
Effect and Compatibility of Phosphite with Trichoderma sp. Isolates in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbiological Material
2.2. Plant Material
2.3. Effect of Phi on In Vitro Fusarium sp. Growth
2.4. Compatibility of Trichoderma Isolates with Phi
2.5. In Vitro Compatibility of Phi with Trichoderma sp. in the Control of Fusarium sp.
2.6. Compatibility of Phi and Trichoderma sp. in the Control of Fusarium sp. in Sugarcane Plants
2.7. Experimental Design
2.8. Statistical Analysis
3. Results
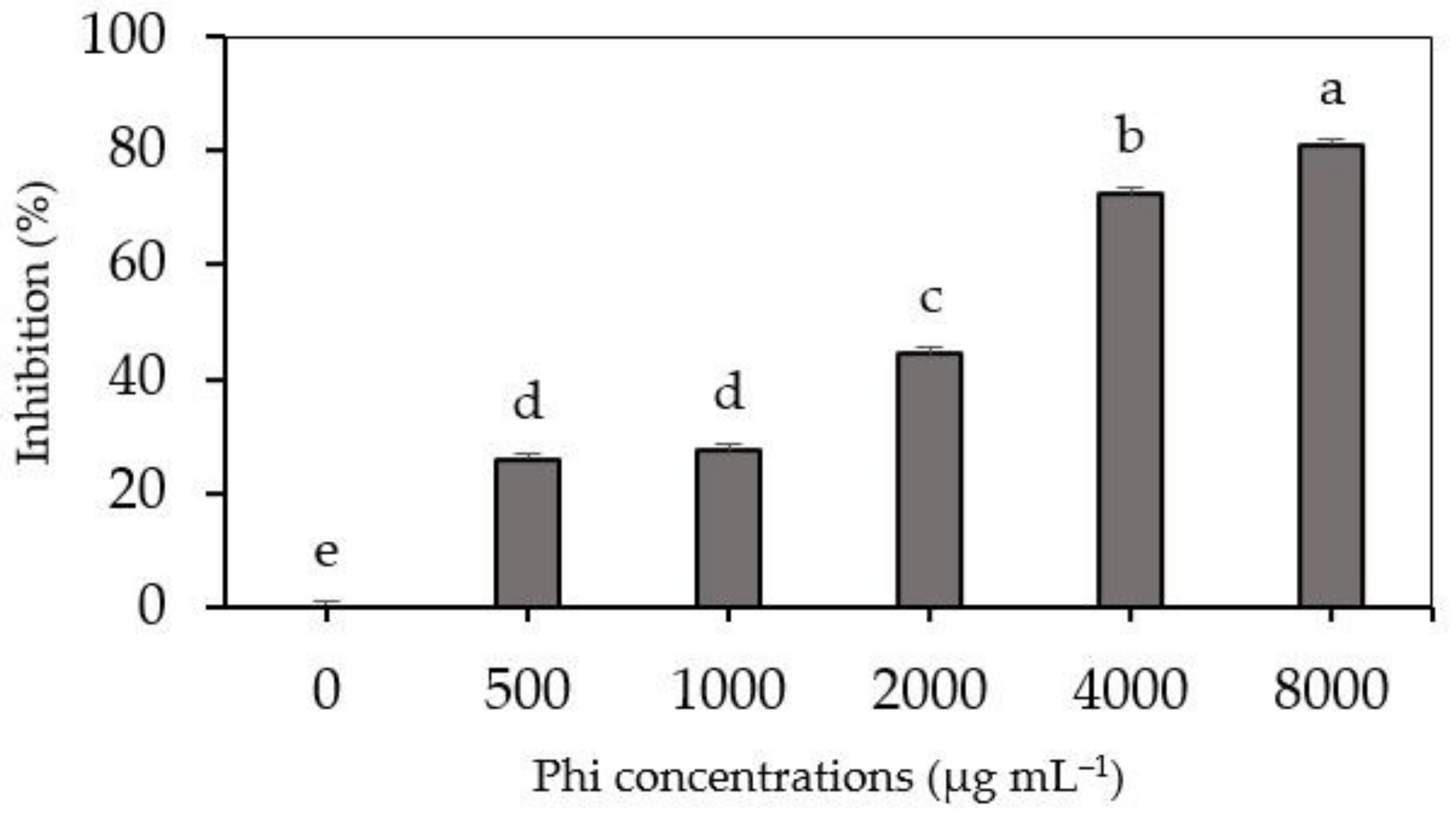
3.1. Effect of Phi on In Vitro Fusarium sp. Growth
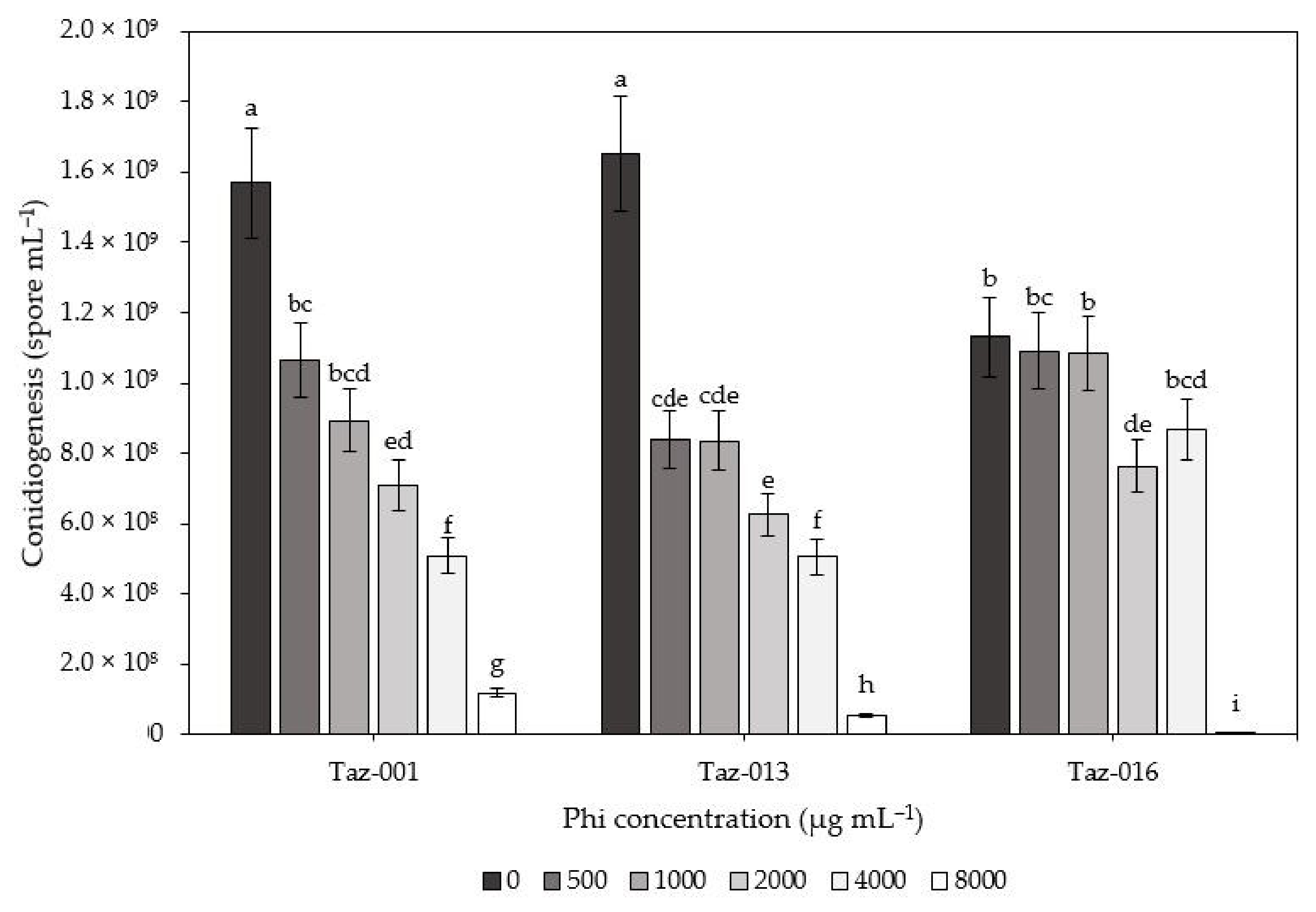
3.2. Compatibility of Trichoderma Isolates with Phi
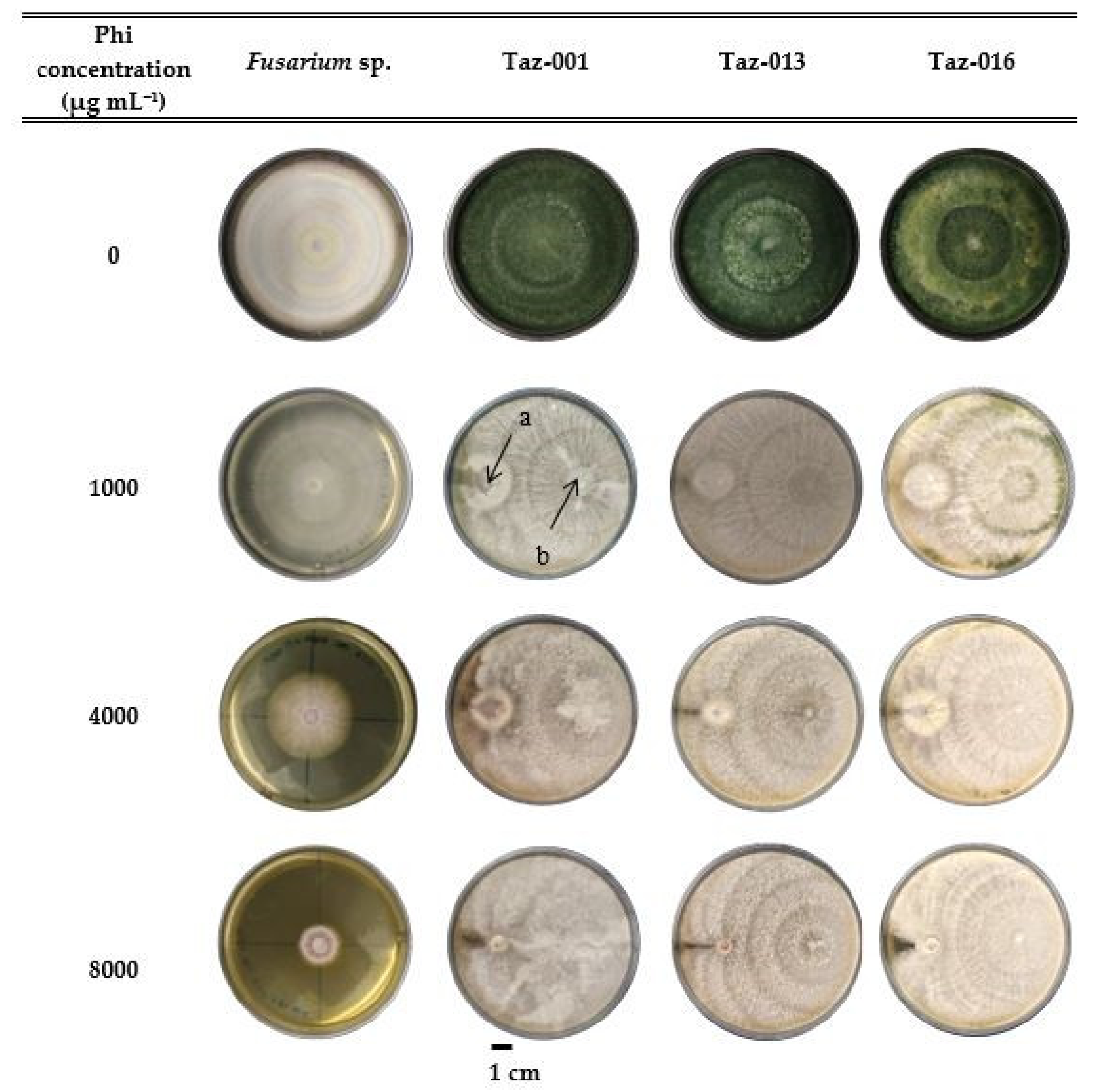
3.3. In Vitro Compatibility of Phi with Trichoderma sp. in the Control of Fusarium sp.
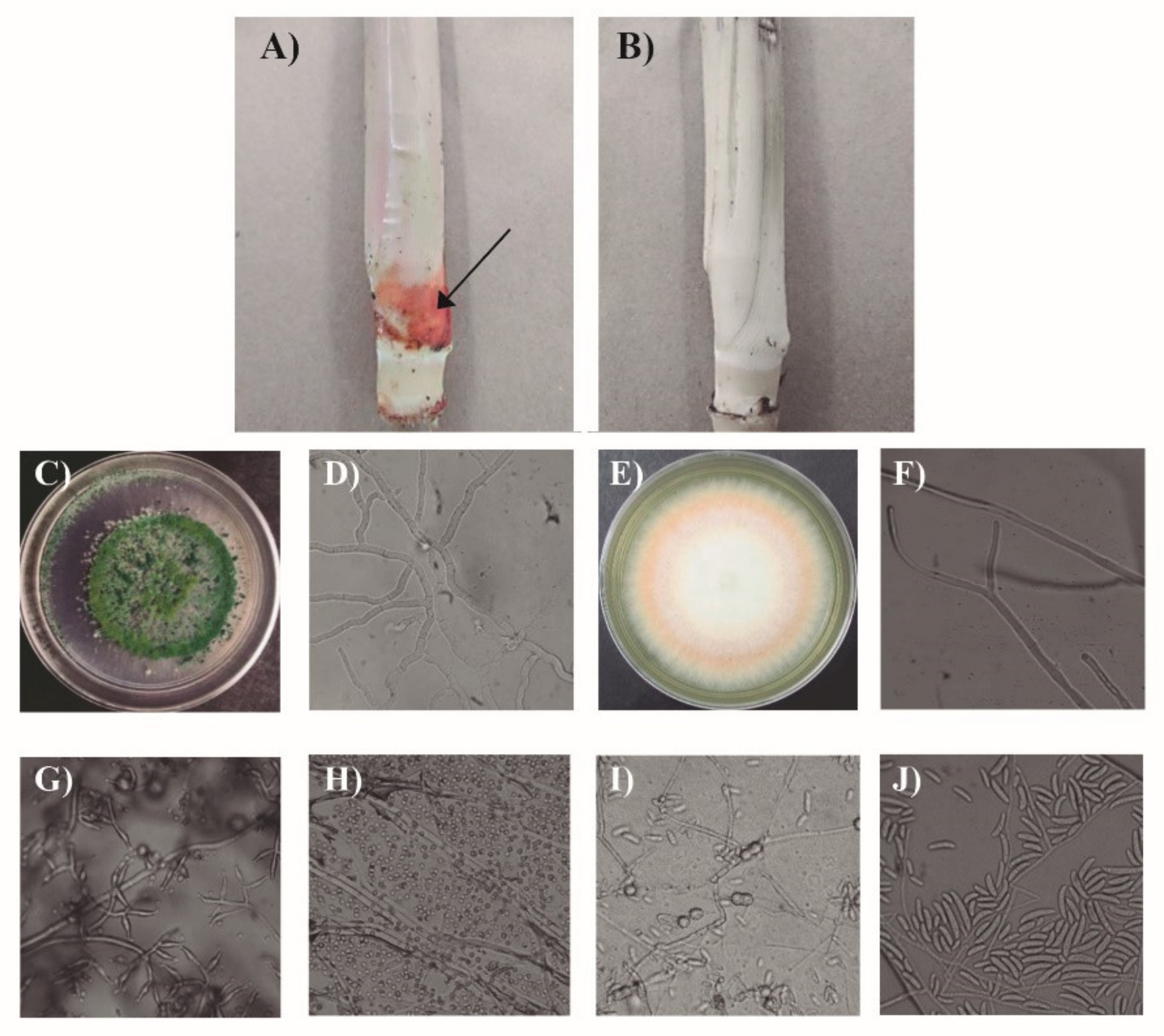
3.4. Compatibility of Phi and Trichoderma sp., in the Control of Fusarium sp. in Sugarcane Plants
4. Discussion
4.1. Phi Significantly Decreases the Growth of the Fusarium Species Complex In Vitro
4.2. Compatibility between Trichoderma and Phi
4.3. Phi Is Compatible with Trichoderma sp. in In Vitro Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane
4.4. The Combination of Phi and Trichoderma sp. Is Efficient in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane in the Planta Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, P.H.; Paterson, A.H.; Tew, T. Sugarcane: The Crop, the Plant, and Domestication. In Sugarcane: Physiology, Biochemistry, and Functional Biology; John Wiley & Sons Ltd.: Chichester, UK, 2013; pp. 1–17. [Google Scholar]
- Sentíes-Herrera, H.E.; Trejo-Téllez, L.I.; Gómez-Merino, F.C. The Mexican Sugarcane Production System: History, Current Status, and New Trends. In Sugarcane: Production Systems, Uses and Economic Importance; Murphy, R., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2017; pp. 39–71. [Google Scholar]
- FAO. Food and Agriculture Organization of the United Nations. FAO Stats. Crops. Sugarcane. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 9 April 2021).
- Gómez-Merino, F.C.; Trejo-Téllez, L.; Salazar-Ortiz, J.; Pérez-Sato, J.A.; Sentíes-Herrera, H.E.; Bello-Bello, J.J.; Aguilar-Rivera, N. Diversification of the Sugar Agroindustry as a Strategy for Mexico. Agroproductividad 2017, 10, 7–12. [Google Scholar]
- Aguilar-Rivera, N.; Michel-Cuello, C.; Serna-Lagunes, R.; de Jesús Debernardi-Vázquez, T.; Trujillo-Mata, A. Ethanol Production from the Mexican Sugar Industry: Perspectives and Challenges. In Sugarcane Biofuels; Springer: Cham, Switzerland, 2019; pp. 203–235. [Google Scholar]
- Azevedo, R.A.; Carvalho, R.F.; Cia, M.C.; Gratão, P.L. Sugarcane Under Pressure: An Overview of Biochemical and Physiological Studies of Abiotic Stress. Trop. Plant Biol. 2011, 4, 42–51. [Google Scholar] [CrossRef]
- Savario, C.F.; Hoy, J.W. Microbial Communities in Sugarcane Field Soils with and without a Sugarcane Cropping History. Plant Soil 2011, 341, 63–73. [Google Scholar] [CrossRef]
- Dinolfo, M.I.; Castañares, E.; Stenglein, S.A. Fusarium–Plant Interaction: State of the Art–A Review. Plant Sci. 2017, 53, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S. Pathogenomics and Management of Fusarium Diseases in Plants. Pathogens 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, J.; Wang, H.; Bao, Y.; Li, Y.; Govindaraju, M.; Yao, W.; Chen, B.; Zhang, M. Molecular Characterization of Carbendazim Resistance of Fusarium species Complex that Causes Sugarcane Pokkah Boeng Disease. BMC Genomics 2019, 20, 115. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Handique, P.J.; Deka, S. Rhamnolipid Biosurfactant against Fusarium sacchari—The Causal Organism of Pokkah Boeng Disease of Sugarcane. J. Basic Microbiol. 2014, 54, 548–557. [Google Scholar] [CrossRef] [PubMed]
- McGovern, R.J. Management of Tomato Diseases Caused by Fusarium oxysporum. Crop Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Sevastos, A.; Kalampokis, I.F.; Panagiotopoulou, A.; Pelecanou, M.; Aliferis, K.A. Implication of Fusarium graminearum Primary Metabolism in its Resistance to Benzimidazole Fungicides as Revealed by 1H NMR Metabolomics. Pestic. Biochem. Physiol. 2018, 148, 50–61. [Google Scholar] [CrossRef]
- Patel, P.; Shah, R.; Joshi, B.; Ramar, K.; Natarajan, A. Molecular Identification and Biocontrol Activity of Sugarcane Rhizosphere Bacteria against Red Rot Pathogen Colletotrichum falcatum. Biotechnol. Rep. 2019, 21, e00317. [Google Scholar] [CrossRef]
- Oyarburo, N.S.; Machinandiarena, M.F.; Feldman, M.L.; Daleo, G.R.; Andreu, A.B.; Olivieri, F.P. Potassium Phosphite Increases Tolerance to UV-B in Potato. Plant Physiol. Biochem. 2015, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Merino, F.C.; Trejo-Tellez, L.I. Conventional and Novel Uses of Phosphite in Horticulture: Potentialities and Challenges. Italus Hortus 2016, 23, 1–13. [Google Scholar]
- Li, Y.; Sun, R.; Yu, J.; Saravanakumar, K.; Chen, J. Antagonistic and Biocontrol Potential of Trichoderma asperellum ZJSX5003 against the Maize Stalk Rot Pathogen Fusarium graminearum. Indian J. Microbiol. 2016, 56, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Massoud, K.; Barchietto, T.; le Rudulier, T.; Pallandre, L.; Didierlaurent, L.; Garmier, M.; Ambard-Bretteville, F.; Seng, J.M.; Saindrenan, P. Dissecting Phosphite-Induced Priming in Arabidopsis Infected with Hyaloperonospora arabidopsidis. Plant Physiol. 2012, 159, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liljeroth, E.; Lankinen, Å.; Wiik, L.; Burra, D.D.; Alexandersson, E.; Andreasson, E. Potassium Phosphite Combined with Reduced Doses of Fungicides Provides Efficient Protection against Potato Late Blight in Large-Scale Field Trials. Crop Prot. 2016, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Martínez, S. Effects of Combined Application of Potassium Phosphite and Fungicide on Stem and Sheath Disease Control, Yield, and Quality of Rice. Crop Prot. 2016, 89, 259–264. [Google Scholar] [CrossRef]
- Cerqueira, A.; Alves, A.; Berenguer, H.; Correia, B.; Gómez-Cadenas, A.; Diez, J.J.; Monteiro, P.; Pinto, G. Phosphite Shifts Physiological and Hormonal Profile of Monterey Pine and Delays Fusarium circinatum Progression. Plant Physiol. Biochem. 2017, 114, 88–99. [Google Scholar] [CrossRef]
- Trejo-Téllez, L.I.; Gómez-Merino, F.C. Phosphite as an Inductor of Adaptive Responses to Stress and Stimulator of Better Plant Performance. In Biotic and Abiotic Stress Tolerance in Plants; Springer: Singapore, 2018; pp. 203–238. [Google Scholar]
- Sharma, V.; Salwan, R.; Sharma, P.N. The Comparative Mechanistic Aspects of Trichoderma and Probiotics: Scope for Future Research. Physiol. Mol. Plant Pathol. 2017, 100, 84–96. [Google Scholar] [CrossRef]
- Amiri, A.; Bompeix, G. Control of Penicillium Expansum with Potassium Phosphite and Heat Treatment. Crop Prot. 2011, 30, 222–227. [Google Scholar] [CrossRef]
- Lopes, L.F.; Cruz, A.F.; de Barreto, M.L.A.; de Vasconcelos, T.M.M.; Blum, L.E.B. Post-Harvest Treatment with Ca-Phosphite Reduces Anthracnose without Altering Papaya Fruit Quality. J. Hortic. Sci. Biotechnol. 2018, 93, 272–278. [Google Scholar] [CrossRef]
- Deliopoulos, T.; Kettlewell, P.S.; Hare, M.C. Fungal Disease Suppression by Inorganic Salts: A Review; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1059–1075. [Google Scholar]
- Kiran, G.V.M.; Thara, S.S.; Jyothi, K.R. Studies on Compatibility of Biocontrol Agents with Chemical Fungicides for Integrated Management of Alternaria Leaf Spot of cabbage. J. Pharmacogn. Phytochem. 2018, 7, 2974–2977. [Google Scholar]
- Widmer, T.L. Compatibility of Trichoderma asperellum Isolates to Selected Soil Fungicides. Crop Prot. 2019, 120, 91–96. [Google Scholar] [CrossRef]
- Dhanya, M.K.; Anjumol, K.B.; Murugan, M.; Deepthy, K.B. Compatibility of Trichoderma viride and Pseudomonas fluorescens with Plant Protection Chemicals and Fertilizers in Cardamom. J. Trop. Agric. 2016, 54, 129–135.h. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, MN, USA, 1998. [Google Scholar]
- Pandey, D.K.; Tripathi, N.N.; Tripathi, R.D.; Dixit, S.N. Fungitoxic and Phytotoxic Properties of the Essential Oil of Hyptis suaveolens. J. Plant Dis. Prot. 1982, 89, 344–349. [Google Scholar]
- Reuveni, M.; Sheglov, D.; Cohen, Y. Control of Moldy-Core Decay in Apple Fruits by β-Aminobutyric Acids and Potassium Phosphites. Plant Dis. 2003, 87, 933–936. [Google Scholar] [CrossRef] [Green Version]
- D’Addazio, V.; Ahnert dos Santos, R.A.; Bastos Leitão, A.S.; Barreto da Silva, M.; Alves Fernandes, A.; Falqueto, A.R. Evaluation of In Vitro Inhibition of Mycelial Growth of Fusarium solani f. sp. piperis by Different Products in Brazil. Afr. J. Microbiol. Res. 2016, 10, 1992–1998. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.A.F.; de Moura, K.E.; de Moura, K.E.; Salomão, D.; Patricio, F.R.A. Compatibility of Trichoderma Isolates with Pesticides Used in Lettuce Crop. Summa Phytopathol. 2018, 44, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Chaparro, A.P.; Carvajal, L.H.; Orduz, S. Fungicide Tolerance of Trichoderma asperelloides and T. harzianum Strains. Agric. Sci. 2011, 2, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a New Biocontrol Gene in Trichoderma atroviride: The Role of an ABC Transporter Membrane Pump in the Interaction with Different Plant-Pathogenic Fungi. Mol. Plant Microbe Interact. 2009, 22, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Perveen, K.; Bokhari, N.A. Antagonistic Activity of Trichoderma harzianum and Trichoderma viride Isolated from Soil of Date Palm Field against Fusarium oxysporum. Afr. J. Microbiol. Res. 2012, 6, 3348–3353. [Google Scholar] [CrossRef]
- Bagwan, N.B. Evaluation of Trichoderma Compatibility with Fungicides, Pesticides, Organic Cakes and Botanicals for Integrated Management of Soil Borne Disease of Soybean [Glycine max (L.) Merril]. Int. J. Plant Prot. 2010, 3, 206–209. [Google Scholar] [CrossRef]
- Tapwal, A.; Kumar, R.; Gautam, N.; Pandey, S. Compatibility of Trichoderma viride for Selected Fungicides and Botanicals. Int. J. Plant Pathol. 2012, 3, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Živković, S.; Stojanović, S.; Ivanović, Ž.; Gavrilović, V.; Popović, T.; Balaž, J. Screening of Antagonistic Activity of Microorganisms against Colletotrichum acutatum and Colletotrichum gloeosporioides. Arch. Biol. Sci. 2010, 62, 611–623. [Google Scholar] [CrossRef]
- Svistova, I.D.; Senchakova, T.Y. Ecological Plasticity of Trichoderma Fungi in Leached Chernozem. Eurasian Soil Sci. 2010, 43, 314–315. [Google Scholar] [CrossRef]
- Infante, D.; Martínez, B.; González, N.; Reyes, Y. Mecanismos de Acción de Trichoderma Frente a Hongos Fitopatógenos. Rev. Protección Veg. 2009, 24, 14–21. [Google Scholar]
- Groves, E.; Howard, K.; Hardy, G.; Burgess, T. Role of Salicylic Acid in Phosphite-Induced Protection against Oomycetes—A Phytophthora Cinnamomic-Lupinus augustifolius Model System. Eur. J. Plant Pathol. 2015, 141, 559–569. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.Q.; Wang, M.; Sun, J.; Gao, J.X.; Chen, J. Effect of Trichoderma harzianum on Maize Rhizosphere Microbiome and Biocontrol of Fusarium stalk Rot. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Chittenden, C.; Singh, T. In Vitro Evaluation of Combination of Trichoderma harzianum and Chitosan for the Control of Sapstain Fungi. Biol. Control. 2009, 50, 262–266. [Google Scholar] [CrossRef]
- Nallathambi, P.; Umamaheswari, C.; Thakore, B.B.L.; More, T.A. Post-Harvest Management of Ber (Ziziphus mauritiana Lamk) Fruit Rot (Alternaria alternata Fr. Keissler) Using Trichoderma Species, Fungicides and Their Combinations. Crop Prot. 2009, 28, 525–532. [Google Scholar] [CrossRef]
- Abd-El-Khair, H.; Elshahawy, I.E.; Haggag, H.E.K. Field Application of Trichoderma spp. Combined with Thiophanate-Methyl for Controlling Fusarium solani and Fusarium oxysporum in Dry Bean. Bull. Natl. Res. Cent. 2019, 43, 19. [Google Scholar] [CrossRef] [Green Version]
- El-Komy, M.H.; Saleh, A.A.; Eranthodi, A.; Molan, Y.Y. Characterization of Novel Trichoderma asperellum Isolates to Select Effective Biocontrol Agents against Tomato Fusarium Wilt. Plant Pathol. J. 2015, 31, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Abhiram, P.; Masih, H. In Vitro Antagonism of Trichoderma viride against Fusarium oxysporum Strains. J. Pharmacogn. Phytochem. 2018, 7, 2816–2819. [Google Scholar]
- Akrami, M.; Golzary, H.; Ahmadzadeh, M. Evaluation of Different Combinations of Trichoderma Species for Controlling Fusarium Rot of Lentil. Afr. J. Biotechnol. 2011, 10, 2653–2658. [Google Scholar] [CrossRef]
- Sharma, P. Complexity of Trichoderma-Fusarium Interaction and Manifestation of Biological Control. Aust. J. Crop Sci. 2011, 5, 1027–1038. [Google Scholar]
- Tapwal, A.; Singh, U.; da Silva, J.A.T.; Singh, G.; Garg, S.; Kumar, R. In Vitro Antagonism of Trichoderma viride against Five Phytopathogens. Pest Technol. 2011, 5, 59–62. [Google Scholar]
- Cherkupally, R.; Amballa, H.; Reddy, B.N. In Vitro Antagonistic Activity of Trichoderma Species against Fusarium oxysporum f. sp. melongenae. Int. J. Appl. Agric. Res. 2017, 12, 87–95. [Google Scholar]
- Suleiman, A.S.; Gambo, M.S.; Sunusi, M. An In Vitro Antagonistic Effect of Trichoderma spp. against Fusarium oxysporum f. sp. lycopersici. FUDMA J. Sci. 2019, 3, 369–374. [Google Scholar]
- Veenstra, A.; Rafudeen, M.S.; Murray, S.L. Trichoderma asperellum Isolated from African Maize Seed Directly Inhibits Fusarium verticillioides Growth in vitro. Eur. J. Plant Pathol. 2019, 153, 279–283. [Google Scholar] [CrossRef]
- Verma, M.; Brar, S.K.; Tyagi, R.D.; Surampalli, R.Y.; Valéro, J.R. Antagonistic Fungi, Trichoderma spp.: Panoply of Biological Control. Biochem. Eng. J. 2007, 37, 1–20. [Google Scholar] [CrossRef]
- Mishra, V.K. In Vitro Antagonism of Trichoderma Species against Pythium aphanidermatum. J. Phytol. 2010, 2, 28–35. [Google Scholar]
- Rajeswari, P.; Kannabiran, B. In Vitro Effects of Antagonistic Microorganisms on Fusarium oxysporum [Schlecht. Emend. Synd & Hans] Infecting Arachis hypogaea L. J. Phytol. 2011, 3, 83–85. [Google Scholar]
- Suneeta, P.; Kumar, S.V.; Eraivan, K.; Aiyanathan, A.; Nakkeeran, S. Promissory Action of Trichoderma spp. and Fungicides in the Management of Fusarium Wilt of Gerbera. J. Pure Appl. Microbiol. 2017, 11, 241–247. [Google Scholar] [CrossRef]
- Singh, R.; Biswas, S.K.; Nagar, D.; Singh, J.; Singh, M.; Kumar Mishra, Y. Sustainable Integrated Approach for Management of Fusarium Wilt of Tomato Caused by Fusarium oxysporum f. sp. lycopersici (Sacc.) Synder & Hansen. Sustain. Agric. Res. 2015, 4. [Google Scholar] [CrossRef]
- Mwangi, M.W.; Muiru, W.M.; Narla, R.D.; Kimenju, J.W.; Kariuki, G.M. Management of Fusarium oxysporum f. sp. lycopersici and Root-Knot Nematode Disease Complex in Tomato by Use of Antagonistic Fungi, Plant Resistance and Neem. Biocontrol Sci. Technol. 2019, 29, 229–238. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Treatments | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 | T13 | T14 | |
| Trichoderma sp. | − | − | − | − | − | − | + | + | + | + | + | + | + | − |
| Phi (µg mL−1) | − | 500 | 1000 | 2000 | 4000 | 8000 | − | 500 | 1000 | 2000 | 4000 | 8000 | − | − |
| Fusarium sp. | + | + | + | + | + | + | + | + | + | + | + | + | − | − |
| Phi Concentration (µg mL−1) | Conidiogenesis (Spores mL−1) |
|---|---|
| 0 | 1.3 × 109 ± 0.3 × 109 a |
| 500 | 3.0 × 106 ± 0.7 × 106 b |
| 1000 | 3.2 × 106 ± 0.7 × 106 b |
| 2000 | 1.1 × 106 ± 0.3 × 106 c |
| 4000 | 1.0 × 106 ± 0.2 × 106 c |
| 8000 | 9.0 × 105 ± 0.2 × 105 c |
| Treatment | Plant Height (cm) | Culm Length (cm) | Leaf Dry Weight (g) |
|---|---|---|---|
| Fus (T1) | 132.6 ± 4.7 ef | 27.6 ± 2.0 d | 10.6 ± 2.3 c |
| 500 Phi vs. Fus (T2) | 144.2 ± 4.3 cd | 37.2 ± 1.9 ab | 15.0 ± 2.1 c |
| 1000 Phi vs. Fus (T3) | 138.6 ± 4.7 edef | 28.6 ± 2.0 cd | 10.5 ± 2.3 c |
| 2000 Phi vs. Fus (T4) | 130.8 ± 4.7 f | 28.7 ± 2.0 cd | 10.5 ± 2.3 c |
| 4000 Phi vs. Fus (T5) | 130.3 ± 4.3 f | 27.8 ± 1.9 d | 10.3 ± 2.1 c |
| 8000 Phi vs. Fus (T6) | 136.5 ± 4.3 ef | 30.0 ± 1.9 cd | 9.8 ± 2.1 c |
| Tri vs. Fus (T7) | 139.2 ± 4.7 cdef | 32.4 ± 2.0 bcd | 14.8 ± 2.3 bc |
| Tri + 500 Phi vs. Fus (T8) | 162.8 ± 4.3 a | 40.7 ± 1.9 a | 20.4 ± 2.1 ba |
| Tri + 1000 Phi vs. Fus (T9) | 149.3 ± 4.3 bc | 32.5 ± 1.9 bcd | 21.4 ± 2.1 a |
| Tri + 2000 Phi vs. Fus (T10) | 147.3 ± 4.3 cd | 33.3 ± 1.9 bc | 23.9 ± 2.1 a |
| Tri + 4000 Phi vs. Fus (T11) | 163.4 ± 4.7 a | 41.6 ± 2.0 a | 26.1 ± 2.2 a |
| Tri + 8000 Phi vs. Fus (T12) | 141.6 ± 4.7 cdef | 31.4 ± 2.0 cd | 22.5 ± 2.2 a |
| Tri (T13) | 160.6 ± 4.7 ab | 32.6 ± 2.0 bcd | 15.1 ± 2.2 bc |
| Control (T14) | 148.0 ± 4.7 bcd | 31.8 ± 2.0 bcd | 14.0 ± 2.2 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solis-Palacios, R.; Hernández-Ramírez, G.; Salinas-Ruiz, J.; Hidalgo-Contreras, J.V.; Gómez-Merino, F.C. Effect and Compatibility of Phosphite with Trichoderma sp. Isolates in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane. Agronomy 2021, 11, 1099. https://doi.org/10.3390/agronomy11061099
Solis-Palacios R, Hernández-Ramírez G, Salinas-Ruiz J, Hidalgo-Contreras JV, Gómez-Merino FC. Effect and Compatibility of Phosphite with Trichoderma sp. Isolates in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane. Agronomy. 2021; 11(6):1099. https://doi.org/10.3390/agronomy11061099
Chicago/Turabian StyleSolis-Palacios, Rocio, Gabriela Hernández-Ramírez, Josafhat Salinas-Ruiz, Juan Valente Hidalgo-Contreras, and Fernando Carlos Gómez-Merino. 2021. "Effect and Compatibility of Phosphite with Trichoderma sp. Isolates in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane" Agronomy 11, no. 6: 1099. https://doi.org/10.3390/agronomy11061099
APA StyleSolis-Palacios, R., Hernández-Ramírez, G., Salinas-Ruiz, J., Hidalgo-Contreras, J. V., & Gómez-Merino, F. C. (2021). Effect and Compatibility of Phosphite with Trichoderma sp. Isolates in the Control of the Fusarium Species Complex Causing Pokkah Boeng in Sugarcane. Agronomy, 11(6), 1099. https://doi.org/10.3390/agronomy11061099