Author Contributions
Conceptualization, A.A.M.A. and M.M.R.; data curation, A.A.M.A., M.M.R., A.M. and E.F.A.; formal analysis: A.A.M.A., A.A.A.S., A.M. and E.F.A.; investigation, A.A.M.A., A.A.A.S. and E.F.A.; methodology, A.A.M.A., A.A.A.S., M.M.R. and E.F.A.; resources, A.A.M.A., A.A.A.S., M.M.R., A.M. and E.F.A.; software, A.A.M.A., A.A.A.S., M.M.R., A.M. and E.F.A.; writing—original draft, A.A.M.A., A.A.A.S., M.M.R., A.M. and E.F.A.; and writing—review and editing, A.A.M.A., M.M.R., A.M. and E.F.A. All authors have read and agreed to the published version of the manuscript.
Figure 1.
TEM image of zinc oxide nanoparticles (ZnONPs).
Figure 1.
TEM image of zinc oxide nanoparticles (ZnONPs).
Figure 2.
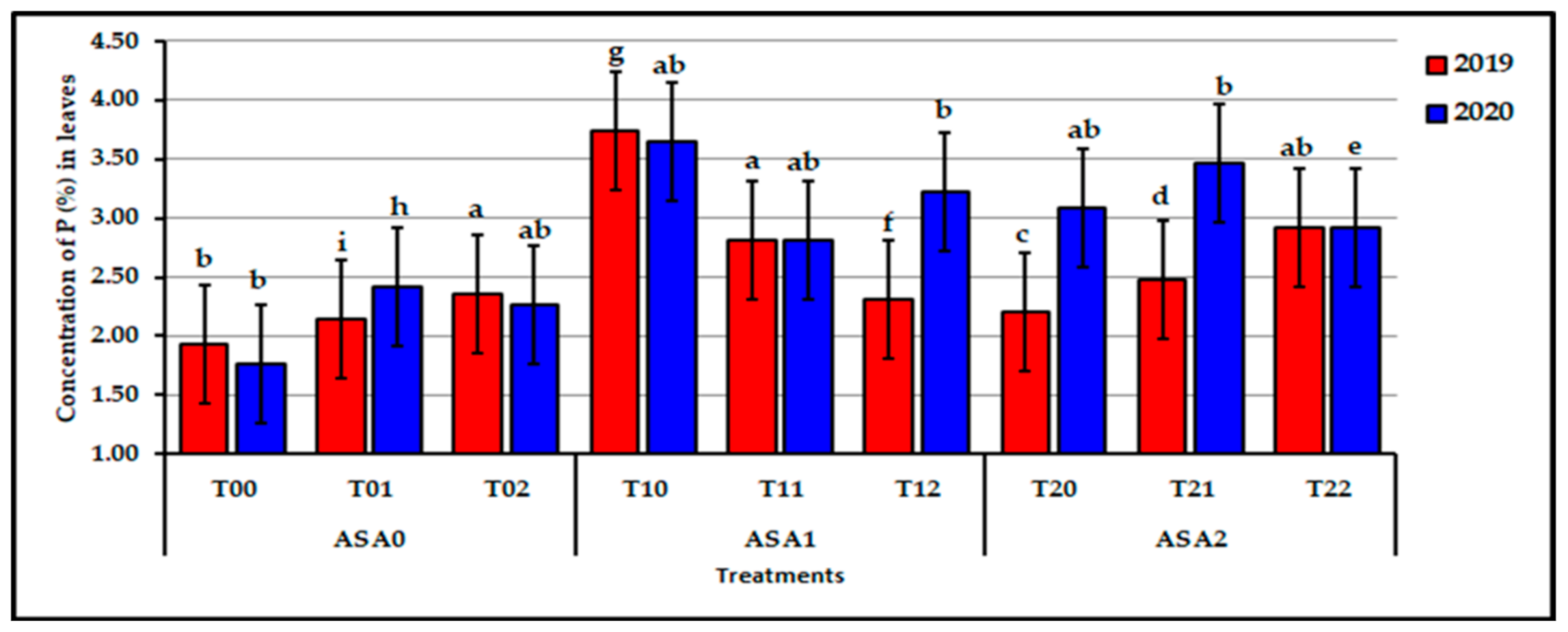
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf P content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 2.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf P content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 3.
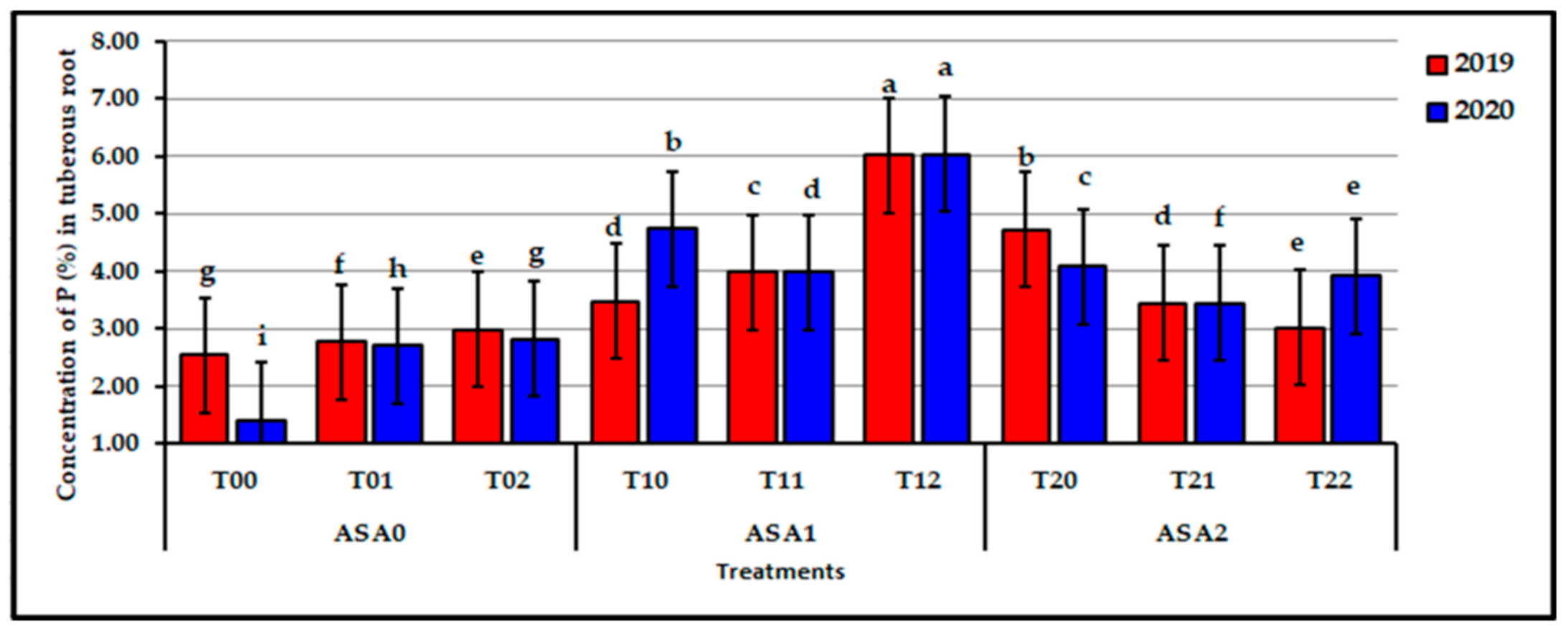
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root P content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 3.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root P content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 4.
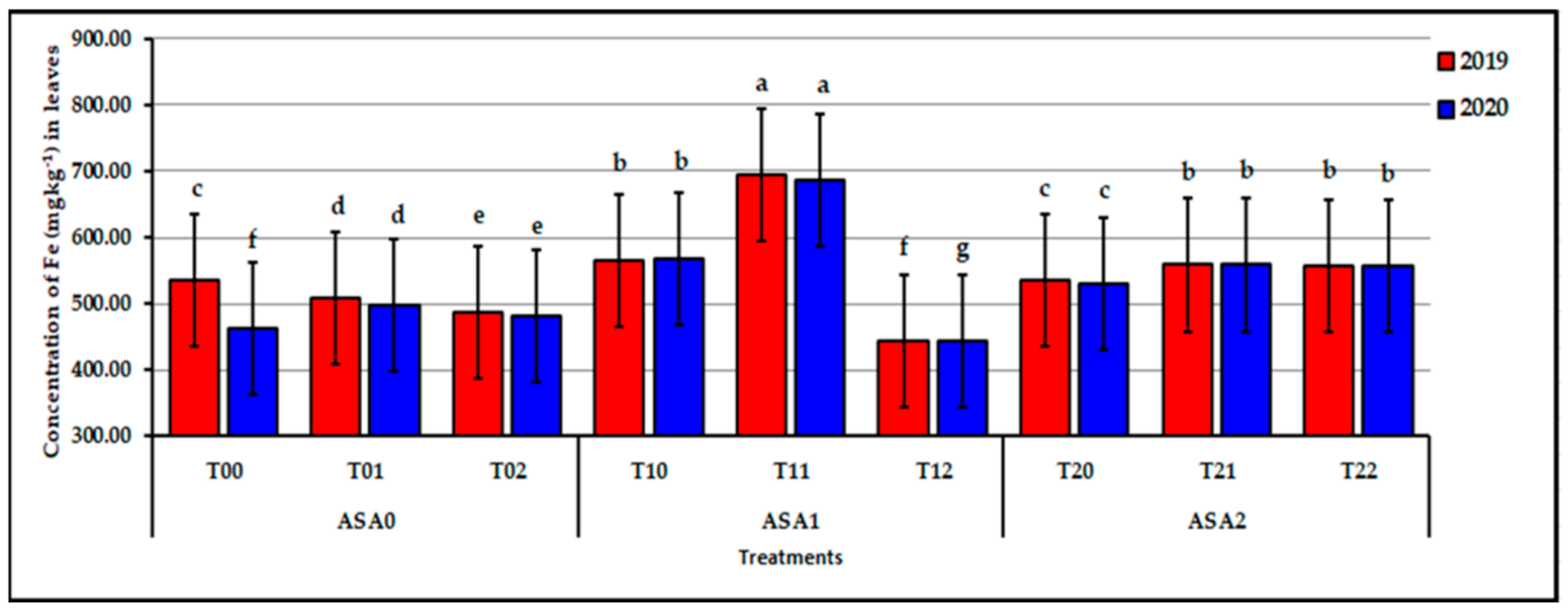
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Fe content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 4.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Fe content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 5.
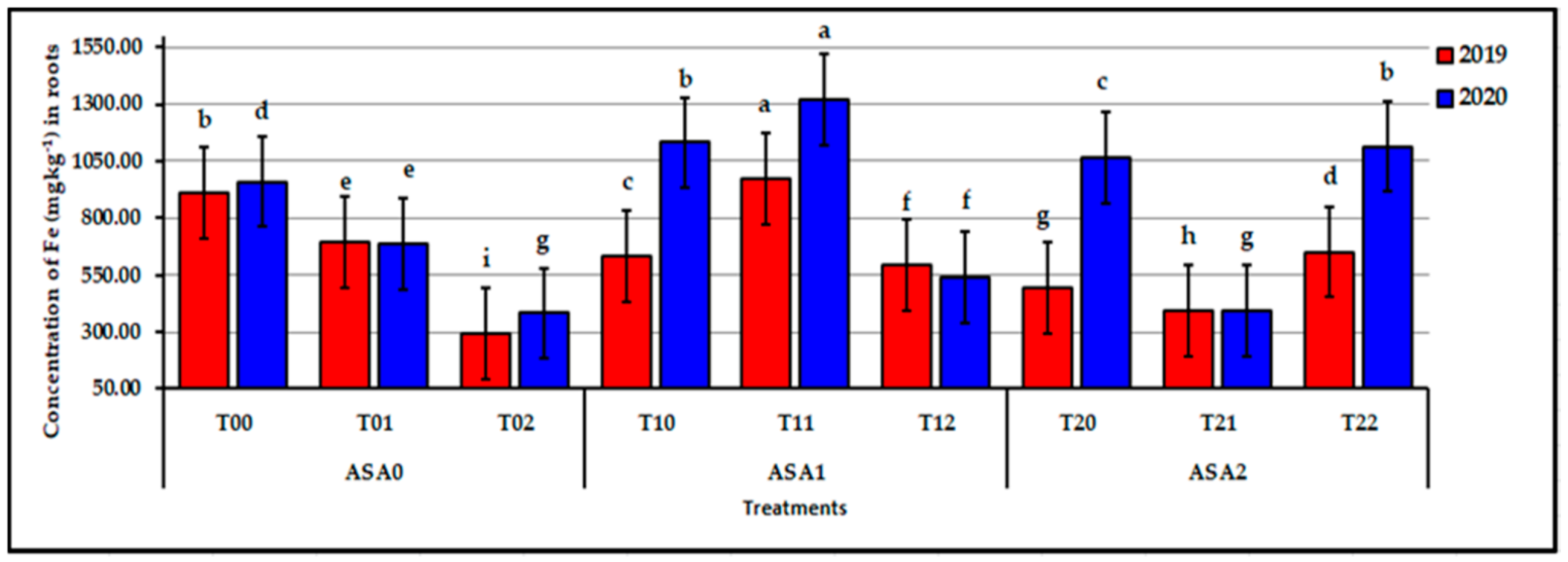
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Fe content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 5.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Fe content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
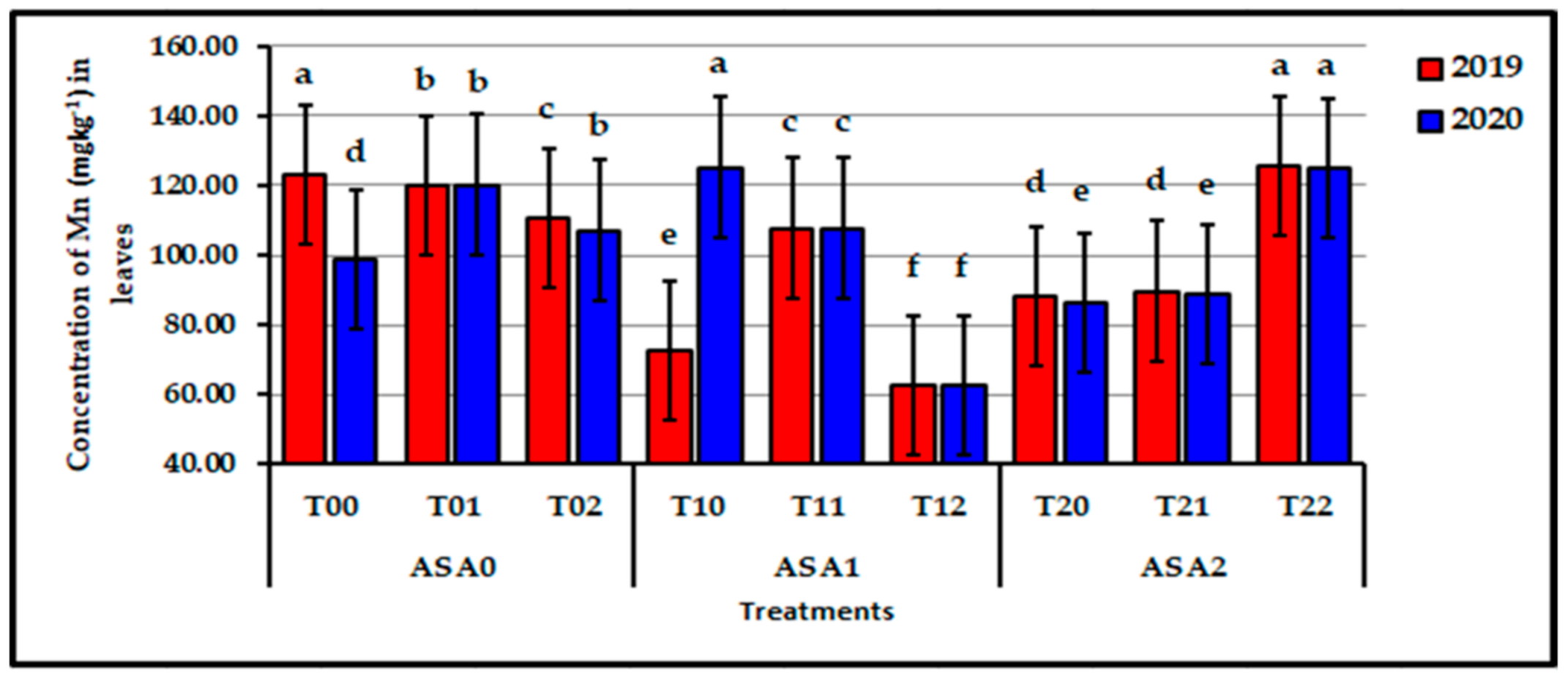
Figure 6.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Mn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 6.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Mn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
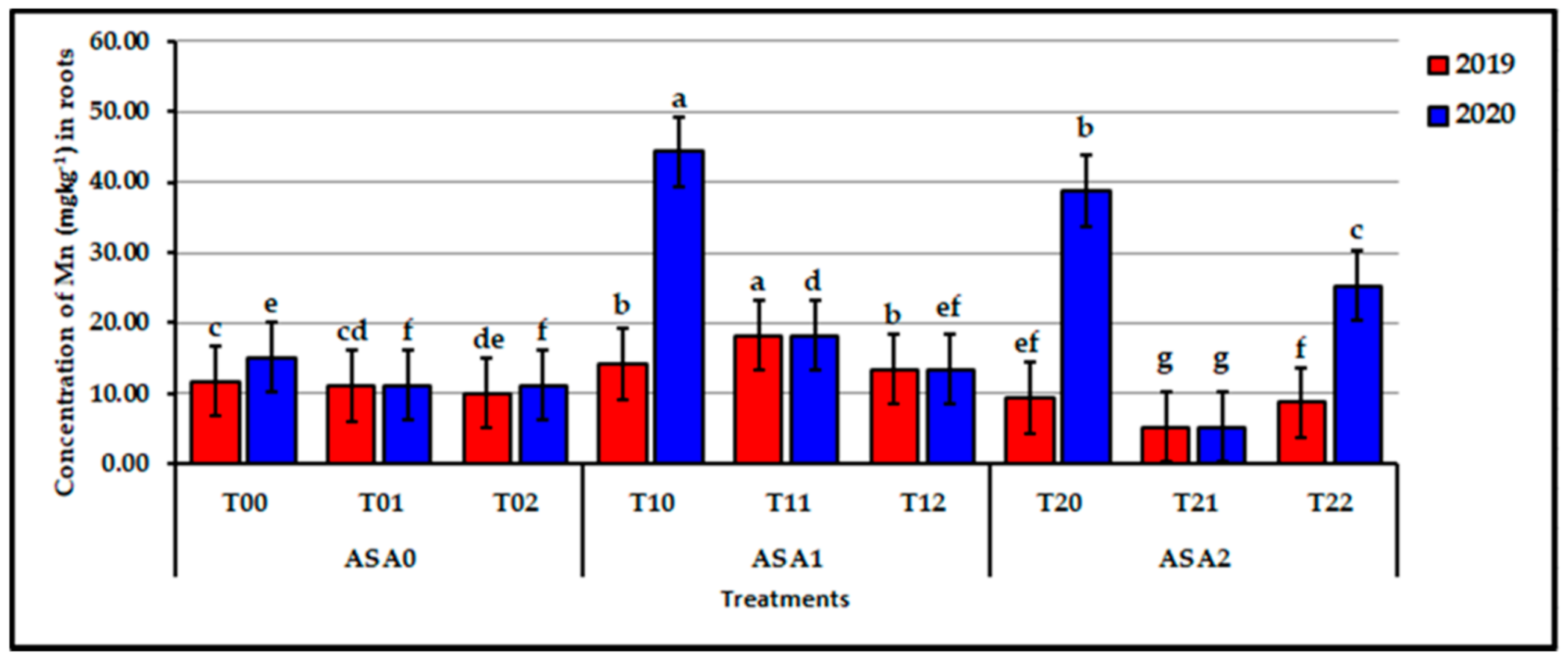
Figure 7.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Mn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 7.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Mn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
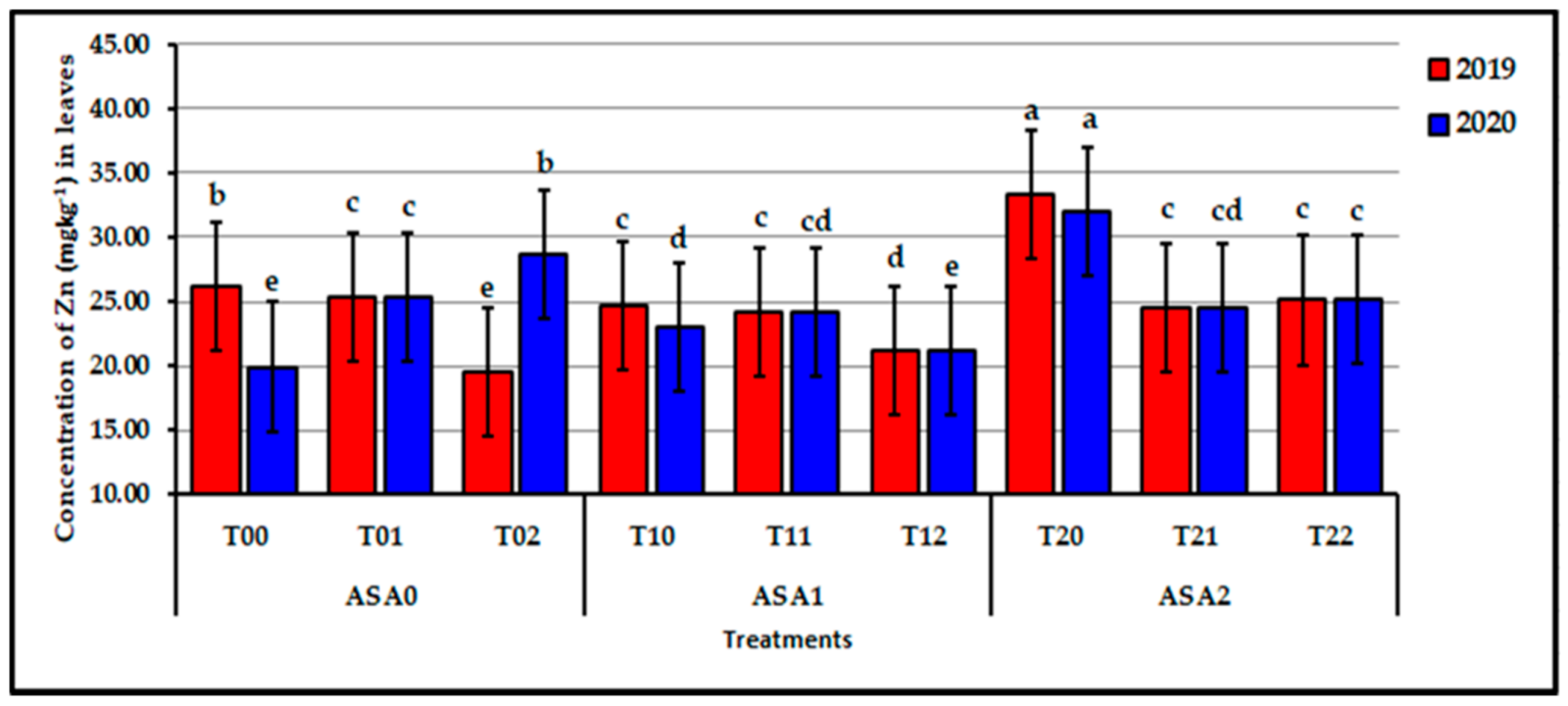
Figure 8.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Zn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 8.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Zn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
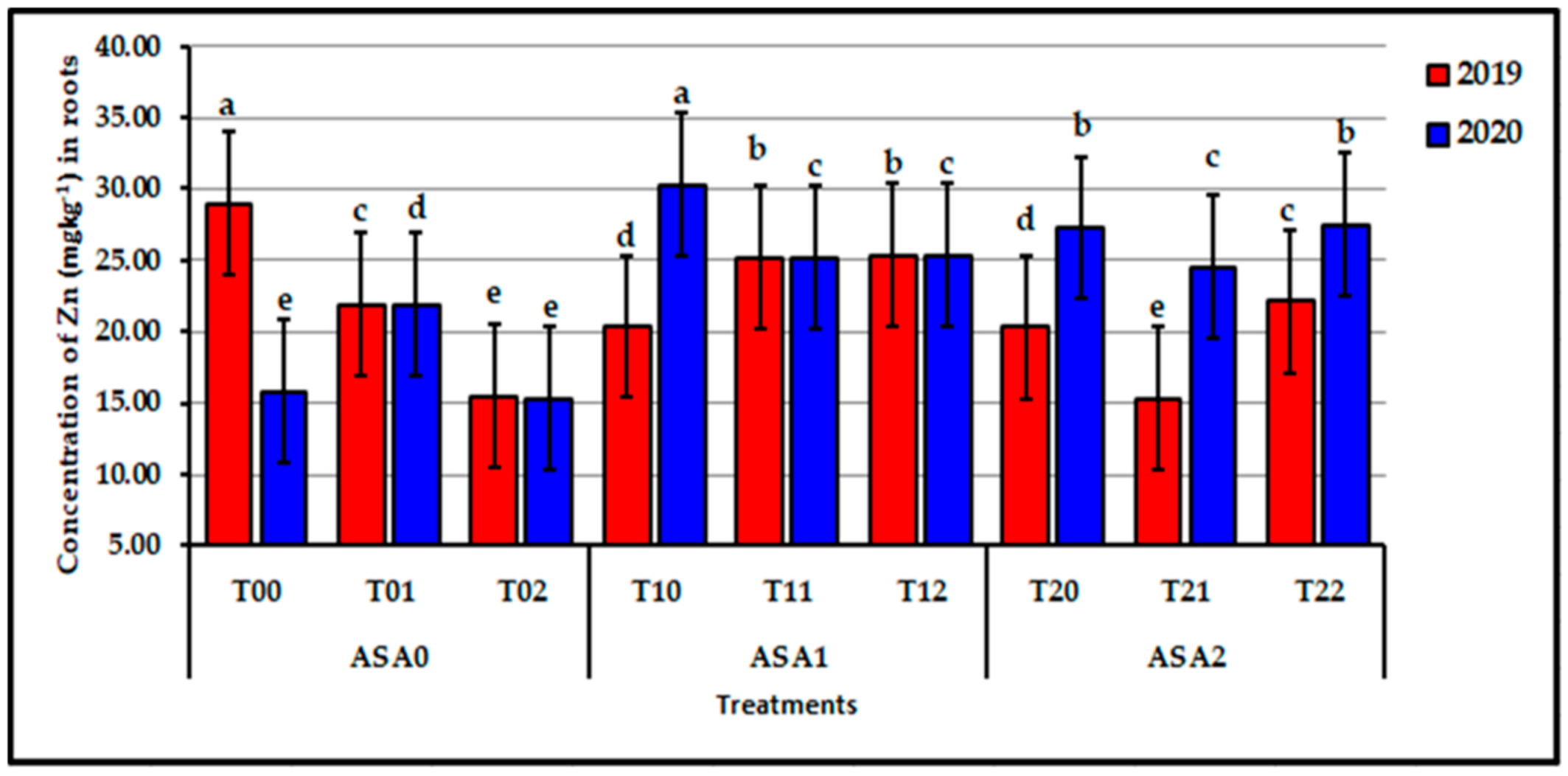
Figure 9.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Zn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 9.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Zn content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
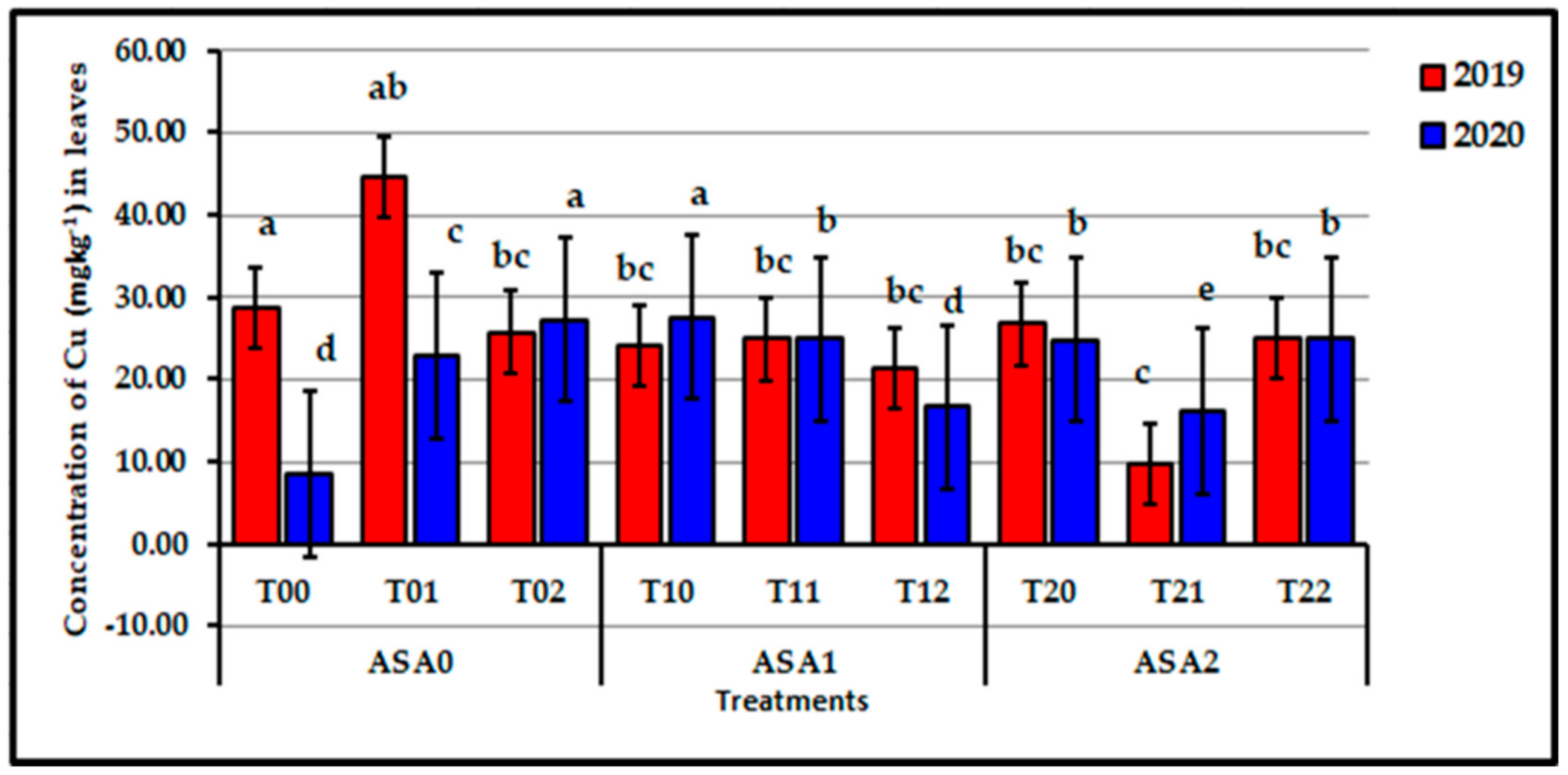
Figure 10.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Cu content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 10.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on leaf Cu content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
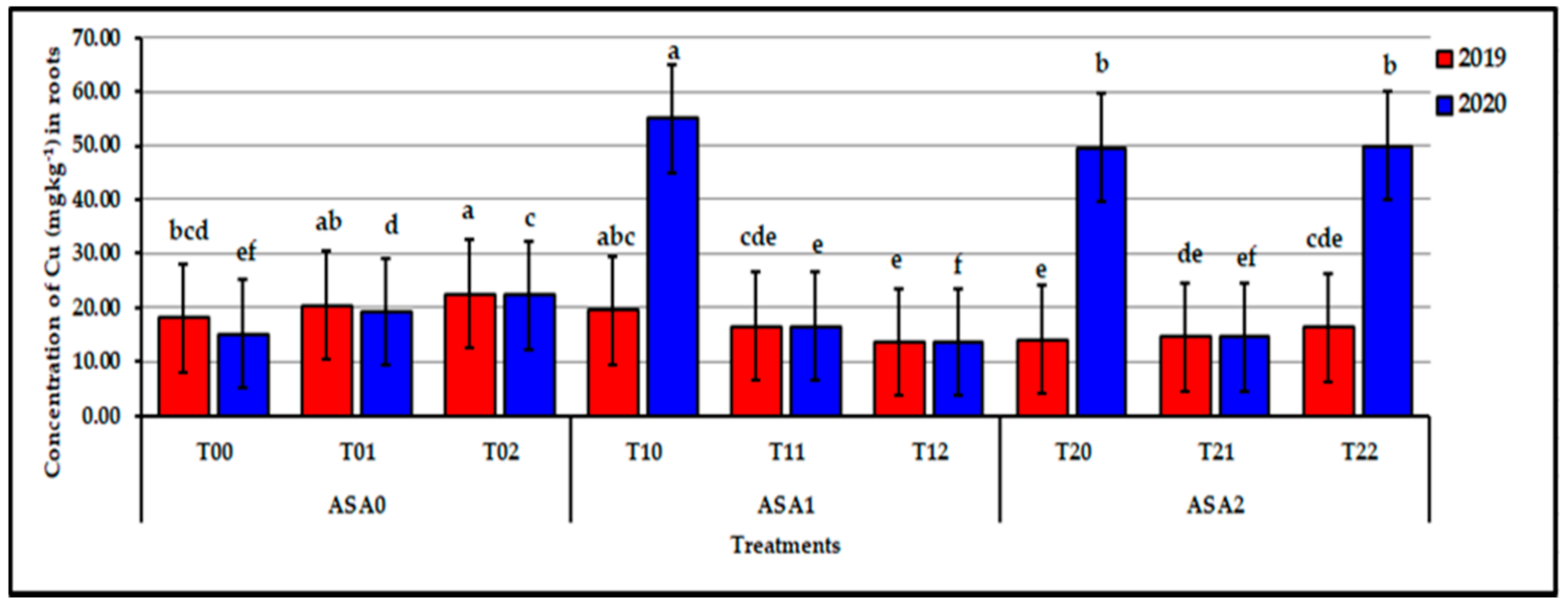
Figure 11.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Cu content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 11.
Influence of interaction between zinc oxide nanoparticles and ascorbic acid on root Cu content of stressed sweet potato plants in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
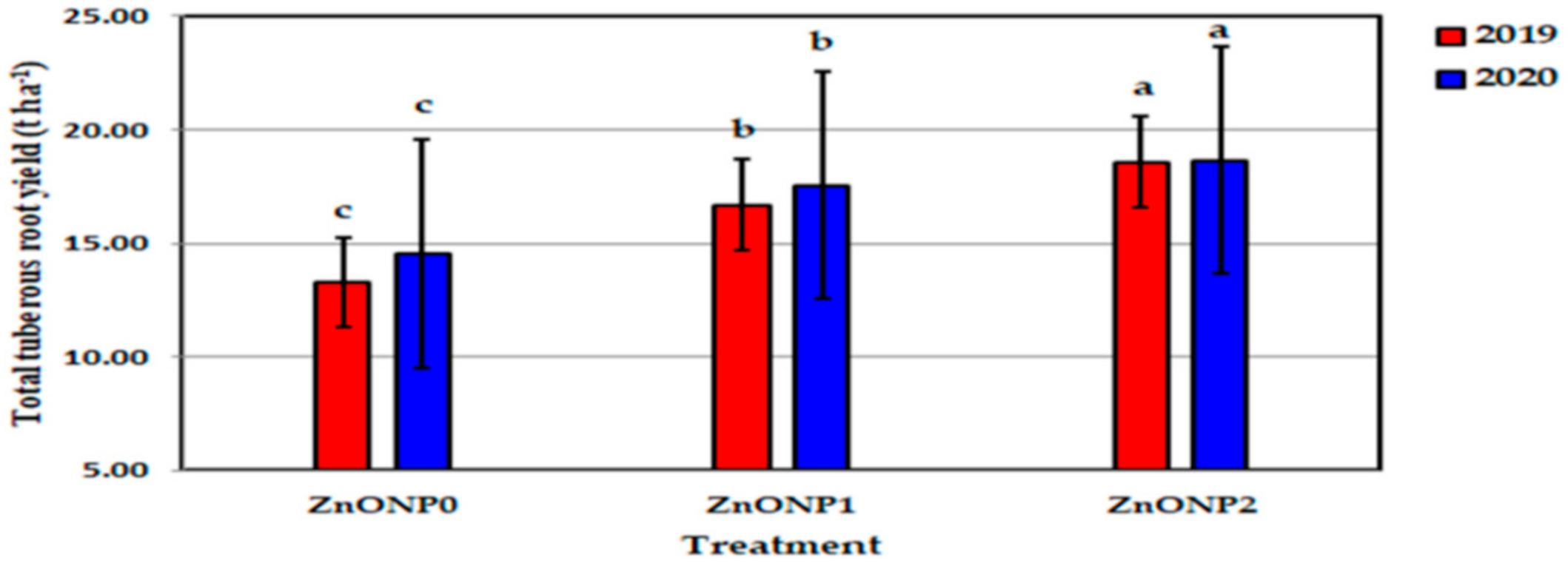
Figure 12.
Influence of zinc oxide nanoparticles doses on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 12.
Influence of zinc oxide nanoparticles doses on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 13.
Influence of ascorbic acid doses on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 13.
Influence of ascorbic acid doses on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
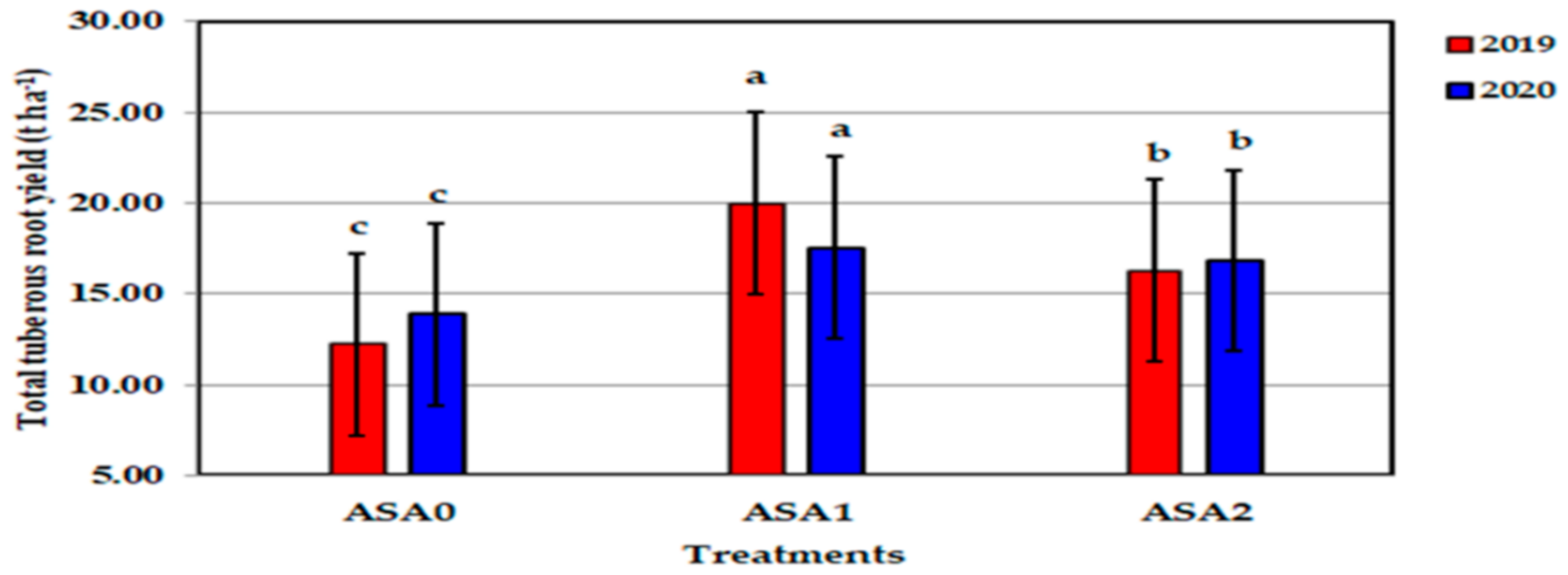
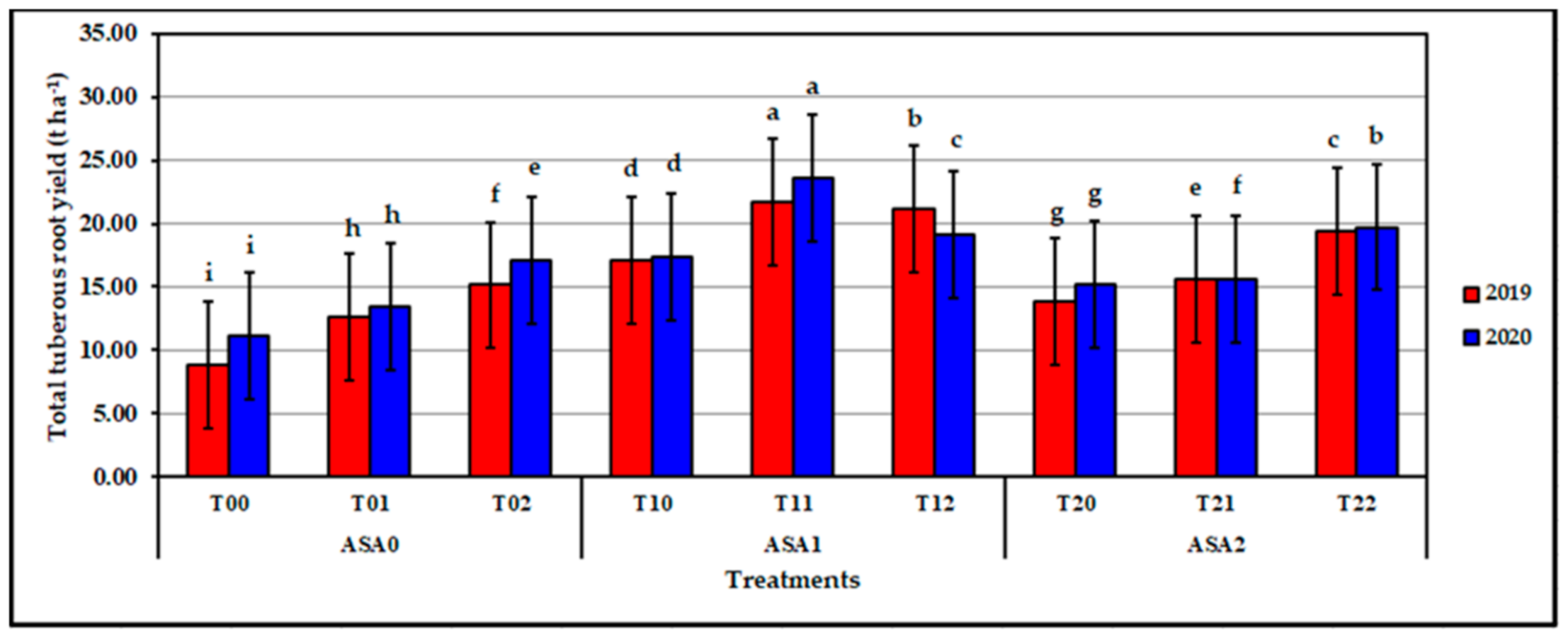
Figure 14.
Influence of ascorbic acid and zinc oxide nanoparticles interaction on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
Figure 14.
Influence of ascorbic acid and zinc oxide nanoparticles interaction on total tuberous roots yield of sweet potato in 2019 and 2020 seasons. Bars with a different letter indicate significant difference between treatments at p ≤ 0.05.
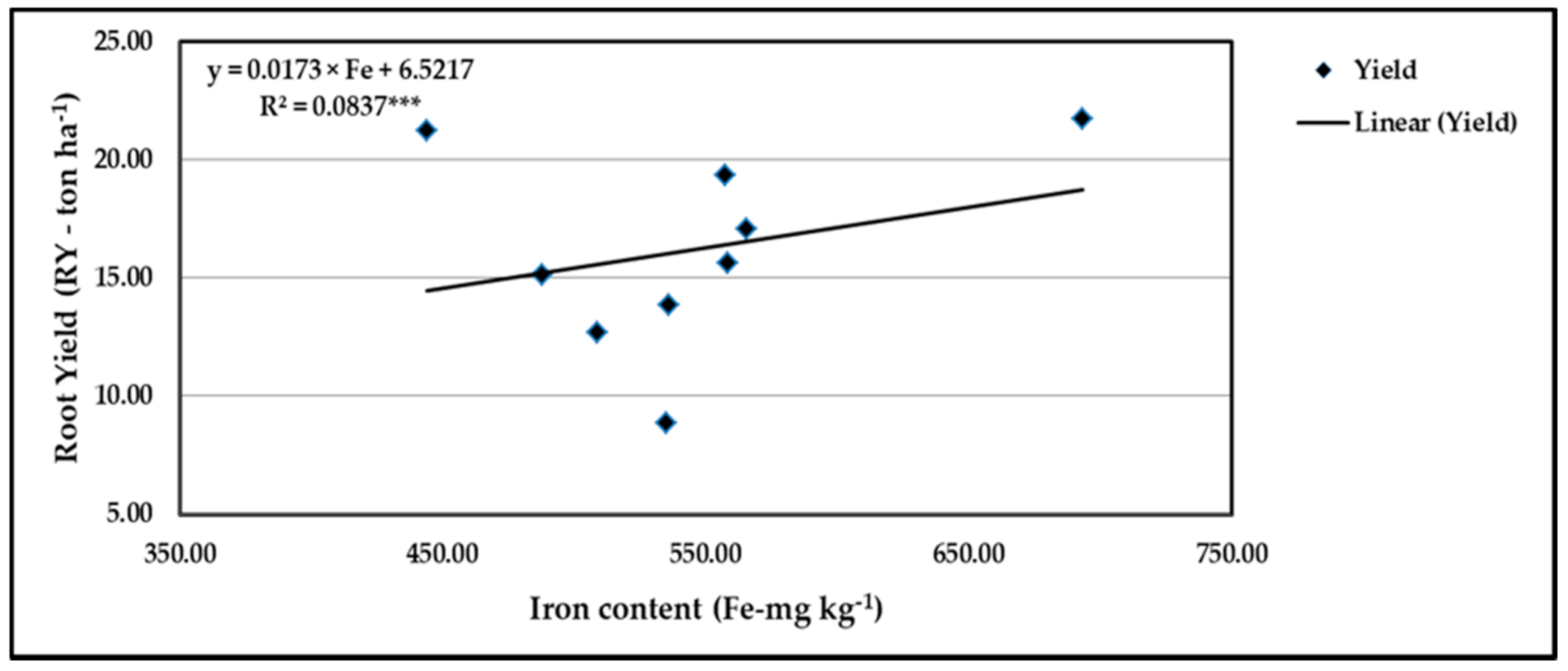
Figure 15.
Contribution of Fe content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 15.
Contribution of Fe content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
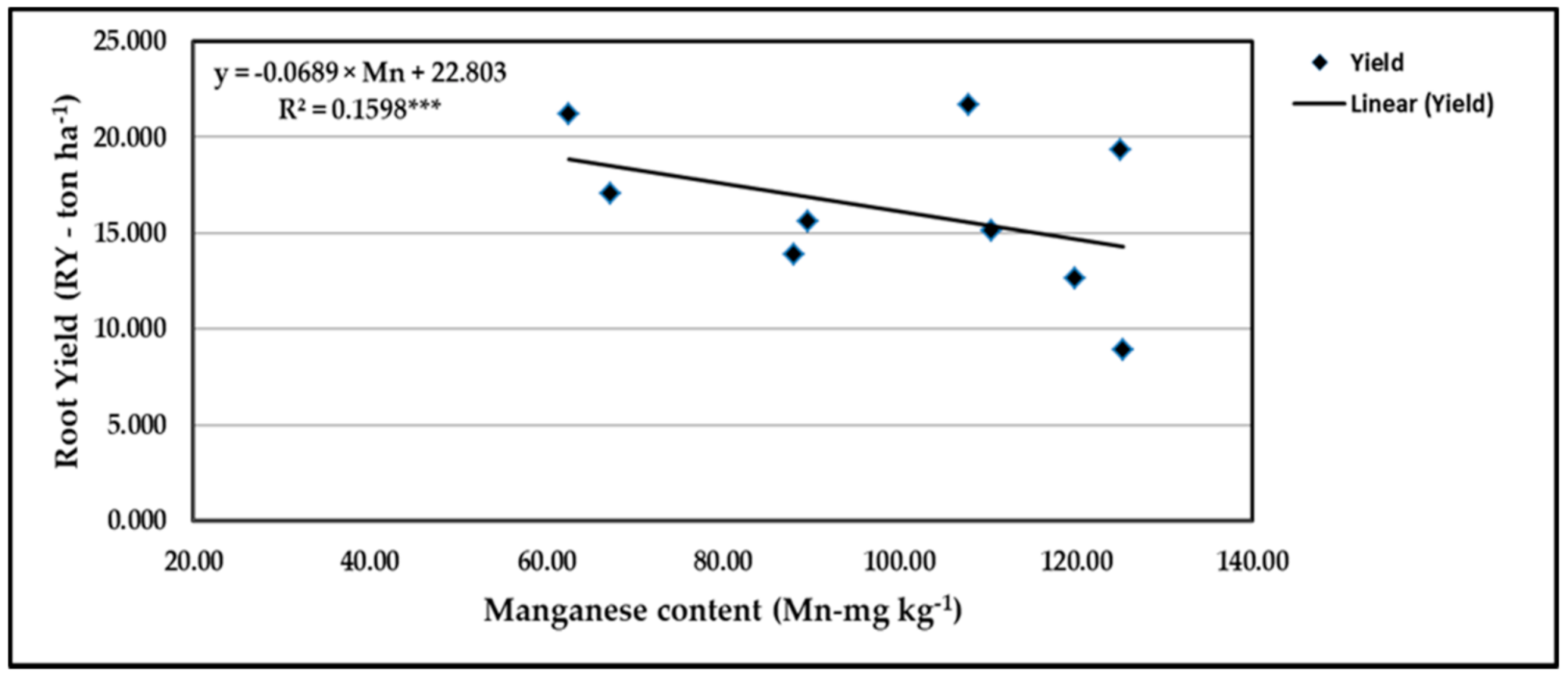
Figure 16.
Contribution of Mn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 16.
Contribution of Mn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
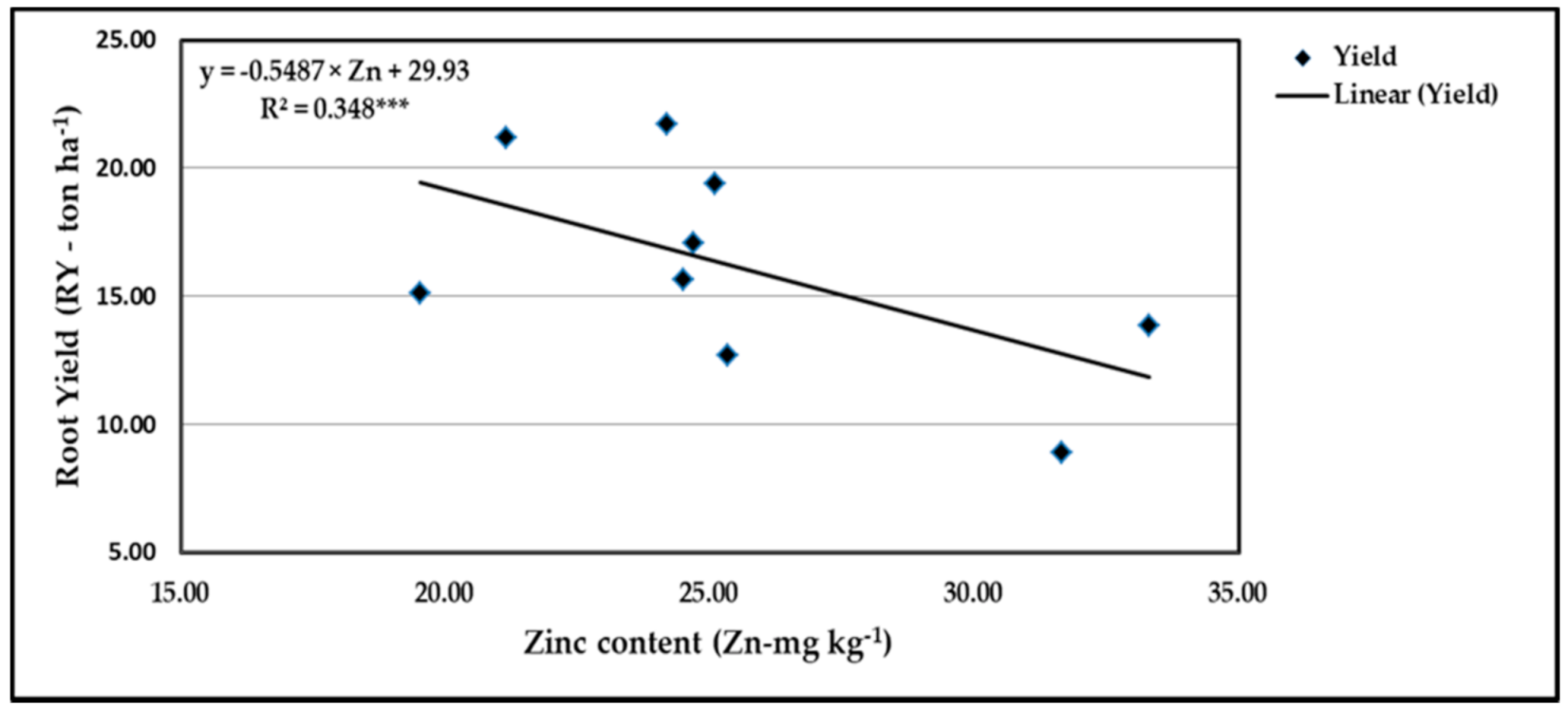
Figure 17.
Contribution of Zn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 17.
Contribution of Zn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2019 season. *** indicates differences at p ≤ 0.001 probability level.
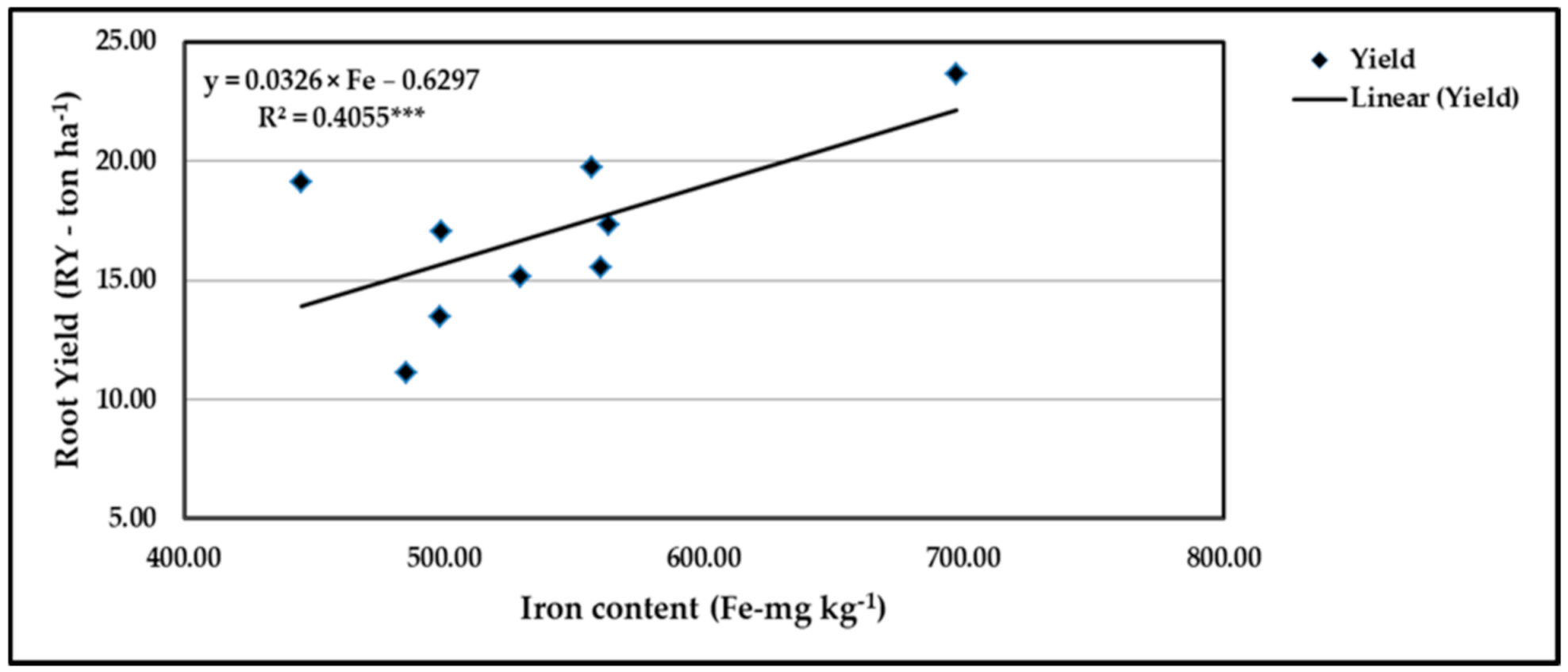
Figure 18.
Contribution of Fe content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 18.
Contribution of Fe content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
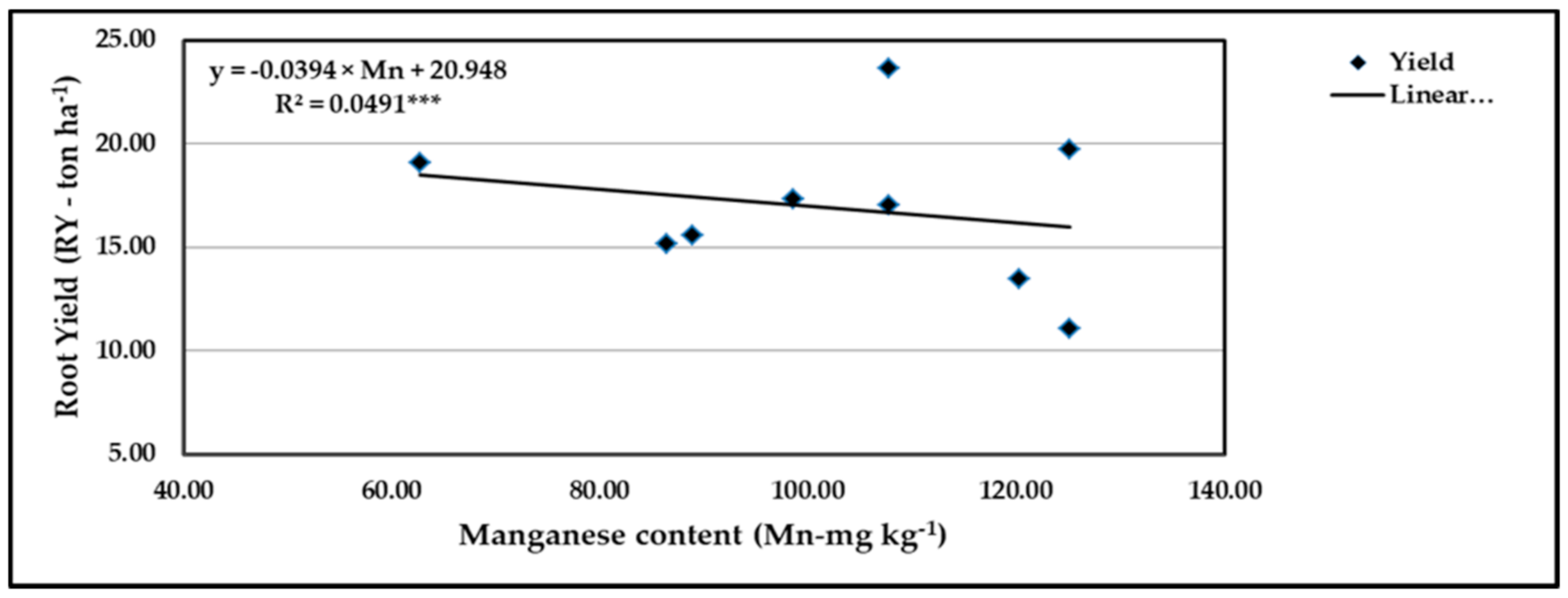
Figure 19.
Contribution of Mn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 19.
Contribution of Mn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
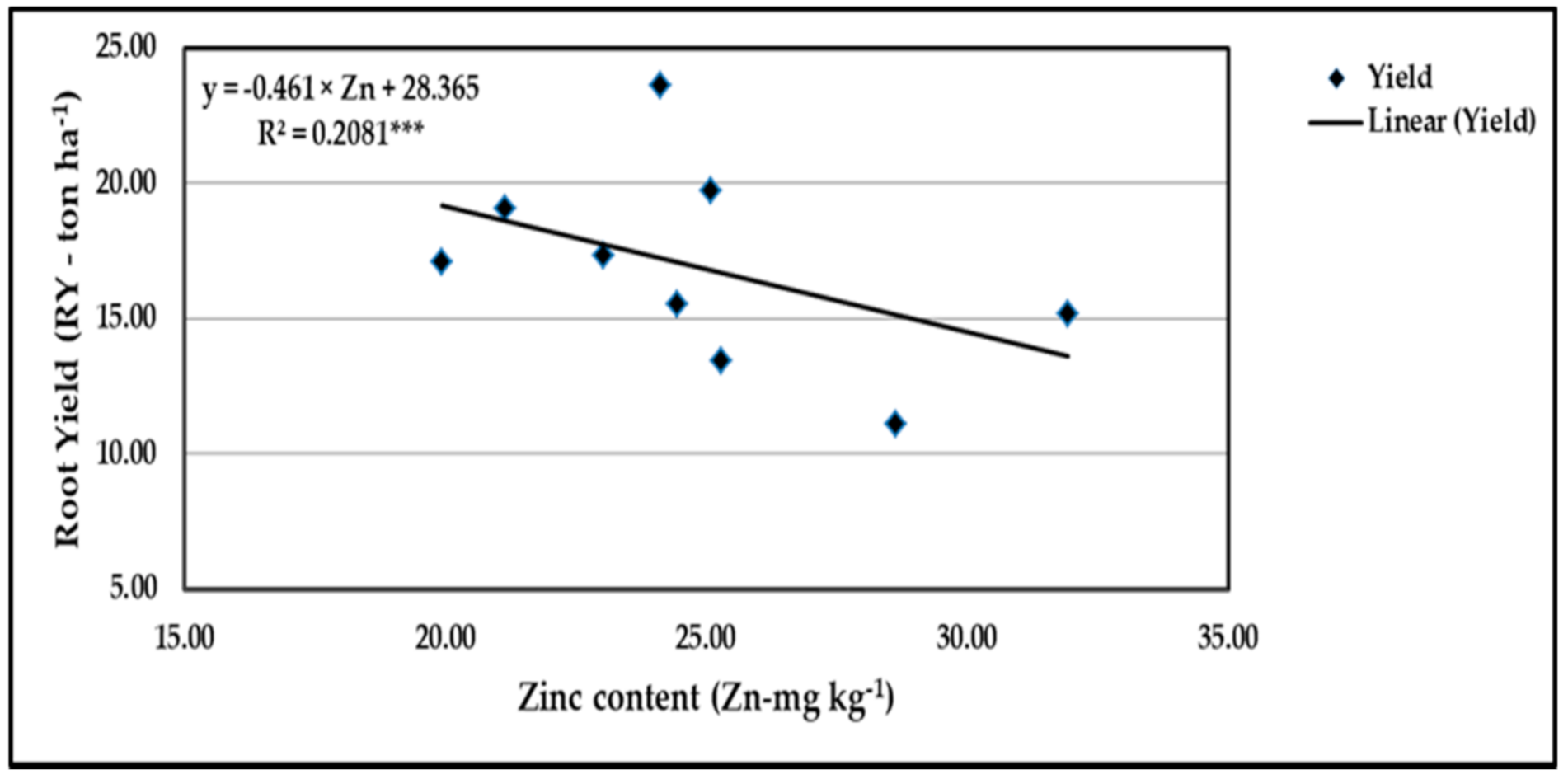
Figure 20.
Contribution of Zn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 20.
Contribution of Zn content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
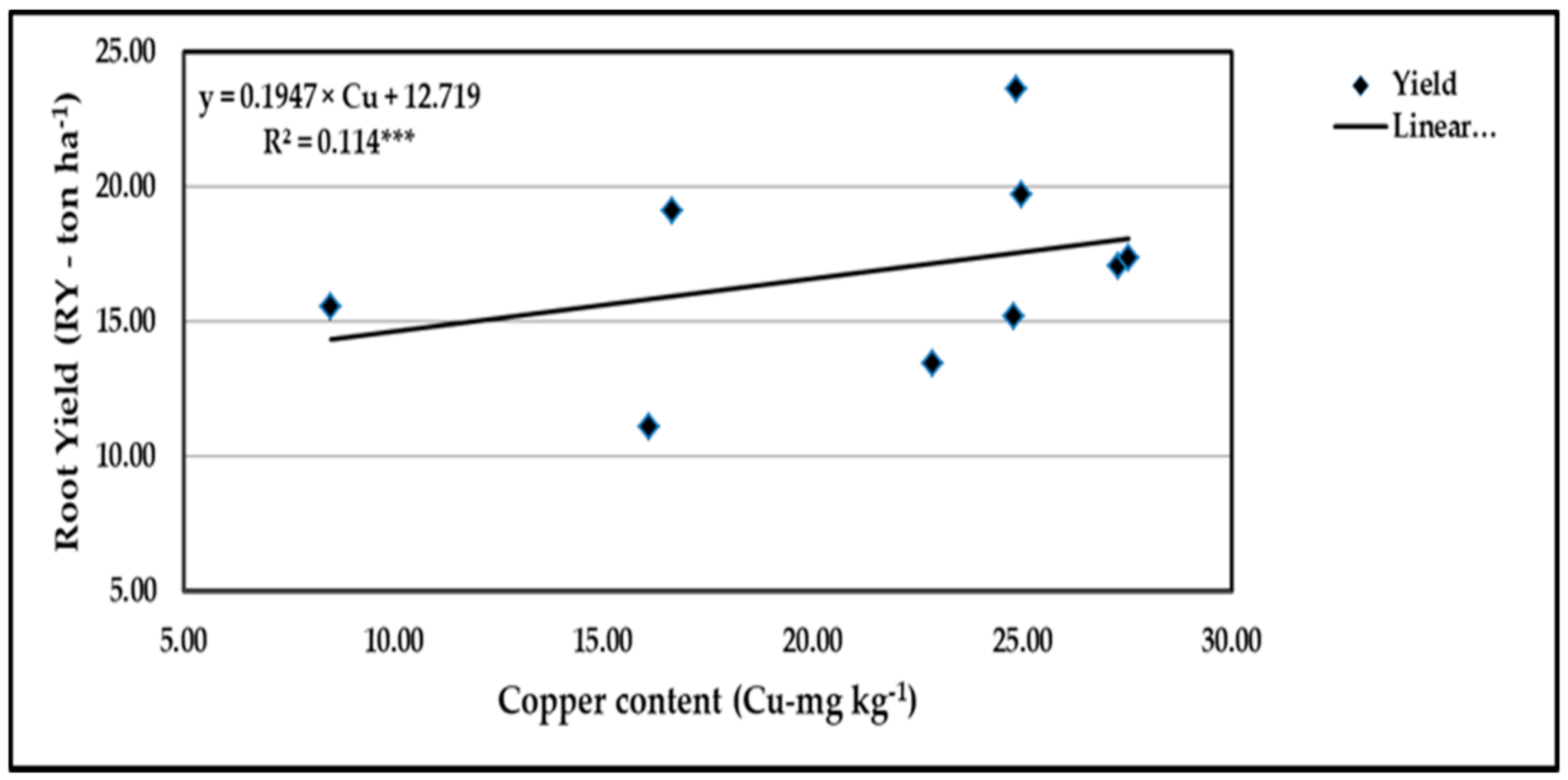
Figure 21.
Contribution of Cu content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
Figure 21.
Contribution of Cu content to the quadratic response of stressed sweet potato to different levels of ascorbic acid and zinc oxide nanoparticles in the 2020 season. *** indicates differences at p ≤ 0.001 probability level.
Table 1.
Average weather data for the Fayoum region, Egypt, over the sweet potato growing seasons.
Table 1.
Average weather data for the Fayoum region, Egypt, over the sweet potato growing seasons.
| Month | Average Day Temperature (°C) | Average Night Temperature (°C) | Average Relative Humidity (%) | Average Speed of Wind (ms−1) | Average of Measured Pan Evaporation Class A (mm d−1) | Average Precipitation (mm d−1) |
|---|
| April | 32.9 | 16.1 | 32.3 | 1.87 | 5.58 | 0.02 |
| May | 35.8 | 18.9 | 34.1 | 1.88 | 6.87 | 0.00 |
| June | 40.0 | 19.8 | 38.4 | 1.49 | 7.56 | 0.00 |
| July | 41.1 | 24.8 | 37.9 | 2.03 | 6.88 | 0.02 |
| August | 41.3 | 25.6 | 36.8 | 1.78 | 6.78 | 0.00 |
Table 2.
The studied experimental treatments.
Table 2.
The studied experimental treatments.
| Symbol | Treatment | Ascorbic Acid | Zinc Oxide Nanoparticles |
|---|
| T00 | ASA0, ZnONP0 | No ascorbic acid was applied | No zinc oxide nanoparticles was applied |
| T01 | ASA0,ZnONP1 | 1000 mg L−1 of zinc oxide nanoparticles |
| T02 | ASA0,ZnONP2 | 1500 mg L−1 of zinc oxide nanoparticles |
| T10 | ASA1,ZnONP0 | 250 mg L−1 of ascorbic acid | No zinc oxide nanoparticles was applied |
| T11 | ASA1,ZnONP1 | 1000 mg L−1 of zinc oxide nanoparticles |
| T12 | ASA1,ZnONP2 | 1500 mg L−1 of zinc oxide nanoparticles |
| T20 | ASA2,ZnONP0 | 500 mg L−1 of ascorbic acid | No zinc oxide nanoparticles was applied |
| T21 | ASA2,ZnONP1 | 1000 mg L−1 of zinc oxide nanoparticles |
| T22 | ASA2,ZnONP2 | 1500 mg L−1 of zinc oxide nanoparticles |
Table 3.
Some soil physical and chemical characters.
Table 3.
Some soil physical and chemical characters.
| Soil Property | 2019 | 2020 |
|---|
| Particle size distribution | | |
| Sand % | 66.8% | 63.6 |
| Silt % | 16.5% | 7.8 |
| Clay % | 16.7% | 28.6 |
| Soil texture | Sandy clay loam | Sandy loam |
| pH in soil paste | 7.77 | 7.19 |
| ECe (dSm−1) in soil paste extracted | 4.24 | 3.95 |
| Organic matter (OM %) | 1.03 | 0.90 |
| CaCO3 (%) | 11.3 | 10.8 |
| Soluble ions (mmol L−1) | | |
| CO3− | --- | --- |
| HCO3− | 2.70 | 2.03 |
| Cl− | 25.6 | 21.1 |
| SO4− | 18.3 | 20.3 |
| Na+ | 31.3 | 31.6 |
| K+ | 0.88 | 0.65 |
| Ca++ | 7.47 | 7.11 |
| Mg++ | 6.98 | 4.03 |
| Total N (mg kg−1) | 515 | 450 |
| Available-P mgkg−1 (Extractable with NaHCO3, pH = 8.5) | 4013 | 3424 |
| Available-K mgkg−1 (Extractable with NH4AOC pH = 7.0) | 1237 | 1816 |
| Fe | (mg kg−1) (Extractable with DPTA) | 4.15 | 6.03 |
| Mn | 10.7 | 18.2 |
| Zn | 0.04 | 0.07 |
| Cu | 0.40 | 0.68 |
Table 4.
Influence of zinc oxide nanoparticles on leaves and tuberous root nutrients content of stressed sweet potato in 2019 and 2020 seasons.
Table 4.
Influence of zinc oxide nanoparticles on leaves and tuberous root nutrients content of stressed sweet potato in 2019 and 2020 seasons.
| Treatment | Parameters |
|---|
| Leaves | Tuberous Roots |
|---|
| ZnONPs | P | Fe | Mn | Zn | Cu | P | Fe | Mn | Zn | Cu |
|---|
| (%) | (mg kg−1) | (%) | (mg kg−1) |
|---|
| | 2019 growth season |
| ZnONP0 | 2.6a | 543.4b | 94.6c | 28.1a | 26.5a | 3.5b | 677.6b | 11.80a | 23.2a | 17.2a |
| ZnONP1 | 2.5b | 586.9a | 105.8a | 24.7b | 26.5a | 3.4c | 684.9b | 11.54a | 20.8b | 17.2a |
| ZnONP2 | 2.5b | 496.4c | 99.3b | 21.9c | 24.0a | 3.7a | 511.8a | 10.79b | 21.0b | 17.5a |
| LSD0.05 | 0.09 | 9.3 | 1.8 | 1.7 | ns | 0.07 | 15.7 | 0.65 | 0.25 | ns |
| | 2020 growth season |
| ZnONP0 | 2.9a | 520.2b | 103.4a | 25.0a | 20.3b | 3.4b | 999.8a | 32.76a | 24.45a | 39.9a |
| ZnONP1 | 2.9a | 583.4a | 105.7a | 24.6a | 21.3b | 3.4c | 800.2b | 11.58c | 23.88a | 16.8a |
| ZnONP2 | 2.8a | 494.5c | 98.5b | 25.0a | 23.0a | 4.3a | 679.2c | 23.11b | 22.71b | 28.6a |
| LSD0.05 | ns | 8.8 | 2.7 | ns | 1.21 | 0.05 | 23.8 | 2.24 | 0.78 | ns |
Table 5.
Influence of ascorbic acid on leaves and tuberous root nutrients content of stressed sweet potato in 2019 and 2020 seasons.
Table 5.
Influence of ascorbic acid on leaves and tuberous root nutrients content of stressed sweet potato in 2019 and 2020 seasons.
| Treatment | Parameters |
|---|
| Leaves | Tuberous Roots |
|---|
| ASA | P | Fe | Mn | Zn | Cu | P | Fe | Mn | Zn | Cu |
|---|
| (%) | mg kg−1 | (%) | mg kg−1 |
|---|
| | 2019 season |
| ASA0 | 2.1a | 510.5b | 117.8a | 23.7b | 33.1a | 2.8c | 631.7b | 11.0b | 22.1b | 20.3a |
| ASA1 | 3.0a | 567.6a | 81.0c | 23.4b | 23.4a | 4.2a | 730.9a | 15.3a | 23.6a | 16.6a |
| ASA2 | 2.5a | 550.5a | 101.0b | 27.6a | 20.5a | 3.5b | 511.7c | 7.8c | 19.3b | 15.0a |
| LSD0.05 | ns | 21.0 | 2.6 | 2.5 | ns | 0.23 | 35.2 | 0.74 | 0.26 | ns |
| | 2020 season |
| ASA0 | 2.2c | 480.6b | 108.9a | 24.6b | 19.6b | 2.3c | 676.5c | 12.5b | 17.7c | 18.9c |
| ASA1 | 3.3a | 568.6a | 98.6b | 22.8c | 23.0a | 4.9a | 997.4a | 25.4a | 27.0a | 28.4b |
| ASA2 | 3.2b | 548.9a | 100.1b | 27.2a | 22.0ab | 3.8b | 856.2b | 23.1a | 26.4b | 38.0a |
| LSD0.05 | 0.08 | 20.5 | 4.4 | 0.17 | 2.7 | 0.04 | 1.1 | 4.12 | 0.50 | 0.58 |
Table 6.
Pearson’s correlation coefficients between total yield (TY) and each of leaf and root nutrient content of stressed sweet potato plants exogenously sprayed with different levels of zinc oxide nanoparticles and ascorbic acid in both the 2019 and 2020 seasons.
Table 6.
Pearson’s correlation coefficients between total yield (TY) and each of leaf and root nutrient content of stressed sweet potato plants exogenously sprayed with different levels of zinc oxide nanoparticles and ascorbic acid in both the 2019 and 2020 seasons.
| Nutrient | TY (t ha−1) |
|---|
| 2019 | 2020 |
|---|
| Pearson r | Probability | Pearson r | Probability |
|---|
| Leaves nutrients content |
| Phosphorus (P) | 0.263 ns | 0.186 | 0.462 * | 0.015 |
| Iron (Fe) | 0.279 ns | 0.159 | 0.658 ** | ˂0.001 |
| Manganese (Mn) | −0.407 * | 0.035 | −0.220 ns | 0.271 |
| Zinc (Zn) | −0.575 ** | 0.002 | −0.452 * | 0.018 |
| Copper (Cu) | −0.525 ** | 0.005 | 0.333 ns | 0.090 |
| Root nutrients content | | | | |
| Phosphorus (P) | 0.563 ** | 0.002 | 0.579 ** | 0.002 |
| Iron (Fe) | 0.113 ns | 0.574 | 0.334 ns | 0.089 |
| Manganese (Mn) | 0.402 * | 0.038 | 0.164 ns | 0.413 |
| Zinc (Zn) | −0.016 ns | 0.939 | 0.214 ns | 0.283 |
| Copper (Cu) | −0.262 ns | 0.186 | 0.097 ns | 0.631 |
Table 7.
Proportional contribution in predicting total tuberous root yield (RY) using stepwise multiple linear regression for stressed sweet potato plants sprayed with different levels of ascorbic acid and zinc oxide nanoparticles in both the 2019 and 2020 seasons.
Table 7.
Proportional contribution in predicting total tuberous root yield (RY) using stepwise multiple linear regression for stressed sweet potato plants sprayed with different levels of ascorbic acid and zinc oxide nanoparticles in both the 2019 and 2020 seasons.
| Year | RY (t ha−1) |
|---|
| r | Adjusted R2 | SEE | Significance | Fitted Equation |
|---|
| 2019 | 0.788 | 0.571 | 2.627 | *** | RY = 21.346 + 0.280Fe − 0.068Mn − 0.538Zn |
| 2020 | 0.876 | 0.724 | 1.862 | *** | RY = 12.498 + 0.032Fe − 0.056Mn − 0.403Zn − 0.133Cu |


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}