. [Google Scholar] [CrossRef]
- Chen, J.; Tian, Y.; Zhang, X.; Zheng, C.; Song, Z.; Deng, A.; Zhang, W. Nighttime warming will increase winter wheat yield through improving plant development and grain growth in north China. J. Plant. Growth Regul. 2014, 33, 397–407. [Google Scholar] [CrossRef]
- Chen, Y.; Han, X.; Si, W.; Wu, Z.; Chien, H.; Okamoto, K. An assessment of climate change impacts on maize yields in Hebei province of China. Sci. Total Environ. 2017, 581, 507–517. [Google Scholar] [CrossRef]
- Loka, D.A.; Oosterhuis, D.M. Effect of high night temperatures on cotton respiration, ATP levels and carbohydrate content. Environ. Exp. Bot. 2010, 68, 258–263. [Google Scholar] [CrossRef]
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Combined effects of elevated [CO2] and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (Oryza sativa L.). Agric. For. Meteorol. 2010, 150, 1174–1181. [Google Scholar] [CrossRef]
- Borrás, L.; Westgate, M.E.; Astini, J.P.; Echarte, L. Coupling time to silking with plant growth rate in maize. Field Crops Res. 2007, 102, 73–85. [Google Scholar] [CrossRef]
- Lizaso, J.I.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.A.; Lorite, I.J.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crops Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Dewar, R.C.; Medlyn, B.E.; McMurtrie, R.E. Acclimation of the respiration/photosynthesis ratio to temperature: Insights from a model. Glob. Chang. Biol 1999, 5, 615–622. [Google Scholar] [CrossRef]
- Loveys, B.R.; Atkinson, L.J.; Sherlock, D.J.; Roberts, R.L.; Fitter, A.H.; Atkin, O.K. Thermal acclimation of leaf and root respiration: An investigation comparing inherently fast- and slow-growing plant species. Glob. Chang. Biol. 2003, 9, 895–910. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, M.H.; Murthy, R.; Griffin, K.L. The relative impacts of daytime and night-time warming on photosynthetic capacity in Popul. deltoides. Plant. Cell Environ. 2002, 25, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Guo, L.; Hou, R.; Zhou, H.; Hao, L.; Li, F.; Cheng, D.; Peng, Z.; Xu, M. Experimental warming enhances the carbon gain but does not affect the yield of maize (Zea mays L.) in the North China Plain. Flora 2018, 240, 152–163. [Google Scholar] [CrossRef]
- Frantz, J.M.; Cometti, N.N.; Bugbee, B. Night temperature has a minimal effect on respiration and growth in rapidly growing plants. Ann. Bot. 2004, 94, 155–166. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Hobbs, R.J.; Mayfield, M.M. Specific leaf area responses to environmental gradients through space and time. Ecology 2014, 95, 399–410. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, G.; He, Q.; Zhou, L.; Ji, Y.; Zhou, M. Environmental explanation of maize specific leaf area under varying water stress regimes. Environ. Exp. Bot. 2020, 171, 103932. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Otegui, M.E.; Bonhomme, R. Grain yield components in maize—II. Postsilking growth and kernel weight. Field Crops Res. 1998, 56, 257–264. [Google Scholar] [CrossRef]
- Otegui, M.E.; Bonhomme, R. Grain yield components in maize—I. Ear growth and kernel set. Field Crops Res. 1998, 56, 247–256. [Google Scholar] [CrossRef]
- Cirilo, A.G.; Andrade, F.H. Sowing date and kernel weight in maize. Crop. Sci. 1996, 36, 325–331. [Google Scholar] [CrossRef]
- Zhou, B.; Yue, Y.; Sun, X.F.; Wang, X.B.; Wang, Z.M.; Ma, W.; Zhao, M. Maize grain yield and dry matter production responses to variations in weather conditions. Agron. J. 2016, 108, 196–204. [Google Scholar] [CrossRef]
- Sun, H.; Liu, C.M.; Wang, Z.H.; Zhang, X.Y.; Dong, B.F. Changing trend of precipitation and its effects on crop productivity in the piedment of Taihang mountain. Chin. J. Eco-Agric. 2007, 15, 18–21. [Google Scholar]
- Fang, Q.; Zhang, X.Y.; Chen, S.Y.; Shao, L.W.; Sun, H.Y.; Yan, Z.Z. Selecting traits to reduce seasonal yield variation of summer maize in the North China Plain. Agron. J. 2019, 111, 343–353. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extreme. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Shim, D.; Lee, K.-J.; Lee, B.-W. Response of phenology- and yield-related traits of maize to elevated temperature in a temperate region. Crop. J. 2017, 5, 305–316. [Google Scholar] [CrossRef]
- Siebers, M.H.; Slattery, R.A.; Yendrek, C.R.; Locke, A.M.; Drag, D.; Ainsworth, E.A.; Bernacchi, C.J.; Ort, D.R. Simulated heat waves during maize reproductive stages alter reproductive growth but have no lasting effect when applied during vegetative stages. Agric. Ecosyst. Environ. 2017, 240, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Vanaja, M.; Poldasari, S.; Kumar, G.; Razzaq, A.; Vagheera, P.; Narayana, J.; Yadav, S.; Sarkar, B.; Mandapaka, M. Elevated temperature and moisture deficit stress impact on phenology, physiology and yield responses of hybrid maize. J. Agrometeorol. 2017, 19, 295–300. [Google Scholar]
- Song, L.B.; Jin, J.M. Improving CERES-Maize for simulating maize growth and yield under water stress conditions. Eur. J. Agron. 2020, 117, 13. [Google Scholar] [CrossRef]
- Li, J.; Dong, W.; Oenema, O.; Chen, T.; Hu, C.; Yuan, H.; Zhao, L. Irrigation reduces the negative effect of global warming on winter wheat yield and greenhouse gas intensity. Sci. Total Environ. 2019, 646, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water-use efficiency in summer maize (Zea mays L.) throughout the growth cycle. Acta Physiol. Plant. 2011, 34, 1043–1053. [Google Scholar] [CrossRef]
- Sun, H.Y.; Zhang, X.Y.; Wang, E.L.; Chen, S.Y.; Shao, L.W.; Qin, W.L. Assessing the contribution of weather and management to the annual yield variation of summer maize using APSIM in the North China Plain. Field Crops Res. 2016, 194, 94–102. [Google Scholar] [CrossRef]
- Chen, X.; Jeong, S.-J. Irrigation enhances local warming with greater nocturnal warming effects than daytime cooling effects. Environ. Res. Lett. 2018, 13, 024005. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | Phenophase Date (day/month) | VT-R1 Interval (days) | Days before R1 (days) | Days after R1 (days) | Whole Growing Season (days) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sowing Date | V6 | VT | R1 | R6 | ||||||
| 2014 | CK | 13-June | 4-July | 7-August | 8-August | 26-September | 1 | 56 | 48 | 104 |
| T1 | 13-June | 4-July | 6-August | 9-August | 25-September | 3 | 57 | 46 | 103 | |
| T2 | 13-June | 4-July | 6-August | 9-August | 25-September | 3 | 57 | 46 | 103 | |
| T3 | 13-June | 4-July | 6-August | 8-August | 25-September | 2 | 56 | 47 | 103 | |
| 2016 | CK | 16-June | 7-July | 5-August | 7-August | 27-September | 2 | 51 | 50 | 101 |
| T1 | 16-June | 7-July | 5-August | 9-August | 26-September | 4 | 53 | 47 | 100 | |
| T3 | 16-June | 7-July | 5-August | 7-August | 26-September | 2 | 51 | 49 | 100 | |
| Year | Treatments | Aboveground Biomass at Blister Stage (kg ha−1) | Aboveground Biomass at Maturity (Mg ha−1) | Aboveground Biomass Accumulation between the Blister and Maturity (kg ha−1) | Yield (kg ha−1) | 1000-Kernel Weight (g) | Kernel Number per Cob | Harvest Index (HI) |
|---|---|---|---|---|---|---|---|---|
| 2014 | CK | 8341.4a | 15.50a | 7163.1a | 8213.0a | 268.8a | 482.1a | 0.56a |
| T1 | 7764.2a | 13.92b | 6158.4b | 7449.4b | 248.0b | 477.7a | 0.52b | |
| T2 | 8151.6a | 15.23a | 7077.5a | 8129.0a | 266.4a | 463.8a | 0.54a | |
| T3 | 8440.2a | 14.58ab | 6142.8b | 7932.7ab | 267.9a | 501.5a | 0.53ab | |
| 2016 | CK | 6713.2a | 14.16a | 7444.2a | 7784.0a | 331.7a | 446.8a | 0.54a |
| T1 | 6312.6a | 12.28b | 5964.4b | 6671.7b | 317.7b | 456.5a | 0.51b | |
| T3 | 6634.5a | 12.89ab | 6257.7b | 6773.1b | 313.0b | 458.9a | 0.51b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, J.; Feng, J.; Zhang, X.; Chen, S.; Shao, L. Open Field Simulating Nocturnal Warming on Summer Maize Performance in the North China Plain. Agronomy 2021, 11, 992. https://doi.org/10.3390/agronomy11050992
Niu J, Feng J, Zhang X, Chen S, Shao L. Open Field Simulating Nocturnal Warming on Summer Maize Performance in the North China Plain. Agronomy. 2021; 11(5):992. https://doi.org/10.3390/agronomy11050992
Chicago/Turabian StyleNiu, Junfang, Junxia Feng, Xiying Zhang, Suying Chen, and Liwei Shao. 2021. "Open Field Simulating Nocturnal Warming on Summer Maize Performance in the North China Plain" Agronomy 11, no. 5: 992. https://doi.org/10.3390/agronomy11050992
APA StyleNiu, J., Feng, J., Zhang, X., Chen, S., & Shao, L. (2021). Open Field Simulating Nocturnal Warming on Summer Maize Performance in the North China Plain. Agronomy, 11(5), 992. https://doi.org/10.3390/agronomy11050992