
The Development of the Genic SSR Markers for Analysis of Genetic Diversity in Gooseberry Cultivars

Abstract
:1. Introduction
2. Materials and Methods
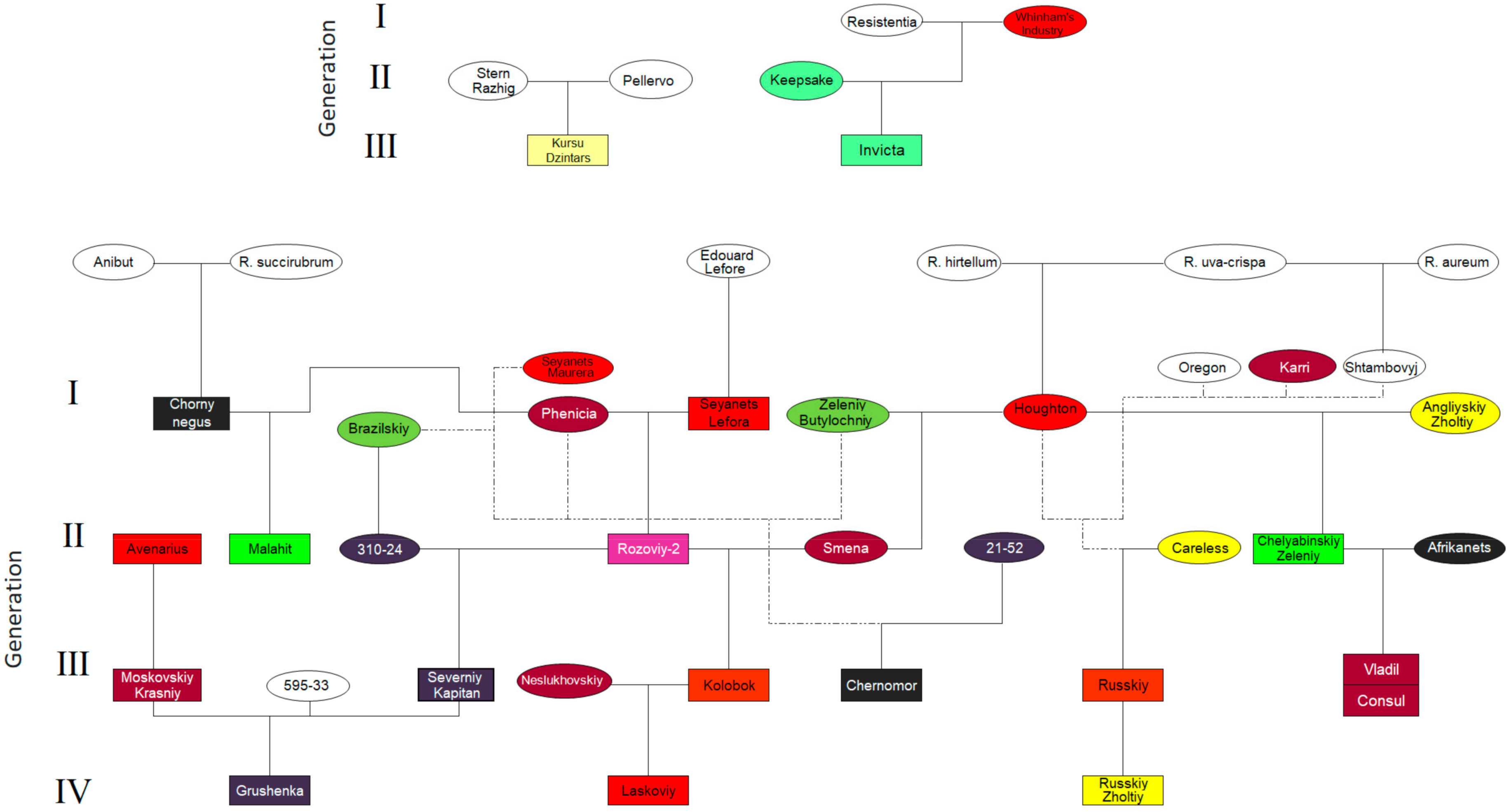
2.1. Plant Materials
2.2. Search for Microsatellite Loci and Primer Design for PCR
2.3. DNA Extraction, PCR Conditions, and Fragment Analysis
2.4. Genetic Data Analysis
3. Results
3.1. Polymorphism and Genetic Diversity Analysis
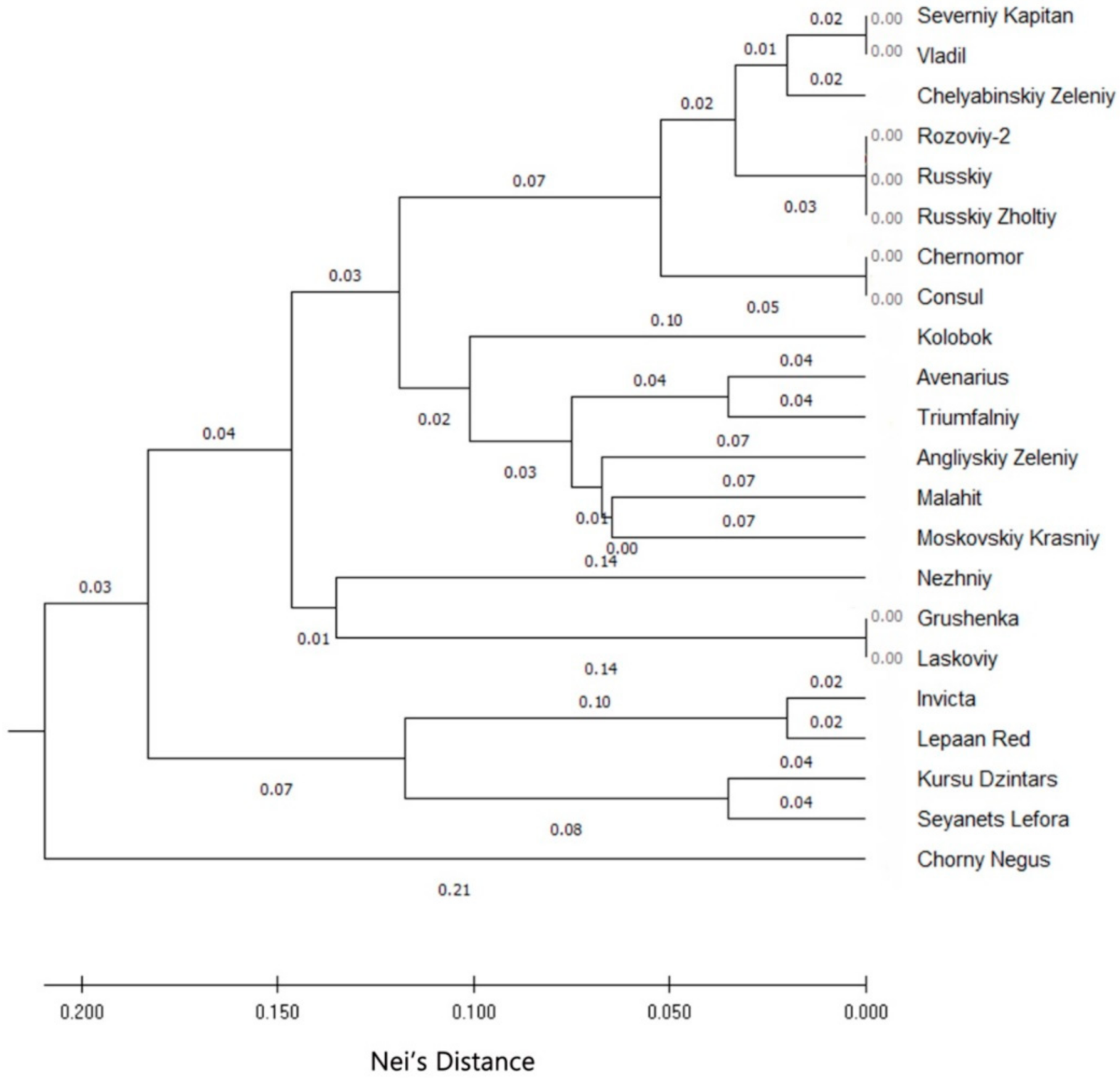
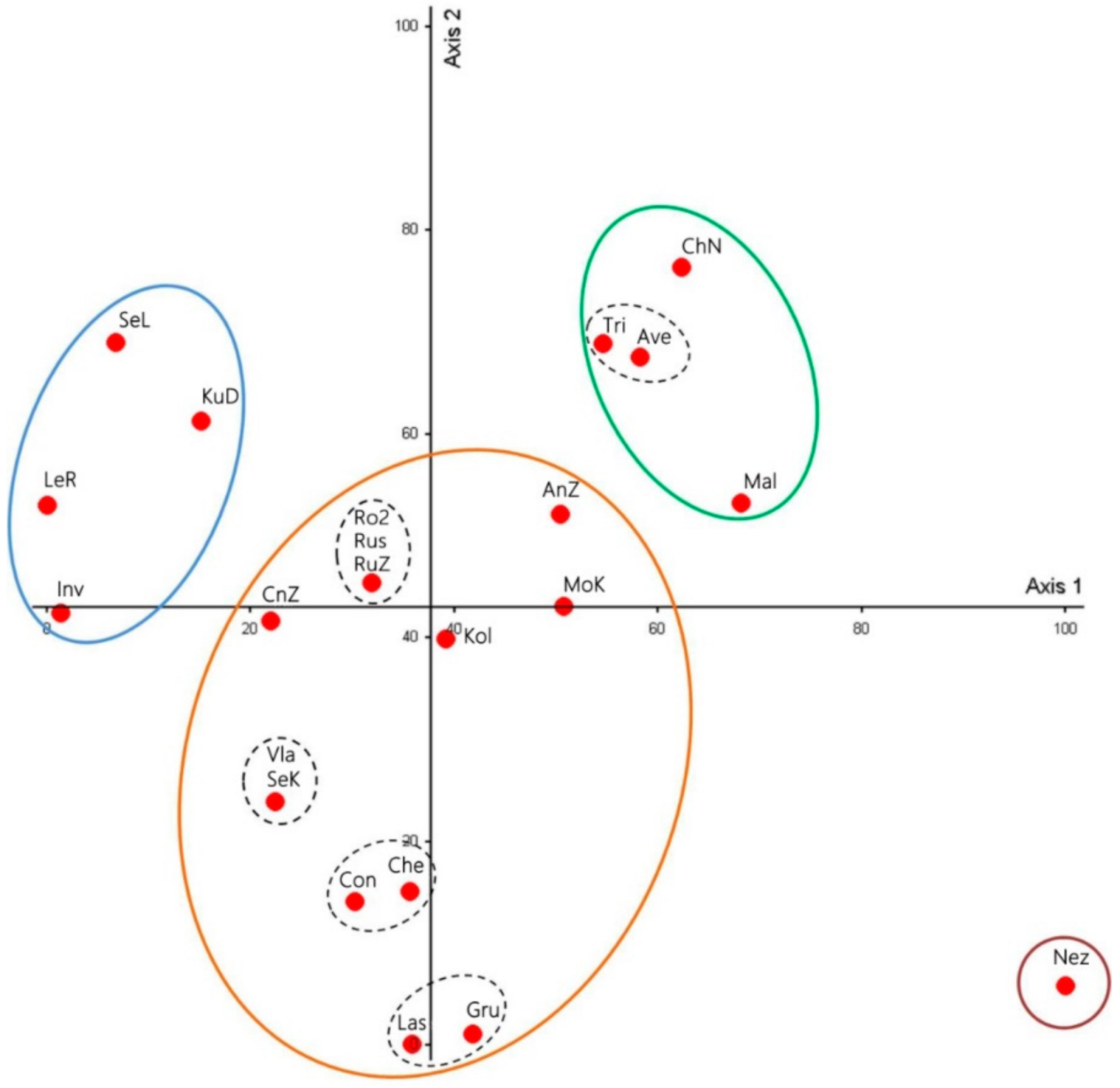
3.2. Cluster Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, M.K.; Nazir, N.; Parray, E.A.; Sundouri, A.S. Production potential of some cape gooseberry selections. J. Pharmacogn. Phytochem. 2019, 8, 1215–1216. [Google Scholar]
- Folmer, F.; Basavaraju, U.; Jaspars, M.; Hold, G.; El-Omar, E.; Dicato, M.; Diederich, M. Anticancer effects of bioactive berry compounds. Phytochem. Rev. 2014, 13, 295–322. [Google Scholar] [CrossRef]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluta, S. New challenges in the Ribes breeding and production. Acta Hortic. 2012, 946, 27–35. [Google Scholar] [CrossRef]
- Popova, I.V. Kryzhovnik; Agropromizdat: Moscow, Russia, 1985; p. 39. [Google Scholar]
- FAOSTAT. 2020. Available online: http://www.fao.org/faostat/ (accessed on 31 January 2021).
- Brennan, R.; Graham, J. Improving fruit quality in Rubus and Ribes through breeding. Funct. Plant Sci. Biotechnol. 2009, 3, 22–29. [Google Scholar]
- Masny, A.; Pluta, S.; Seliga, Ł. Breeding value of selected blackcurrant (Ribes nigrum L.) genotypes for early-age fruit yield and its quality. Euphytica 2018, 214, 89. [Google Scholar] [CrossRef]
- Abreu, I.N.; Brennan, R.M.; Kanichukattu, E.N.; Stewart, D.; Hancock, R.D.; McDougall, G.J.; Hackett, C.A. Quantitative trait loci mapping of polyphenol metabolites in blackcurrant (Ribes nigrum L.). Metabolomics 2020, 16, 25. [Google Scholar] [CrossRef]
- Brennan, R.; Jorgensen, L.; Hackett, C.; Woodhead, M.; Gordon, S.; Russell, J. The development of a genetic linkage map of blackcurrant (Ribes nigrum L.) and the identification of regions associated with key fruit quality and agronomic traits. Euphytica 2008, 161, 19–34. [Google Scholar] [CrossRef]
- Collard, B.C.; Mackill, C.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Phil. Trans. R. Soc. B. 2008, 363, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Cavanna, M.; Marinoni, D.T.; Beccaro, G.L.; Bounous, G. Microsatellite-based evaluation of Ribes spp. germplasm. Genome 2009, 52, 839–848. [Google Scholar] [CrossRef]
- Karhu, S.; Antonius, K.; Rantala, S.; Kaldmae, H.; Pluta, S.; Rumpunen, K.; Ryliskis, D.; Sasnauskas, A.; Schulte, E.; Strautina, S.; et al. A multinational approach for conserving the European genetic resources of currants and gooseberry. Acta Hortic. 2012, 926, 27–32. [Google Scholar] [CrossRef]
- Wu, X.; Gu, L.; Prior, R.L.; McKay, S. Characterization of anthocyanins and proanthocyanidins in some cultivars of Ribes, Aronia, and Sambucusand their antioxidant capacity. J. Agric. Food Chem. 2004, 52, 7846–7856. [Google Scholar] [CrossRef]
- Nour, V.; Trandafir, I.; Ionica, M.E. Ascorbic acid, anthocyanins, organic acids and mineral content of some black and red currant cultivars. Fruits 2011, 66, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Barney, D.L.; Hummer, K.E. Currants, Gooseberries, and Jostaberries: A Guide for Growers, Marketers, and Researchers in North America, 1st ed.; CRC Press: Boca Raton, FL, USA, 2005; p. 266. [Google Scholar]
- State Variety Register. Available online: https://reestr.gossortrf.ru (accessed on 30 April 2021).
- Royal Horticultural Society. Available online: https://www.rhs.org.uk (accessed on 30 April 2021).
- Almeida, J.R.M.; D’Amico, E.; Preuss, A.; Carbone, F.; de Vos, C.H.R.; Deiml, B.; Mourgues, F.; Perrotta, G.; Fischer, T.C.; Bovy, A.G.; et al. Characterization of major enzymes and genes involved in flavonoid and proanthocyanidin biosynthesis during fruit development in strawberry (Fragaria ×ananassa). Arch. Biochem. Biophys. 2007, 465, 61–71. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Flavonoids are determinants of freezing tolerance and cold acclimation in Arabidopsis thaliana. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Brennan, R.M. Currants and gooseberries. In Fruit Breeding Temperate Fruit Crop Breeding; Janick, J., Moore, J.N., Eds.; Wiley: Hoboken, NJ, USA, 1996; Volume 2, pp. 191–295. [Google Scholar]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pervaiz, T.; Songtao, J.; Faghihi, F.; Haider, M.S.; Fang, J. Naturally occurring anthocyanin, structure, functions and biosynthetic pathway in fruit plants. J. Plant Biochem. Physiol. 2017, 5, 1–9. [Google Scholar] [CrossRef]
- Starkevič, P.; Ražanskienė, A.; Starkevič, U.; Kazanavičiūtė, V.; Denkovskienė, E.; Bendokas, V.; Šikšnianas, T.; Rugienius, R.; Stanys, V.; Ražanskas, R. Isolation and analysis of anthocyanin pathway genes from Ribes genus reveals MYB gene with potent anthocyanin-inducing capabilities. Plants 2020, 9, 1078. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis. Plant Signal. Behav. 2014, 9, 1–7. [Google Scholar] [CrossRef]
- Khadgi, A.; Weber, C.A. RNA-Seq Analysis of Prickled and Prickle-Free Epidermis Provides Insight into the Genetics of Prickle Development in Red Raspberry (Rubus ideaus L.). Agronomy 2020, 10, 1904. [Google Scholar] [CrossRef]
- Pikunova, A.V.; Knyazev, S.D.; Bakhotskaya, A.Y.; Kochumova, A.A. Microsatellite loci polymorphism in russian black currant (Ribes nigrum L.) varieties from collection of all-russian research institute of breeding fruit crops. Sel’skokhozyaistvennaya Biologiya 2015, 50, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Liebhard, R.; Koller, B.; Gianfranceschi, L.; Gessler, C. Creating a saturated reference map for the apple (Malus × domestica Borkh.) genome. Theor. Appl. Genet. 2003, 106, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Pluta, S.; Broniarek-Niemiec, A.; Żurawicz, E. Productive value of eighteen gooseberry (Ribes grossularia L.) cultivars of different origin evaluated in Central Poland. J. Fruit Ornam. Plant Res. 2010, 18, 197–209. [Google Scholar]
- Duman, R.; Doğan, H.H.; Dinç, M.; Tuncer, P. Cytotoxic and antiviral activity of Ribes uva-crispa Linn. and Ribes multiflorum Kit. ex Romer and Schultes extracts. Int. J. Pharm. Sci. Res. 2018, 9, 1779–1787. [Google Scholar] [CrossRef]
- Antonius, K.; Karhu, S.; Kaldmäe, H.; Lacis, G.; Rugenius, R.; Baniulis, D.; Sasnauskas, A.; Schulte, E.; Kuras, A.; Korbin, M.; et al. Development of the Northern European Ribes core collection based on a microsatellite (SSR) marker diversity analysis. Plant Genet. Resour. 2012, 10, 70–73. [Google Scholar] [CrossRef]
- Mezhnina, O.A.; Urbanovich, O.Y. Study of genetic variability of representatives of Ribes L. grown in Belarus. EuroBiotech J. 2017, 1, 184–187. [Google Scholar] [CrossRef] [Green Version]
- Brennan, R.; Jorgensen, L.; Woodhead, M.; Russell, J. Development and characterization of SSR markers in Ribes species. Mol. Ecol. Notes 2002, 2, 327–330. [Google Scholar] [CrossRef]
- Martins, W.S.; Lucas, D.C.S.; Neves, K.F.S.; Bertioli, D.J. WebSat—A web software for microsatellite marker development. Bioinformation 2009, 3, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Nunes, C.F.; Ferreira, J.L.; Fernandes, M.C.N.; Breves, S.D.S.; Generoso, A.L.; Soares, B.D.F.; Dias, M.S.C.; Pasqual, M.; Borem, A.; Cançado, G.M.D.A. An improved method for genomic DNA extraction from strawberry leaves. Ciência Rural 2011, 41, 1383–1389. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Grandin, U. PC-ORD version 5: A user-friendly toolbox for ecologists. J. Veg. Sci. 2006, 17, 843–844. [Google Scholar] [CrossRef]
- Ahmad, A.; Wang, J.-D.; Pan, Y.-B.; Rahat Sharif, R.; Gao, S.-J. Development and use of simple sequence repeats (SSRs) markers for sugarcane breeding and genetic studies. Agronomy 2018, 8, 260. [Google Scholar] [CrossRef] [Green Version]
- Çelenk, A.G.; Özcan, T. Characterisation of some Ribes L. accessions from Turkey based on SSRs patterns. Int. J. Environ. Agric. Res. 2016, 2, 12–19. [Google Scholar]
- Hackett, C.A.; Russell, J.; Jorgensen, L.; Gordon, S.L.; Brennan, R.M. Multi-environment QTL mapping in blackcurrant (Ribes nigrum L.) using mixed models. Theor. Appl. Genet. 2010, 121, 1483–1488. [Google Scholar] [CrossRef]
- Robinson, D.O.; Roeder, A.H. Themes and variations in cell type patterning in the plant epidermis. Curr. Opin. Genet. Dev. 2015, 32, 55–65. [Google Scholar] [CrossRef]
- Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Lo Piero, A.R. Expression analysis in response to low temperature stress in blood oranges: Implication of the flavonoid biosynthetic pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef]
- Clark, J.R.; Finn, C.E. Blackberry Breeding and Genetics. Fruit Veg. Cereal Sci. Biotechnol. 2011, 5, 27–43. [Google Scholar]
- Saigo, T.; Wang, T.; Watanabe, M.; Tohge, T. Diversity of anthocyanin and proanthocyanin biosynthesis in land plants. Curr. Opin. Plant Biol. 2020, 55, 93–99. [Google Scholar] [CrossRef]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB-bHLH-WD repeat (MBW) pigment regulatory model: Addition of a WRKY factor and co-option of an anthocyanin MYB for betalain regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, L.; Grando, M.S.; Sordo, M.; Grisenti, M.; Martens, S.; Giongo, L. Establishment of molecular markers for germplasm management in a worldwide provenance Ribes spp. collection. Plant Omics 2013, 6, 165–174. [Google Scholar]
- Wang, H.; Fan, W.; Li, H.; Yang, J.; Huang, J.; Zhang, P. Functional characterization of dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, X.; Cai, X.; Shan, X.; Gao, R.; Yang, S.; Han, T.; Wang, S.; Wang, L.; Gao, X. Dihydroflavonol 4-reductase genes from Freesia hybrida play important and partially overlapping roles in the biosynthesis of flavonoids. Front. Plant Sci. 2017, 8, 428. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, V.G.; Subbotina, N.M.; Maluchenko, O.P.; Lebedeva, T.N.; Krutovsky, K.V.; Shestibratov, K.A. Transferability and polymorphism of SSR markers located in flavonoid pathway genes in Fragaria and Rubus species. Genes 2020, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Hülskamp, M. Evolutionary analysis of MBW function by phenotypic rescue in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Wang, J.; Hu, Z.; Zhao, T.; Yang, Y.; Chen, T.; Yang, M.; Yu, W.; Zhang, B. Genome-wide analysis of bHLH transcription factor and involvement in the infection by yellow leaf curl virus in tomato (Solanum lycopersicum). BMC Genomics 2015, 16, 39. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.-A.; Song, J.Y.; Choi, H.-R.; Chung, J.-W.; Jeon, Y.-A.; Lee, J.-R.; Ma, K.-H.; Lee, M.-C. Novel microsatellite markers acquired from Rubus coreanus Miq. and cross-amplification in other Rubus species. Molecules 2015, 20, 6432–6442. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Wu, S.; Niu, E.; Cheng, C.; Guo, W. Identification of genes related to salt stress tolerance using intron-length polymorphicmarkers, associationmapping and virus-induced gene silencing in cotton. Sci. Rep. 2017, 7, 528. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, V.G.; Subbotina, N.M.; Maluchenko, O.P.; Krutovsky, K.V.; Shestibratov, K.A. Assessment of genetic diversity in differently colored raspberry cultivars using SSR markers located in flavonoid biosynthesis genes. Agronomy 2019, 9, 518. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cultivar | Abbr. | Genetic Origin and Background | Degree of Spinosity | RHS Hardiness Rating | Fruit Color | Origin |
|---|---|---|---|---|---|---|
| Avenarius | Ave | - | Medium | H7 | red | Russia |
| Angliyskiy Zeleniy | AnZ | - | High | H7 | yellow-green | Russia |
| Chorny Negus | ChN | Anibut × R.cuccirubra | High | H7 | black | Russia (Michurinsk) |
| Chernomor | Che | 2152 × mixed pollen (Phenicia, Zeleniy Butylochniy, Brazilskiy, Seyanets Maurera) | Lower | H7 | black | Russia (Michurinsk) |
| Consul (Senator) | Con | Chelyabinskiy Zeleniy × Afrikanets | Lower | H6 | dark red | Russia (Southern Urals) |
| Chelyabinskiy Zeleniy | ChZ | Houghton × Angliyskiy Zholtiy | Medium | H7 | green | Russia (Southern Urals) |
| Grushenka | Gru | Severniy Kapitan × Moskovskiy Krasniy × GF 595-33 | Lower | H7 | nearly black | Russia (Moscow) |
| Invicta | Inv | Keepsake × (Whinham’s Industry ×Resistentia) | Medium | H7 | pale green | England, U.K. |
| Kolobok | Kol | Rozoviy-2 × Smena | Lower | H5 | dark red | Russia (Moscow) |
| Kursu Dzintars | KuD | Stern Razhig × Pellervo | High | H7 | yellow | Latvia |
| Laskoviy | Las | Neslukhovskiy × Kolobok | Lower | H7 | red | Russia (St. Peterburg) |
| Lepaan Red | LeR | - | Medium | H7 | dark red | Finland |
| Malahit | Mal | Chorny Negus × Phenicia | Lower | H7 | bright green | Russia (Michurinsk) |
| Moskovskiy Krasniy | MoK | Avenarius (open pollination) | Lower | H6 | dark red | Russia (Moscow) |
| Nezhniy | Nez | - | Lower | H6 | pale green | Russia |
| Rozoviy-2 | Ro2 | Seyanets Lefora×Phenicia | Medium | H6 | dark red | Russia (Moscow) |
| Russkiy | Rus | Careless× mixed pollen (Houghton, Oregon, Karri, Shtamboviy) | Medium | H6 | dark red | Russia (Michurinsk) |
| Russkiy Zholtiy | RuZ | Bud mutation of Russkiy | Medium | H6 | yellow | Russia (Michurinsk) |
| Severniy Kapitan | SeK | 310-24 × Rozoviy-2 | Lower | H7 | nearly black | Russia (Moscow) |
| Seyanets Lefora | SeL | Eduard Lefor (open pollination) | Medium | H7 | red | Russia (Vologda Region) |
| Triumfalniy | Tri | - | Medium | H7 | yellow | Russia |
| Vladil | Vla | Chelyabinskiy Zeleniy × Afrikanets | Lower | H6 | red | Russia (Southern Urals) |
| Locus | Gene | Species | NCBI GenBank Accession Number | Motif and Number of Repeats | Location in the Gene | PCR Primer Nucleotide Sequence | Allele Size, bp | ||
|---|---|---|---|---|---|---|---|---|---|
| Forward | Reverse | Expected | Observed | ||||||
| RucANS | anthocyanidin synthase (ANS) | Ribes uva-crispa | LN736353 | (TA)6 (AGTGA)2 | Intron/exon | TCTTAACCCTAAAATTGCAGCC | CCATTCCACCAACTTCTTTCTC | 236 | 237, 241 |
| RucHLH-1 | bHLH3 | Ribes uva-crispa | LN736347 | (ACTA)3 | Intron | TTTCACTAGAGCCATTCTTGCC | GAAAATACGTTCACGATGGAGC | 208 | 198, 208 |
| RucHLH-2 | bHLH3 | Ribes uva-crispa | LN736347 | (T)10 | Intron | TTTTCTCTTCCTCGTGTTGCTC | CCCTCTCTGTAGTGCCAAATTC | 245 | 243, 247, 248 |
| RucHLH-3 | bHLH3 | Ribes uva-crispa | LN736347 | (TTTCTC)2 | Intron | GAATTTGGCACTACAGAGAGGG | TGAAGTTGAGTGTTCGGAGAGA | 317 | 314, 318, 319 |
| RucHLH-4 | bHLH3 | Ribes uva-crispa | LN736347 | (AGA)5 (GAG)4 | Exon | CGTAAACCCTAACCGAGTCATC | ATTATTTGAAGCGTCGTCGG | 247 | 246 |
| RucWD-1 | WD40 | Ribes uva-crispa | LN736318 | (CCA)4 | Exon | CTTCTCGCCTACACCATCAAG | AAGAGTTGGGTTGGAACATGAG | 191 | 189 |
| RucWD-2 | WD40 | Ribes uva-crispa | LN736318 | (CCAAC)2 | Exon | CGACGAAACCCTAAGCATAAAG | CAAGCAATGTCGTAAACCTCCT | 372 | 378 |
| RucWD-3 | WD40 | Ribes uva-crispa | LN736318 | (TTTGA)2 | Exon | CCACGATAAGGAGGTTTACGAC | CCATCAAAATAGTCGCCATGTA | 203 | 204 |
| RucDFR1-1 | dihydroflavonol 4-reductase (DFR) | Ribes uva-crispa | LN736355 | (AAGAA)2 (AAAC)3 | Intron | AATAGGACGGAGGGAGTACGA | ACTTCATTCTGCCAAGTAGGGT | 396 | 399 |
| RucDFR1-2 | dihydroflavonol 4-reductase (DFR) | Ribes uva-crispa | LN736355 | (A)15 (TTTTC)2 | Intron | ACCCTACTTGGCAGAATGAAGT | CGTGGTCTTCGACACAAAATAC | 362 | 364, 370, 375, 377, 388, 390, 393, 397 |
| RucDFR1-3 | dihydroflavonol 4-reductase (DFR) | Ribes uva-crispa | LN736355 | (T)19 | Intron | CTAGTGGTTGGTCCTTTCATCA | CTAGGCTGGTCCCTAAATCGTA | 329 | 318, 320, 323, 324, 326 |
| RucDFR1-4 | dihydroflavonol 4-reductase (DFR) | Ribes uva-crispa | LN736355 | (ATGTT)2 | Intron | CGTGTATGAGAACCCTAAAGCC | CTTTTCGTCAATGCCCTTAAAC | 323 | 326 |
| RucDFR2-1 | dihydroflavonol 4-reductase (DFR) | Ribes uva-crispa | LN736356 | (TTAATT)2 (A)14 | Intron | CTATATCGTTCGAGCAACCGTA | TGGCAAGTCTAACAAATGCTTC | 259 | 257, 258, 259, 261 |
| RucUFGT | UDP glucose flavonoid 3-O-glycosyltransferase-like protein (UFGT) | Ribes uva-crispa | LN736351 | (TTGGAG)2 (TTTAT)2 (GCATA)2 (TATGC)2 (TTA)4 (TATAG)2 | Exon/Intron | GTGCTCATGTTTATACCGACTTCA | CAAAGCAAAGGGAAGAGGTTG | 351 | 343, 346, 355 |
| RucCHS-1 | chalcone synthase (CHS) | Ribes uva-crispa | LN736358 | (GCAAC)2 | Exon | CACGAATCCACTTGTGTGTTTT | ATTTAGGTTGACCCCATTCCTT | 343 | 343 |
| RucCHS-2 | chalcone synthase (CHS) | Ribes uva-crispa | LN736358 | (CACGT)2 | Exon | GAAGTGGGCCTTACATTTCATC | CTCGTCCAAGATAAACAACACG | 267 | 268 |
| RnMYB-1 | transcription factor MYB 10 | Ribes nigrum | LN736314 | (TA)11 | Intron | AGTGGTTTCGGAGTTAGGA | TCCAGGAATTCTTCCAGCAA | 340–350 | - |
| LfMYB-1 | MYB113 | Liquidambar formosana | KU987934 | (AAATA)2 (AAAATA)1,8 (AAAAATA)2 (AAATAG)1,6 | Intron | ATTAGGAATGGAGGAGGCTAGG | CATGAGCTGGTTCACTTGACAT | 353 | - |
| LfMYB-2 | MYB113 | Liquidambar formosana | KU987934 | (AGTGAA)2 (GCCTCG)2 | Exon | GAAAGATAGTGCCCAGAAGACG | TCTAAGGGAAACTCATTCCAGC | 375 | - |
| RJL-2 | microsatellite DNA, clone RJL-2 | Ribes nigrum | AJ439089 | (AG)11 | - | CGAAGGTTGAATCGGTGAGT | TGTGGAACTACGTGGCTACG | 207 | 222, 228, 231, 234, 236, 240, 243, 254 |
| RJL-6 | microsatellite DNA, clone RJL-6 | Ribes nigrum | AJ439093 | (CT)12(TTCA)3(CT)6 | - | TGTTCCCTGTTTCCTTCAAAA | GGACGTGGACGATGAGAGTT | 291 | 283, 285, 287, 291, 293, 295, 301 |
| Locus | Location in the Gene | Major Allele Frequency | Number of Alleles | Heterozygosity | Polymorphism Information Content (PIC) | |
|---|---|---|---|---|---|---|
| Expected (He) | Observed (Ho) | |||||
| RucDFR2-1 | intron | 0.39 | 4 | 0.71 | 0.95 | 0.66 |
| RucDFR1-2 | intron | 0.45 | 8 | 0.69 | 0.91 | 0.65 |
| RucDFR1-3 | intron | 0.48 | 5 | 0.67 | 0.77 | 0.63 |
| RucANS | intron/exon | 0.98 | 2 | 0.04 | 0.05 | 0.04 |
| RucUFGT | intron/exon | 0.93 | 3 | 0.13 | 0.14 | 0.12 |
| RucHLH2 | intron | 0.80 | 3 | 0.34 | 0.41 | 0.31 |
| RucHLH3 | intron | 0.64 | 3 | 0.52 | 0.73 | 0.47 |
| RJL-2 | - | 0.48 | 8 | 0.69 | 0.86 | 0.65 |
| RJL-6 | - | 0.39 | 7 | 0.74 | 1.00 | 0.71 |
| Mean | - | 0.61 | 4.8 | 0.51 | 0.65 | 0.47 |
| Cultivar Name | Allele Size, bp | Locus |
|---|---|---|
| Angliyskiy Zeleniy | 393 | RucDFR1-2 |
| Angliyskiy Zeleniy | 234 | RJL-2 |
| Angliyskiy Zeleniy | 287 | RJL-6 |
| Malahit | 240 | RJL-2 |
| Nezhniy | 241 | RucANS |
| Nezhniy | 355 | RusUFGT |
| Nezhniy | 231 | RJL-2 |
| Kolobok | 222 | RJL-2 |
| Seyanets Lefora | 388 | RucDFR1-2 |
| Triumfalniy | 397 | RucDFR1-2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidyagina, E.O.; Lebedev, V.G.; Subbotina, N.M.; Treschevskaya, E.I.; Lebedeva, T.N.; Shestibratov, K.A. The Development of the Genic SSR Markers for Analysis of Genetic Diversity in Gooseberry Cultivars. Agronomy 2021, 11, 1050. https://doi.org/10.3390/agronomy11061050
Vidyagina EO, Lebedev VG, Subbotina NM, Treschevskaya EI, Lebedeva TN, Shestibratov KA. The Development of the Genic SSR Markers for Analysis of Genetic Diversity in Gooseberry Cultivars. Agronomy. 2021; 11(6):1050. https://doi.org/10.3390/agronomy11061050
Chicago/Turabian StyleVidyagina, Elena O., Vadim G. Lebedev, Natalya M. Subbotina, Ella I. Treschevskaya, Tatyana N. Lebedeva, and Konstantin A. Shestibratov. 2021. "The Development of the Genic SSR Markers for Analysis of Genetic Diversity in Gooseberry Cultivars" Agronomy 11, no. 6: 1050. https://doi.org/10.3390/agronomy11061050
APA StyleVidyagina, E. O., Lebedev, V. G., Subbotina, N. M., Treschevskaya, E. I., Lebedeva, T. N., & Shestibratov, K. A. (2021). The Development of the Genic SSR Markers for Analysis of Genetic Diversity in Gooseberry Cultivars. Agronomy, 11(6), 1050. https://doi.org/10.3390/agronomy11061050