1. Introduction
Current global crop production needs to double to meet the projected global demand by the year 2050 with wheat yield gains needing to increase from 0.9 to 2.4% yr
−1 [
1]. Maximizing crop production efficiency and profitability is challenged by market instability [
2] and the increasing production costs are often a critical barrier to achieving these goals. Yield is the primary factor affecting profitability when paired with the efficient use of crop production inputs, such as fertilizers and seed [
3]. Hard red spring wheat (HRSW) grain yield is determined by the complex interaction between genotype, environment, and management [
4]. Environment cannot be fully controlled nor accurately predicted, however, producers can select and anticipate the effect of management practices, such as crop rotation [
5], genotype [
6], seeding date [
7], seeding rate [
5,
7,
8], row spacing [
8], weather and soil management [
9], genetic disease control [
10], and pest management [
11].
Since the advent of hybrid maize (
Zea mays L.) [
12], the exploitation of heterosis as a means of increasing yields in various crops, such as rice (
Oryza sativa L.) and barley (
Hordeum vulgare L.) has been explored [
13,
14,
15]. Interest in exploiting wheat’s heterosis began after male-sterility advances were reported by Kihara [
16], Fukasawa [
17], and Kihara [
18]. Livers and Heyne [
19] found hybridized wheat genotypes yielding 30% more than the best performing inbred cultivar at the time. More recent research has shown a 20% yield improvement in hybrids in comparison to the best commercial cultivar and greater yield stability between environments [
20]. European winter wheat hybrids displayed a higher nitrogen-use efficiency, compared to inbred cultivars [
21]. The adoption of hybrid wheat may be an effective way to increase wheat yield. However, hybrid wheat seed production has been inefficient and costly. Currently, blend hybrids [
22], a mixture of male parent and hybrid seed, may be more cost-effective approaches to exploit the benefits of hybrids.
Seed production costs are an important component in the determination of hybrid seed prices. The additional cost of hybrid technology must be paid for by an increase in revenue from improved yield, nutrient content, or grazing value. Retzlaff [
23] reported wheat hybrid seed costs of USD 0.84 kg
−1 to be five times greater than the average price of USD 0.15 kg
−1 for nonhybrid wheat seed. Hybrid wheat seed has been relatively unavailable in the USA but prices in 2002 were 2.5 times greater for hybrid seed, compared to certified seed [
24]. Cisar and Cooper [
24], nevertheless found that even with its higher cost, hybrid seed use could increase profits by USD 25 ha
−1. Future hybrid seed costs in the USA may offset the expected increase in yield generated from hybrid wheat.
The seeding rate is an integral component of the management practices required for high wheat yields. The optimal seeding rate has been shown to vary between HRSW cultivars (inbred wheat genotype) in eastern North Dakota and western Minnesota [
25]. Guitard et al. [
26] found the seeding rate to be a direct determinant of optimal spikes plant
−1 and yield. Chen et al. [
8] reported the optimum seeding rate of spring wheat to be 2.15 million live seeds ha
−1 in central Montana, whereas Wiersma [
7] found maximum yield was achieved with seeding rates between 4.84−5.31 million live seeds ha
−1 for seven HRSW cultivars in northwest Minnesota. When combined over eight HRSW cultivars, it was concluded that the highest seeding rate of 4.30 million seeds ha
−1 resulted in the highest yield in Saskatchewan, Canada [
27]. Utilizing optimal plant densities limits potential yield loss by reducing light-use inefficiency [
28] and maximizing nutrient use efficiency [
29].
An important crop production goal is for the input use to be economically optimal. Agronomic and economic optimal seeding rates can differ depending on the yield response of a cultivar and the cost of seed used. The agronomic optimum seeding rate (AOSR) describes the point where the maximum yield is obtained, whereas the economic optimum seeding rate (EOSR) usually includes economic factors associated with the seeding rate and yield to find the point at which the maximum net profit is achieved. When the yield followed a quadratic response to seeding rate, the AOSR was found to be 5.43 million seeds ha
−1 and the EOSR to be between 4.24 and 4.83 million seeds ha
−1 for winter wheat in Ohio [
30]. McKenzie et al. [
31] reported an EOSR to range from 2.00 to 2.40 million live seeds ha
−1 for irrigated soft white spring wheat in southern Alberta, Canada. Similarly, Khah et al. [
32] found 2.00 million seeds ha
−1 to be the economic optimum for spring wheat in the United Kingdom. Seeding rates above the optimum can potentially result in decreased yield because of increased lodging [
33]. Limited information on the response of hybrids of spring wheat to agronomic inputs is available; however, Lloveras et al. [
34] found a linear relationship between hybrid winter wheat yield and seeding rate up to 5.00 million seed ha
−1. As seeding rate is an important input in wheat production, it is a management practice that can be targeted to reduce production costs by minimizing seed-related inputs while maximizing economic profit. Seed costs for wheat grown in the northern plains typically represent about 13% of the yearly variable input costs [
35].
Wheat grain protein content and quality are largely determined by the genotype which can vary widely in agronomic, quality, and yield characteristics. Previous studies have highlighted the importance of genotype in determining protein and baking quality characteristics [
36,
37,
38]. Agronomic practices, such as seeding rate, can affect yield and quality characteristics. Chen et al. [
8] reported decreased grain protein content as seeding rate increased from 1.08 to 4.30 million seeds ha
−1 in one year of the study. However, other studies noted the absence of significant seeding rate influencing grain protein content [
6,
39,
40,
41]. Controlling or predicting wheat grain protein content is an important consideration for growers as HRSW is typically grown for its high protein content relative to other wheat market classes.
Yield components are usually reported as a representation of the final makeup of yield. These components do not develop independently and there is a complex relationship between the various yield components [
42,
43,
44,
45]. Some components, such as tiller numbers, are more plastic than others, such as kernels per spike [
46]. However, environment and genetics strongly influence yield component composition [
45,
46].
The objective of this research was to determine the AOSR for new spring wheat hybrids, determine the contribution of their various yield components to yield, and to determine if hybrids have increased end-use qualities relative to conventional cultivars.
2. Materials and Methods
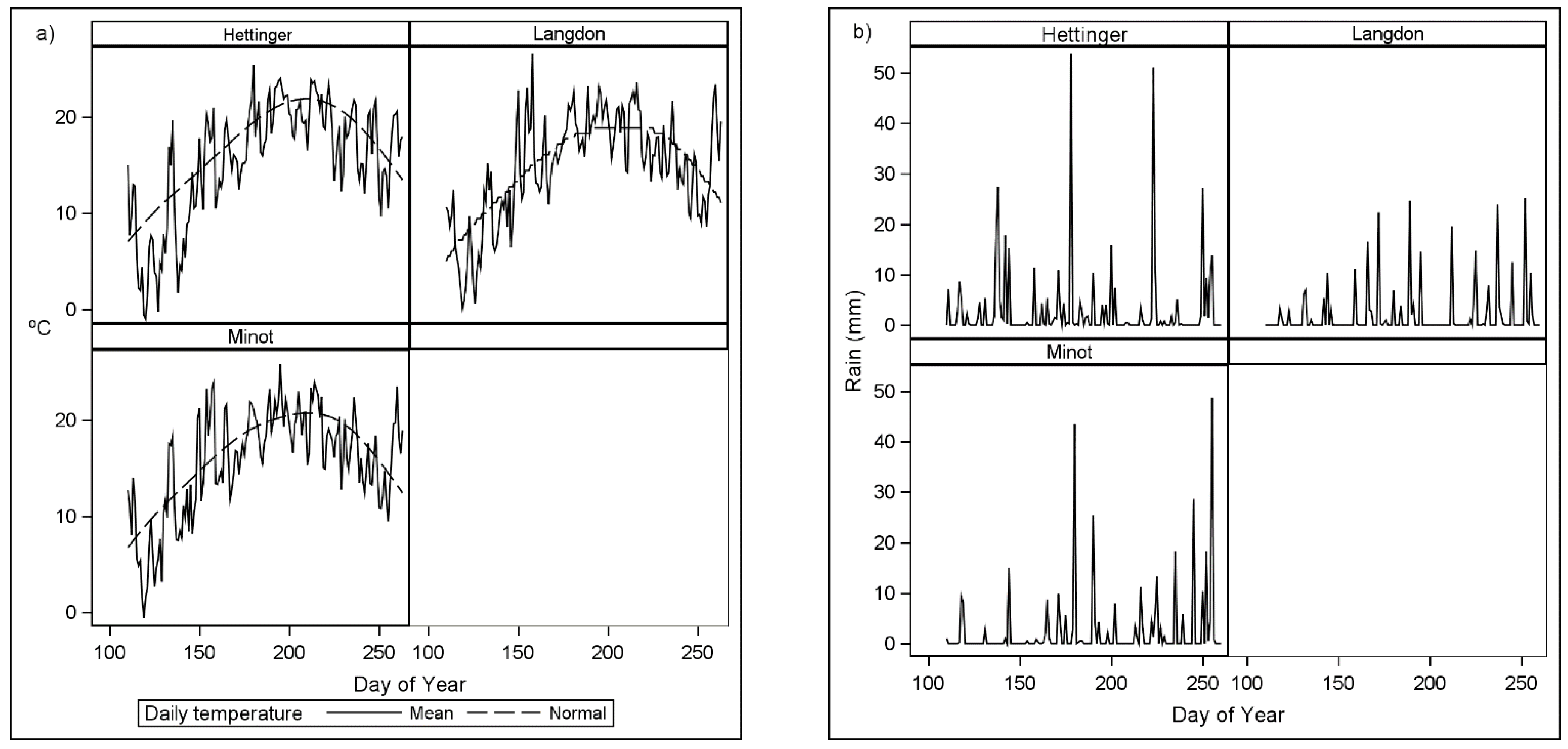
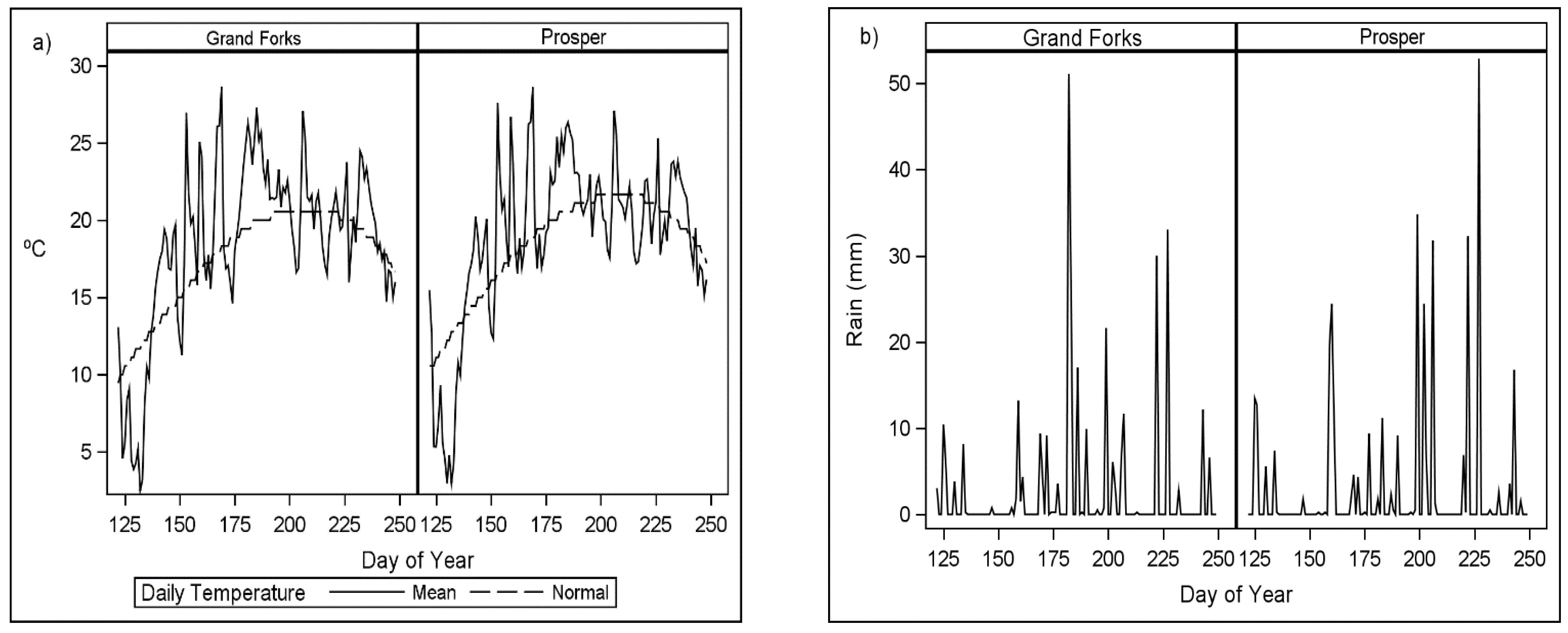
Field experiments were established in three environments in 2019 and two environments in 2020. In 2019, experiments were located in Hettinger, Langdon, and Minot, North Dakota, USA, representing a large geographical area of HRSW production. In 2020, field experiments were conducted in Grand Forks and Prosper, North Dakota, USA.
Table 1 summarizes the physical characteristics of the experimental locations.
Treatments consisted of a factorial combination of seven genotypes (two inbred cultivars and five HRSW hybrids) and seeding rates in a randomized complete block design with a split-plot arrangement with four replications. The whole plot was seeding rate and the sub-plot was genotype. In most environments, there were two planting dates, the first date, which was considered the optimal date, occurred as soon as practical in the spring as recommended by Wiersma and Ransom [
47]. The second date was two weeks thereafter. Seeding rates were 2.22, 2.96, 3.71, 4.45, and 5.19 million live seed ha
−1 which will be referred to using the million seeds ha
−1 unit. The genotypes included were “SY Ingmar” and “SY Valda” (inbred cultivars), which will be referred to as Ingmar and Valda, and five experimental hybrids which will be identified as H
A, H
B, H
C, H
D, and H
E. All genotypes were developed by AgriPro (Syngenta AgriPro, LLC, Greensboro, NC, USA). Ingmar and Valda were among the most commonly grown cultivars and occupied 33% of the total HRSW hectares cultivated in North Dakota in 2019 [
48]. Both the agronomic traits and disease resistance of these cultivars are summarized in (
Table 2). The hybrids included had not yet been commercially released. Their pedigrees were not disclosed and, because of no previous testing, their agronomic characteristics were not known.
The quantity of seed planted for each genotype and seeding rate was calculated on a live seed basis based on a germination test. Plot size, seeding date, and harvest date information are summarized for each location in
Table 3. Management of the trials varied slightly at each location, based on the preferred practices of cooperating researchers at the Hettinger, Langdon, and Minot locations. Soils were tested for plant essential nutrients before seeding to ensure fertility was not a limiting factor (
Table 4) and N, P, and K rates were adjusted to ensure they were not limiting as recommended by Wiersma and Ransom [
47]. Fertilizers were applied before planting if necessary.
The fungicide combination of pydiflumetoen (150 g ai ha
−1) and propiconazole (126 g ai ha
−1) commercially marketed as Miravis Ace (Syngenta Crop Protection, LLC, Greeensboro, NC, USA) was applied to all locations at Zadoks 65 at a rate of 0.164 L ha
−1 to reduce Fusarium head blight incidence (
Fusarium graminearum) and fungal leaf spots. Other management practices were based on North Dakota State University Extension recommendations, regarding cultivation, fertilization, and herbicide and pesticide applications [
47].
Plant density and spike density were obtained by counting plants and spikes in two of the innermost rows of each plot from a 0.91 m length from a stake randomly placed after sowing. Plant density was determined at approximately Zadoks 11 and prior to tiller production. Productive spike density was determined by counting spikes at approximately Zadoks 92 within the same 0.91 m of rows used for plant density measurements. Small spikes that were deemed not to contribute to yield were not counted.
Spikes plant−1 and kernels spike−1 were derived from plant density, spike count, and kernel weight measurements. Yield was collected for each plot using a small plot combine and was adjusted to 13.5% moisture. Moisture and test weight were determined using a GAC 2100 moisture tester (Dickey-John Corp., Minneapolis, MN, USA). Percent grain protein content was measured using a DA 7250 NIR analyzer (Perten Instruments, Stockholm, Sweden) and was reported on a 12% moisture basis. The moisture contents used for reporting yield and protein content were those commonly used by the wheat industry in the USA. To report protein yield, protein and yield, which are commonly reported at 12 and 13.5% moisture, respectively, were corrected to 12% moisture and multiplied.
An ANOVA was performed on each measured variable using the MIXED procedure in SAS 9.4 (SAS Institute, Cary, NC, USA). Environments with the ratio of the highest and lowest error mean square for yield, protein, or protein yield being less than 10 were combined for analysis [
49]. In the combined analysis, location-year were considered an environment and a random effect while seeding rate, genotype, and the seeding rate by genotype interaction were considered fixed effects and analyzed as described by Carmer [
50]. Analysis of variance was conducted for yield (seven environments), protein (eight environments), and protein yield (eight environments). Significant ANOVA seeding rate by genotype interactions for yield and protein yield were analyzed using the MIXED procedure with treatment means separated by Tukey’s multiple comparison test at P = 0.05. The seeding rate by genotype interaction term was further analyzed using regression analysis and the REG procedure in SAS.
The wheat quality analysis was performed on the grain of each genotype from the 2.96 million seeds ha
−1 seeding rate treatment. The 2.96 million seeds ha
−1 rate was chosen to represent a typical seeding rate that farmers use in the area. The harvested samples from replicates one and two and from replicates three and four were combined for each of these treatments from each location to form two replicates for each genotype and environment. Grain samples were analyzed by the North Dakota State University Wheat Quality Laboratory for test weight, kernel size distribution, kernel weight, protein, falling number, milling extraction, mixograph score, peak maximum time, maximum torque, total energy, and loaf volumes, according to standard protocols [
51].
Planting dates within environments had differing yield responses, so data from each planting date were partitioned into high (>5000 kg ha
−1) and low (<5000 kg ha
−1) yielding environment datasets by considering individual planting dates as a single environment for a total of nine environments similar to Mehring et al. [
52]. Yield within an environment was evaluated by standardizing the distribution of each, using z-scores. Data were transformed using the Standard procedure in SAS to calculate a z-score for yield using the formula
where x is yield,
is the yield mean of the high or low yielding environment, and
is the standard deviation of the high or low yield environment [
53]. The z-score adjusts the data distribution to have a mean of 0 and a standard deviation of 1. Next, the z-score data were used to adjust yield values to be relative to the yield of the individual environment using the formula Ŷ = (z-score ×
) + μ where Ŷ is estimated yield, z-score is z-score of yield,
is the standard deviation of the individual environment, and μ is the mean of the high or low yield environment.
Relationships between yield components and yield for high and low yielding environments were analyzed using yields adjusted by the z-score approach previously discussed. Multiple linear regression was used to determine the relative importance of each yield component on yield using the REG procedure which also derived partial correlation coefficients (r2) and adjusted R2. Wheat end quality characteristics were analyzed using the GLM procedure in SAS with single degree of freedom linear contrasts for cultivar and hybrid comparison evaluated at α = 0.05.






{kind=link}
{kind=link}