1. Introduction
The
Taraxacum genus is a well-known medicinal plant; however, limited propagation studies have been carried out since the interest in
Taraxacum koksaghyz as a natural rubber source during World War II came to an end. In the last decades, propagation studies of this genus have gained relevance due to the necessity of natural medicines that are spreading worldwide. In this sense,
Taraxacum is now being considered to be a potential source of interesting medicinal compounds because several compounds targeted for their anticancer, antibacterial, antioxidants, and anti-inflammatory properties are present in its tissues, such as terpenoids and phenolics [
1,
2]. In vitro propagation methods are widely used on plants that have difficulties in cultivation or with low bioactive compounds extraction yields. Among in vitro cultures, callus cultures provide rapid plant multiplication and allow us to manipulate the environment for producing specific compounds or for increasing their yields [
3]. The impact that has been achieved through this in vitro plant cultivation method for producing valuable commercially important products which include pigments (e.g., anthocyanins and betacyanins), anti-inflammatory agents (e.g., berberine and rosmarinic acid), and molecules (e.g., paclitaxel and podophyllotoxin) [
4,
5].
In this environment, callus culture is influenced by internal (explant type, plant age, and genotype) and external factors (growth medium, temperature, and light), among others [
6,
7]. For example, the formulation of the growth medium affects the initiation of the culture, as well as the growth rate, biomass production, and the synthesis of specific products [
8]. The selection of carbohydrates as carbon sources and energy influence cell biomass accumulation, due to low CO2 levels in closed cultures during the photoperiod [
9]. Light influences both growth and secondary metabolism in plants, for example, by regulating the diurnal pattern of monoterpenoid synthase transcript levels and monoterpenoid production [
10]. It has been demonstrated that the application of different combinations and concentrations of PGRs on
Bidens pilosa were able to manipulate the metabolome, producing in vitro specialized metabolites for pharmacological purposes [
11].
The proper establishment of the in vitro culture is necessary to obtain successful long-term results, by preventing organ regeneration, promoting callus friability and viability, and maintaining the cells growing at a high rate, with high cell viability and minimum organogenesis [
6]. For a successful in vitro culture establishment, proper explant selection for callus induction and suitable conditions required for callus survival and growth need to be determined [
12]. Once the suitable explant is selected, the conditions for the induction are established, with the medium composition being the first step. Sugars have been reported as necessary for biomass accumulation because they act as a source of energy (since the photosynthetic activity of in vitro growing tissues is usually reduced), carbon supply for biosynthetic processes and cell wall synthesis [
13], and they are also signal molecules repressing or activating plant genes [
14]. In cell cultures, plant growth regulators (PGRs) are added to the basal growth medium to stimulate and control plant development [
15,
16]. Among these, especially auxins and cytokinins act synergistically to control cell division in undifferentiated cells [
17] and induce callogenesis and further callus proliferation. Theoretically, it has been postulated that equal amounts of auxin and cytokinin promote callus induction; however, in practice, this differs to a great extent due to variations in the endogenous levels of phytohormones in individual plants [
18]. The mode of interaction between these PGRs varies with the plant species and organs being studied [
19]. Light also has a significant effect on callus growth and morphogenesis as it influences the rate of cell division in different plants and directly controls plant growth and development [
20]. Studies on
Taraxacum tissue culture are limited. Some authors have studied optimal culture media for shoot formation [
21]; other authors have also tried to induce callus for shoot regeneration, but the results have not been very efficient [
22]; the transformation of this species by means of
Agrobacterium rhizogenes has also been studied to obtain transgenic plants and improve the accumulation of secondary metabolites of interest [
23].
Therefore, there are no studies on the Taraxacum genus that determine the impact of several simultaneous nutritional and physical factors through factorial designs. This is the first study describing an experimental design approach that includes multiple physical factors (explant type and light) and biological factors (carbohydrate source and plant growth regulators) in three experiments. Our results allowed us to establish an efficient protocol for the induction and maintenance of Taraxacum officinale callus in vitro.
The aim of this study was to investigate the effects of the explant type, carbon source, PGRs, and light on an in vitro callus culture of T. officinale in order to determine an effective protocol for callus induction and maintenance to be used in further in vitro culture programs.
3. Results
3.1. Callus Induction
Explant type (hypocotyls, roots, stems, and leaves), light, carbon source type (glucose and sucrose) and concentration (from 1.0 to 5.5%), and PGR treatments (NAA and BAP in combination) significantly affected the evaluated parameters for callus induction (
p < 0.05) both in quantity and quality. However, light did not affect the fresh and dry weight of the callus (
p < 0.05). Moreover, carbon source (CS) did not affect the time of callusing and the interaction of CS type × CS concentration did not affect the friability parameter (
p < 0.05). The MANOVA results from Experiments 1, 2 and 3 regarding the effect of light, explant type, carbon source, and PGR on callus induction from
T. officinale are presented in
Tables S4 and S5.
The effects of explant type and photoperiod on callogenesis parameters, morphology, and organogenesis, at the fourth week after sowing, are presented in
Table 1. As shown, the callus response, morphologies, and physical characteristics of calli derived from the different explants under light (18/6 h light/dark) or dark (0/24 h light/dark) conditions were found to be different. Hypocotyls and root explants showed the maximum percentage of callogenesis (100%), while leaves were also suitable, but showed less induction with percentages of 73% and 56% under light and dark conditions, respectively. A high degree of callus formation was observed for root explants, especially under complete darkness, with an average value of 98 ± 1%; under light, these explants showed slightly lower results (ca. 5%) than in the dark. Leaf explants showed a higher difference in callus formation when light conditions were tested, with values of 60 ± 7% and 41 ± 1% under dark and light conditions, respectively. This parameter was not applied for hypocotyls because callus only grew from one side of the explant, covering it completely. The induction period (or the time of callusing) was lower for hypocotyls and roots, with average values of 7 and 5 days under light and dark, respectively. For leaves, at least two weeks were needed to observe significant callus masses for light and dark conditions. Nearly 15% of leaf explants that turned dark died under light conditions during the first week, while stems did not develop any callus masses during the entire period. Under dark conditions, leaf explants increased their survival to 94 ± 7%, while stems did not respond to the induction conditions. On the contrary, hypocotyls and roots were completely viable, and none of the explants presented partial or total necrosis under light and dark conditions.
Callus color varied because of the explant type and the light regime. Under light conditions, green cells were observed in all the calli derived from the explants; however, pale green calli predominated when derived from hypocotyls, green when derived from roots, and dark green when derived from leaves. Under dark conditions, yellow calli predominated in the three types of explants, followed by pale green calli, and to a lesser extent, green calli from leaves explants. Stem explants lost their natural green color until becoming translucent without showing any type of cell growth. Callus color was observed as directly related to the friability of the cell masses, in which pale green and green colors were observed for the most friable calli, especially for root- and hypocotyl-derived calli. Under light, these calli presented a friability of 86 ± 6% and 60 ± 8%, respectively. Both cell masses were soft on the surface and in the inside, but the callus from roots turned more compact during the course of the experiments while the green color became predominant. The friability of the callus from leaves was lower with a value of 48 ± 6% and showed, from the beginning, a more compact aspect than the callus derived from roots and hypocotyls, sometimes with a soft surface but highly compact in the inside. Under dark, friability was higher than in light conditions and similar for root-derived and hypocotyl-derived calli 87 ± 7% and 85 ± 7%, respectively. For leaf-derived calli, friability had a value of 56 ± 7%. In all cases, calli were soft in the outside and in the inside. The color was similar for all calli, varying in different shades of yellow.
Considering the organogenesis obtained from the beginning of the experiment to the second week after sowing, all calli (in light and dark conditions) produced shoots during the experiment with no exception (100% for all the conditions tested), only differing in the profuseness of the young leaves and in their extent on the callus surface. Viability was maximum (100%) for root-derived and hypocotyl-derived calli, while the viability of leaf-derived calli was 86 ± 5% and 95 ± 3% in light and dark conditions, respectively. No cell masses were observed in the stem explants at the end of the period, indicating null viability in both light and dark conditions.
Fresh weight was higher for the root-derived calli (3.69 ± 0.03 g and 5.87 ± 0.05 g in light and dark conditions, respectively) followed by the hypocotyl-derived calli (2.86 ± 0.33 g and 5.35 ± 0.62 g in light and dark conditions, respectively), while the lowest was obtained for the leaf-derived calli (1.68 ± 0.29 g and 2.54 ± 0.39 g in light and dark conditions, respectively). On the contrary, dry weight was higher for the leaf-derived calli (0.27 ± 0.01 g and 0.20 ± 0.02 g in light and dark conditions, respectively), followed by the hypocotyl-derived calli (0.23 ± 0.01 g and 0.17 ± 0.01 g in light and dark conditions, respectively), while the lowest values were obtained for the root-derived calli (0.14 ± 0.01 g and 0.12 ± 0.02 g in light and dark conditions, respectively). Higher humidity content was calculated for the calli in dark conditions. Calli developed under light presented humidity values of 95 ± 1%, 94 ± 1%, and 83 ± 3% for hypocotyl-, root-, and leaf-derived calli, respectively. Under dark conditions, these explants showed humidity values of 97 ± 1%, 98 ± 1%, and 91 ± 1%, respectively. Fresh and dry weight showed a similar pattern regardless of the light condition (
Table 2).
In
Figure 1, callus induction from
T. officinale leaf explants under light and dark conditions are presented. As shown, leaf explants generated callus were in the principal vein and cutting edges (
Figure 1a). After 4 weeks of culture, leaf regeneration from leaf explants and highly compact calli were observed (
Figure 1b). Developing leaves showed a green color under light conditions but were translucent under dark conditions (
Figure 1c), while callus was yellow and highly friable when explants were kept under complete darkness.
In
Figure 2 callus induction from
T. officinale hypocotyls under different PGRs and carbon source concentrations is presented. Callus derived from roots (
Figure 2a) was considerably distinct from the callus derived from hypocotyls (
Figure 2b), the latter being more friable and pale green. Root explants showed a high degree of callus formation in which cell masses were all over the explant, especially compact in the outside wall of the primary root and more friable on the cutting side. For the hypocotyl explants, callus was also observed friable on the cutting side. A similar pattern was observed in calli grown under dark conditions; however, cell masses were generally friable and characteristically yellow (
Figure 1c) regardless of the type of explant.
3.2. Effect of Carbon Source on Callus Induction from Hypocotyls of T. officinale
In general, sucrose gave slightly better results than glucose in callogenesis at concentrations of 3.2 and 5.5%, while at concentrations of 1.0 and 2.3%, no differences were observed and maximum callogenesis was achieved (100%) for both sugars. Callogenesis in sucrose was 98 ± 4% and 69 ± 10% at 3.2% and 5.5% of the carbon source, respectively. In glucose, values were 83 ± 3% and 58 ± 9% at 3.2% and 5.5% of the carbon source, respectively. Induction time was not influenced by the carbon source, with an average value of 5.9 ± 1.0 days, regardless of whether glucose or sucrose was present in the medium. Callus color varied with the carbon source concentrations but did not vary regarding the carbon source type. In general, a pale-yellow color was observed at the lowest concentrations (1.0% and 2.3%), while a darker color (shades of orange and brown) was observed at higher concentrations (3.2% and 5.5%). This color change was found related to the friability of the callus. Sucrose proved better than glucose on the friability parameter at every concentration tested, while friability was also related to the carbon source concentration. This parameter was higher at 2.3%, with values of 91 ± 3% and 75 ± 4% for sucrose and glucose, respectively. For sucrose, values of 69 ± 6%, 78 ± 8%, and 41 ± 4% were obtained at 1.0%, 3.2%, and 5.5% of the carbon source, respectively. For glucose, values of 52 ± 8%, 57 ± 6%, and 28 ± 6% were obtained at 1.0%, 3.2%, and 5.5% of the carbon source, respectively (
Table 3).
Similarly, to what was observed in the previous experiment, organogenesis was present in all of the explants at every condition tested with no exceptions (100% of organogenesis at all carbon source conditions tested) showing only shoot proliferation. For sucrose and glucose, viability was almost 100% at 1.0% and 2.3% of the carbon source. Viability decreased at higher carbon source concentrations to 87 ± 4% and 45 ± 7% for sucrose 3.2% and 5.5%, respectively. For glucose, viability was 69 ± 11% and 44 ± 7% at 3.2% and 5.5%, respectively.
Fresh weight was greater for sucrose than glucose (up to 30% lower) at every carbon source concentration. The highest fresh weight was obtained at sucrose 2.3% with an average value of 1.24 ± 0.11 g, while at 1.0%, 3.2%, and 5.5% of the carbon source, fresh weight was 1.02 ± 0.07, 0.71 ± 0.02, and 0.44 ± 0.10 g, respectively. For glucose, values of 0.76 ± 0.04, 1.00 ± 0.11, 0.65 ± 0.08, and 0.43 ± 0.10 g were obtained at 1.0%, 2.3%, 3.2%, and 5.5% of the carbon source, respectively. Dry weight was similar for sucrose and glucose at 2.3%, 3.2%, and 5.5% of the carbon source, with average values of 0.08 ± 0.01, 0.07 ± 0.01, and 0.06 ± 0.01 g, respectively. At 1.0% of the carbon source, dry weight on sucrose was higher than on glucose, with values of 0.07 ± 0.01 and 0.04 ± 0.01 g, respectively Humidity also decreased at higher carbon source concentrations. However, this parameter was not influenced by the carbon source type, being statistically similar for sucrose and glucose, with a maximum value of 94 ±1% when the medium was supplemented with 1.0% or 2.3% of the carbon source. Minimum values of 85 ± 2% and 87 ± 2% were obtained at sucrose 5.5% and glucose 5.5%, respectively (
Table 4).
3.3. Effect of PGRs on Callus Induction from Hypocotyls of T. officinale
PGR treatments were evaluated considering the best conditions selected from Experiment 1 (hypocotyl-derived callus induced in dark conditions) and Experiment 2 (medium supplemented with sucrose 2.3%).
Developmental and morphological parameters of
T. officinale callus derived from hypocotyls induced under different NAA/BAP concentrations are presented in
Table 5. Hypocotyl explants showed diverse responses when different concentrations of NAA and BAP in combination were used. The maximum callusing value of 93 ± 3% was noticed on the hypocotyl explants grown under darkness on medium supplemented with 0.5 mg/L of BAP and NAA each, while less callogenesis was observed when higher BAP concentrations (3.0 mg/L) and lower NAA concentrations (0.05 mg/L) were in the medium. The minimal callus induction, with a value of 10 ± 4%, was induced in the basal medium without the addition of PGR, in which a high percentage of the explants (75 ± 2%) showed shoot proliferation by direct organogenesis. Regarding the addition of PGRs, callogenesis showed the faster induction time with an average value of 5.1 ± 0.7 days at 0.5 mg/L BAP and 0.5 mg/L of NAA in combination, while induction times were up to 15 days for higher BAP concentrations, such as 3.0 mg/L of BAP and 0.05 mg/L of NAA. The control condition (no PGR) showed the minimum induction time, with values of 4.5 ± 0.6 days. At this condition, small callus appeared on the explants, but they were highly compact and showing a fast regeneration, covering the callus with shoots less than 5 days after induction.
Organogenesis was total for all the PGR combinations evaluated and similar to what was observed in the two previous experiments; only shoots were developed profusely when organogenesis was reached. Nevertheless, when NAA and BAP were added to the medium in equal concentrations of 0.5 mg/L each, the callus surface with shoot presence was considerably lower (<30%) than when the NAA/BAP ratios were different than 1.0, and especially when NAA was added at 0.05 mg/L. The color was affected by the BAP and NAA combination, turning from pale green at 0.5 mg/L of BAP and 0.5 mg/L of NAA to dark green at 3.0 mg/L of BAP and 0.05 mg/L of NAA. Brown calli were only observed when no PGR was present in the medium. As observed under different carbon source conditions in the previous experiment, friability was affected by the BAP and NAA concentrations. At 0.5 mg/L of BAP and 0.5 mg/L of NAA, calli with higher friability were observed, with a value of 88.0 ± 8.9%, while at 3.0 mg/L of BAP and 0.05 mg/L of NAA, only a 30 ± 3% of friability was obtained. For the control, only highly compact calli were developed before they rapidly showed shoot regeneration. The final weight was higher at 0.5 mg/L of BAP and 0.5 mg/L of NAA, with an average value of 3.6 ± 1.7 g. The viability parameter was strongly affected with the BAP and NAA combination, with maximum values (100%) at lower concentrations of BAP, decreasing while BAP increased in the medium with and average value of 87.2 ± 8.6% at 3.0 mg/L of BAP and 0.05 mg/L of NAA. Moreover, a similar value was observed for the control, with a viability percentage of 87 ± 3%.
Fresh weight was higher at 0.05 mg/L of NAA and 0.05 mg/L of BAP with a value of 2.75 ± 0.41 g at the second week after induction. At this condition, the humidity was maximum with a value of 94 ± 2%, giving a dry weight of 0.16 ± 0.05 g. In general, fresh weight and humidity results were lower when the medium was supplemented with 0.05 mg/L of NAA or there were no PGRs in the medium, ranging from 0.2 to 1.0 g and from 0.1 to 0.4 g for fresh and dry weight, respectively, and from 55% to 67% for humidity. On the contrary, higher values were obtained when NAA was supplemented at 0.5 mg/L to the medium, ranging from 1.2 to 2.8 g and from 0.2 to 0.5 g for fresh and dry weight, respectively, and from 71% to 94% for humidity. It is worth noticing that at lower NAA/BAP ratios at a given NAA concentration (0.05, 0.225, and 0.5 mg/L) more cell growth was observed. For instance, in the medium supplemented with 0.05 mg/L of NAA, higher fresh weight was obtained when BAP was added at 0.5 mg/L (0.99 ± 0.13 g), indicating an NAA/BAP ratio of 10. Moreover, in a medium supplemented with 0.225 mg/L NAA, fresh weight was maximum when BAP was added at 0.5 mg/L (1.86 ± 0.17 g) or with an NAA/BAP ratio of 2. Humidity was not observed as related to the NAA/BAP ratio, and therefore other factors should be also influencing this parameter (
Table 6).
3.4. Callus Maintenance
After the callus induction, the cell masses with the best quality in terms of low organogenesis and high viability and friability (identified as the ones induced at sucrose 2.3%, 0.5 mg/L of NAA, 0.5 mg/L of BAP, and kept in darkness) were transferred to fresh medium containing different PGR concentrations (NAA and BAP in combination at equal concentrations).
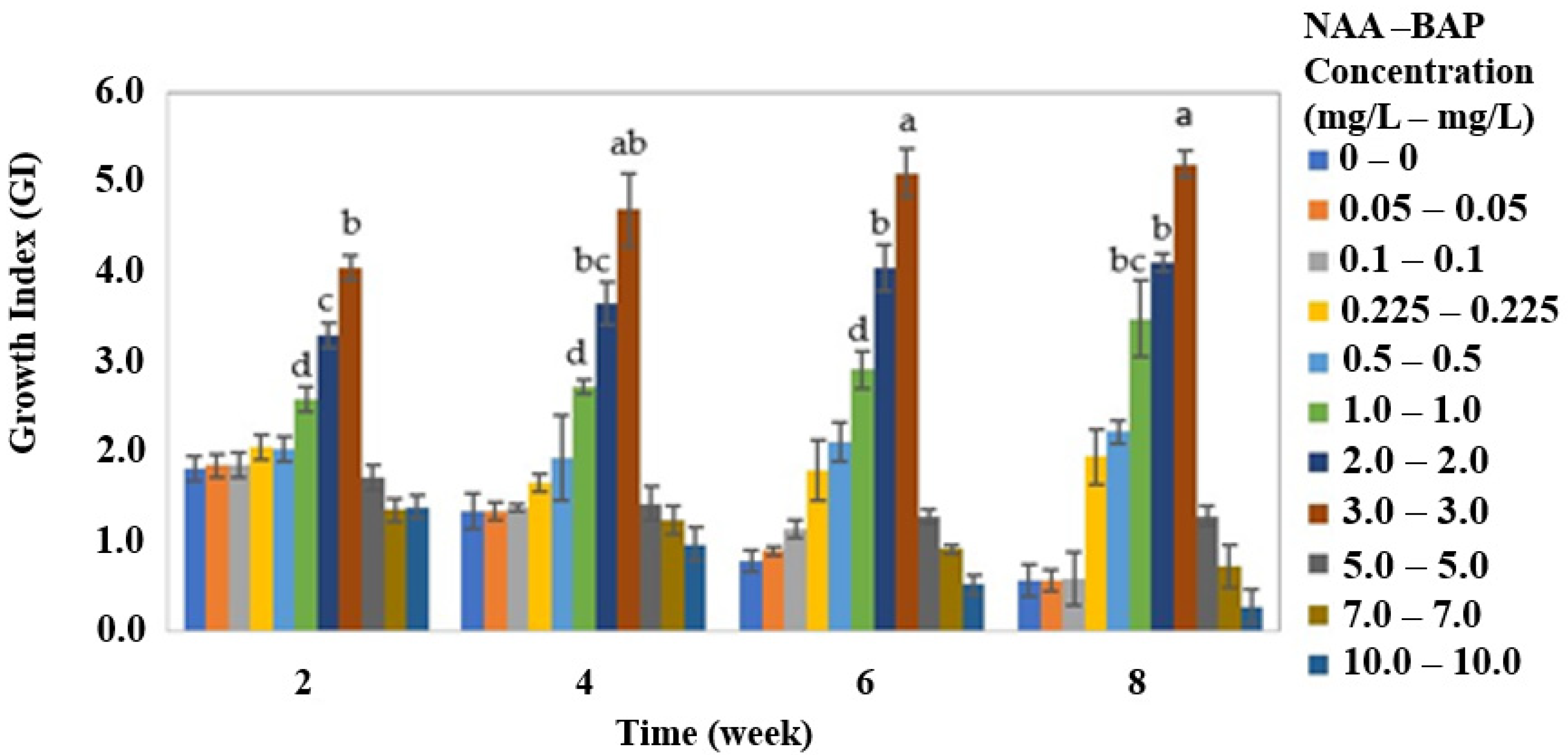
In
Figure 3, the GI of callus culture at the different PGR treatment concentrations is shown. As observed, the increase in the PGR concentration up to 3.0 mg/L of NAA and 3.0 mg/L of BAP was directly related to the increment in the callus proliferation (
p < 0.05). Moreover, a relation between PGR concentration up to 3.0 mg/L and GI in the eighth week was determined. Best callus proliferation was achieved at this concentration, with an average GI value of 5.2 ± 2.4, while above 5.0 mg/L of these PGRs GI was negatively affected, and decreasing until complete callus necrosis was reached at 10.0 mg/L of NAA and 10.0 mg/L of BAP. GI values above 1, indicating cell proliferation at the end of the period (4 months of maintenance) were 4.1 ± 0.1, 3.5 ± 0.4, 2.2 ± 0.2, 1.9 ± 0.1, and 1.3 ± 0.1 for PGR treatments of NAA and BAP at equal amounts of 2.0, 1.0, 0.5, 0.25, and 5.0 mg/L each, respectively. GI values below 1, indicating cell loss at higher rate than cell proliferation, were 0.73 ± 0.2, 0.59 ± 0.3, 0.57 ± 0.2, 0.56 ± 0.2, and 0.27 ± 0.1 for PGR treatments of NAA and BAP of 7.0, 0.1, 0.05, 0.0, and 10.0 mg/L each, respectively.
As observed, PGR treatment (NAA and BAP in combination) did influence the quality of the callus during the maintenance period (
p < 0.05) in all the parameters evaluated. Organogenesis was higher when NAA and BAP were present in the medium at concentrations below 1.0 mg/L of each PGR, with values ranging from 56% to 100%. When supplementing the medium with NAA and BAP from 2.0 to 10.0 mg/L, organogenesis decreased from 38 ± 7% to 6.4 ± 2%. The organogenic ratio also decreased with an increase of PGRs in the medium, from 100% when no PGRs were added to 0% at 7.0 mg/L of NAA and 7.0 mg/L of BAP. At 10.0 mg/L of NAA and 10.0 mg/L of BAP, calli were not able to grow during the evaluated period. The color changed from yellow to brown during the course of the experiment, especially at extreme concentrations of PGRs (0 and 0.5 mg/L, and 5.0 to 10.0 mg/L of NAA and BAP each). At intermediate PGR concentrations (0.1 to 3.0 mg/L NAA and BAP each) only, a normal yellow color was maintained during the four-month period. Friability was also dependent on the NAA and BAP supplementation, showing a maximum of 92 ± 3% at 3.0 mg/L of NAA and 3.0 mg/L of BAP. From this point, friability decreased up to 0% when no PGRs were in the medium and to 32 ± 4% at 7.0 mg/L of NAA and 7.0 mg/L of BAP (
Table 7).
The results indicated that maximum fresh weight and humidity were obtained by supplementing the maintenance medium with equal concentrations of NAA and BAP at 3.0 mg/L each. In this condition, fresh and dry weights were 4.81 ± 0.43 and 0.28 ± 0.20 g, respectively, with a humidity of 95 ± 4%. In general, below and above this NAA/BAP concentration, fresh and dry weights decreased in a direct relation with the PGR content in the medium, up to 0.18 ± 0.03 g FW and 0.06 ± 0.01 g DW when no PGRs were added, and to 0.05 ± 0.01 g FW and 0.03 ± 0.00 g DW when NAA and BAP were supplemented at 10.0 mg/L each. Humidity was also related to the PGR combinations, decreasing from 95% to 66 ± 3% when no PGRs were added, and to the minimum value obtained of 43 ± 7% when NAA and BAP were supplemented at 10.0 mg/L each (
Table 8).
In
Figure 4, calli maintained at different PGR concentrations are shown. As observed, at low PGR concentrations (<1.0 mg/L NAA and BAP in combination) shoots predominated sooner as a lower concentration of PGR was supplemented to the medium (
Figure 4a–c). Intermediate concentrations (1.0–3.0 mg/L of NAA and BAP in combination) gave lower shoot development and less compactness (
Figure 4d–f), while high concentrations (5.0–10.0 mg/L of NAA and BAP in combination) promoted low organogenesis (
Figure 4g–i) but also cell necrosis, by which calli showed total necrosis at 10.0 mg/L. Nevertheless, few calli exposed at high PGR concentrations presented small clusters of new growing cells, perhaps as an adaptation mode. When the callus was transferred into the fresh medium without PGR for monitoring organogenesis capability, the results indicated that at every PGR condition, shoots developed normally between the second and third week of culture in PGR-deficient medium. No significant difference was observed in the response time (
p < 0.05), which points out that this is the period that cells need in order to respond to environmental changes.
4. Discussion
Our findings indicated that hypocotyls and root explants were the most totipotent with the highest percentage of callogenesis (100%) at all the conditions tested and with a profuse callus formation all over the surface (high degree of callus formation), while leaf explants showed less callogenesis than hypocotyls and roots, with moderate or minor calli (less than 66% of the explant surface). These results agree with those reported by several authors who showed that the hypocotyl, root, and cotyledon of
Taraxacum are more callogenic as compared with the leaf [
26,
27,
28,
29]. The latter was not a suitable explant source for callus induction, due to the rapid organogenesis process that was observed as compared with the other explants. Leaves proved to be a proper source when organogenesis was the final goal, rather than for callus maintenance and proliferation. Some authors have agreed that leaf explant was better for callus induction for further shoot regeneration as compared with the other explants [
30].
Considering the response of explants as callus sources, different results can be obtained in the literature depending on the species tested. Even for a unique species, different outcomes can be obtained by modifying the callus induction conditions. Specifically, for
T. officinale, it has been reported that root explants are the best for callus induction, followed by the petiole, and finally, the leaves [
26]. Other authors have reported that, for
Taraxacum, petioles and cotyledons [
27,
28] and shoot tips [
29] are the best source for callus induction, findings which are also in accordance with our results.
In terms of velocity, calli developed two times faster for hypocotyls and roots than for leaf explants under both photoperiods, agreeing with the results reported by Kumar et al. [
31] for cotton callus culture, in which hypocotyl explants showed quick response, within 5–7 days, similar to our results, whereas cotyledon showed slower response in callus induction, despite the PGR concentration tested. An average induction time of 14 days for potato (
Solanum tuberosum L.) callus derived from leaves was reported [
32], similar to what we identified in this work.
We observed that explant type indeed affected the organogenesis capacity of the callus culture, even when shoot proliferation was achieved in all cases. Studies have suggested that the organogenic potential is related to callus structure. Gürel et al. [
33] indicated that white and friable callus of sugar beet was able to produce roots and shoots, while green and compact callus with small cells showed no organogenic capacity. This is contrary to what we observed in this work, in which more compact and greener calli, mainly derived from leaf explants, showed the greatest number of shoots on their surface.
In this study, darkness generated better callogenesis and callus growth than light, considering callus induction percentage, callus quality, and final growth weight (see
Table 5 and
Table S5). For
Taraxacum, no study regarding the effect of light on the callus induction and growth has been reported; however, our results agree with those indicated for callus culture of
Barringtonia racemosa, which showed a maximum fresh weight in the dark as compared with those incubated in light condition [
34]. On the contrary, authors have reported that the fresh weight of rapeseed callus was higher under light, but the browning and necrosis of calli increased, with the growth under dark being more suitable for callus growth [
20]. Some authors indicated that explants of
Licopersicum esculentum, Daucus carota,
Arabidopsis thaliana,
Centella asiatica, and
Beta vularis need to be cultured in dark conditions in order to increase the callus efficiency and reduce the secretion of phenolic compounds, which generally affects the explant survival [
35,
36,
37].
The difference between the response of leaf-derived callus and hypocotyl-derived callus observed in this study could be due to the fact that leaf explants are full of chlorophyll, and therefore dependent on the light environment for cell stimulation, while hypocotyls are less responsive to light and its photomorphogenesis modulation. Nevertheless, the light did affect induction time, with callus induction between 20 to 30% faster under dark conditions. This is opposite to the observations made by Mousavi et al. [
38], who indicated a slightly slower induction time from leaf explants under dark (28–29 days) than under light (26–27 days) for
Eustoma grandiflorum callus. Considering callus weight, Behbahani et al. [
34] indicated that the dry weight of callus of
B. racemosa incubated in darkness was higher as compared with that in the light conditions, which was opposite to our results, in which light promoted dry weight and darkness promoted fresh weight (
Table 5).
Considering organogenesis, morphogenic responses of
T. officinale leaves to light conditions and to the addition of several concentrations of IBA and BAP to the medium have been reported by [
39] who proposed that differentiation seemed to be affected by light and that although light favored differentiation, it did not prove essential. Nevertheless, when cultures were grown in the dark, buds produced albino leaves, and all the explants produced a white and compact callus on the cut surface, especially near the veins. This observation agrees with our outcomes, i.e., when organogenesis was reached under dark, translucent shoots developed (
Figure 1c).
Our study shows that callogenesis and the degree of callus formation from hypocotyls were slightly greater when the medium was supplemented with sucrose rather than with glucose. Moreover, the concentration of the carbon source was extremely important in the quantity and quality of the callus induction and growth (
Table 7). In the literature, no other studies related to the
Taraxacum genus and the effect of carbon source on its callogenesis have been reported. However, our results obtained for hypocotyl-derived callus are in accordance with those published by Rutkowska-Krause et al. [
40], which indicated a positive response of callogenesis to sucrose in callus cultures of flax (
Linum usitatissimum L.). On the contrary, other authors have reported low callus induction and growth using sucrose as compared with other carbon sources [
41].
Callus coloration varied with the carbon source treatments. Similarly, observations made by other authors have indicated that callus of potato turned from yellowish to light green during the incubation time, and finally to dark green under light [
32]. Considering the carbon source type, calli of cotton grown on glucose was less friable than the calli grown on sucrose, while in terms of sugar concentration, calli grown at 2.3% of both carbon sources were more friable, softer, and healthier than the calli grown at 3.2% and 5.5% of the carbon sources; this fact partially agrees with previous reports [
42]. It has been said that sucrose as a carbon source is related to the secretion of phenolic compounds, in which high sucrose concentrations resulted in lower growth and final cell death, while glucose has been proven to control tissue browning of explants in cotton callus induction due to phenolic oxidation as indicated by Kumar et al. [
31]. Koefoed et al. [
43] indicated that callus induction and embryogenic callus formation on shoot apices and immature inflorescences did not differ significantly between media containing sucrose, glucose, fructose, maltose, or a mixture of glucose and fructose; however, callus induction and embryogenic callus formation from leaf explants of
Miscanthus ×
ogiformis Honda ”Giganteus” were best on glucose. This indicates a higher organogenesis rate in glucose, as observed in this study, even when callus induction was similar. Because organ regeneration is not desired for callus proliferation, sucrose seems to be the best carbon source for callus induction under the proposed conditions. Additionally, sucrose is cheaper than glucose, a reason that economically justifies sucrose selection as a carbon source for callus induction experiments.
PGR is the most extensively studied variable for the in vitro growth of the
Taraxacum genus. Several authors have been testing with different PGR types and concentrations; however, these studies have focused more on organ regeneration than on callus proliferation. Theoretically, it has been postulated that equal amounts of auxin and cytokinin promote callus induction; however, in practice, this differs to a great extent, due to variations in the endogenous levels of phytohormones in individual plants of potato [
18]. Thus, in our experiments, PGR treatments were evaluated on callus induction and on callus maintenance in two separate instances. NAA and BAP in combination were critical for callus induction, since no callus was observed when PGRs were absent in the medium (see
Table 2). The highest callus induction and maximum callus weight were achieved when hypocotyls were cultured with 0.5 to 1.25 mg/L of BAP and 0.225 to 0.5 mg/L of NAA in combination. In general, for the
Taraxacum genus, studies have reported the highest callogenesis values (>95%) when cytokinin and auxin are added in concentrations ranging from 0.5 to 2.0 mg/L each. For instance, Chen et al. [
44] indicated that fragile calli can be obtained by using 0.5 mg/L 2,4-D and 2.0 mg/L 6-BA, while Jamshieed et al. [
45] indicated high callusing percentage of
T. officinale leaf explants with IAA and BAP combinations in concentrations ranging from 1.0 to 2.0 mg/L after 4 weeks of induction. Recently, Uteulin et al. [
46] optimized the medium for tissue culture of
T. kok-saghyz by studying callus formation and plant regeneration from leaf explants, indicating that treatment with 1.0 mg/L of BAP and 1.0 mg/L of zeatin was suitable for friable callus production from leaf explants.
In this study, when NAA was present at the lowest concentration (0.05 mg/L, independent of the BAP concentration), green/dark green and highly compact callus were present over the explant. At higher NAA and BAP concentrations (0.225–0.5 mg/L of NAA and 1.25–3.0 mg/L of BAP), callus were lighter green than at the lowest PGR concentrations (0.05 mg/L of NAA and 0.5 mg/L of BAP), which can be explained by the inhibitory effects of auxins on chlorophyll formation, even though the presence of cytokines tend to promote chlorophyll formation [
9].
Considering organogenesis, only shoot development was observed at every PGR condition proposed in this study. Jamshieed et al. [
45] indicated for
T. officinale that low concentrations of auxin and high concentrations of cytokinin always promoted shoot development, as found here, and that the combination of these PGRs is necessary to achieve shoot initiation and elongation. Ermayanti and Martin [
26] pointed out that BAP in higher concentrations than NAA, as our proposed conditions, enhanced shoot proliferation from petioles, leaves, and roots explants. Interestingly, despite the NAA/BAP concentration, no root development was observed during the course of this experiment, even when NAA was indicated as necessary for root induction, whether BAP was present or not, exhibiting a high frequency of rooting in both cotyledon and hypocotyl derived calli of rapeseed [
20], which was contrary to the observations made in this study. The conditions of 0.5 mg/L of NAA and 0.5 mg/L of BAP were selected for further protocols in callus induction.
For callus maintenance, testing different BAP-NAA concentrations at equal amounts resulted in the selection of 3.0 mg/L of NAA and 3.0 mg/L of BAP as conditions that promote cell proliferation in good quality (low organogenesis, high friability, and viability). Concentrations below 1.0 mg/L promote growth towards shoot development, while concentrations above 5.0 mg/L did not allow a proper callus proliferation and total necrosis was observed at 10.0 mg/L. Extreme PGR concentrations can be toxic to T. officinale cell growth, as results evidenced that the growth under these conditions is lower than the growth of calli in the medium even when PGRs are not added.



 and
and 





{kind=link}
{kind=link}
{kind=link}
{kind=link}