
Physiological, Biochemical, and Biometrical Response of Cultivated Strawberry and Wild Strawberry in Greenhouse Gutter Cultivation in the Autumn-Winter Season in Poland—Preliminary Study

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Cultivation Conditions, and Evaluation of Crop Productivity
2.2. Gas-Exchange Measurements
2.3. Biochemical Analysis of Leaves and Fruits
2.4. Statistical Analysis
3. Results
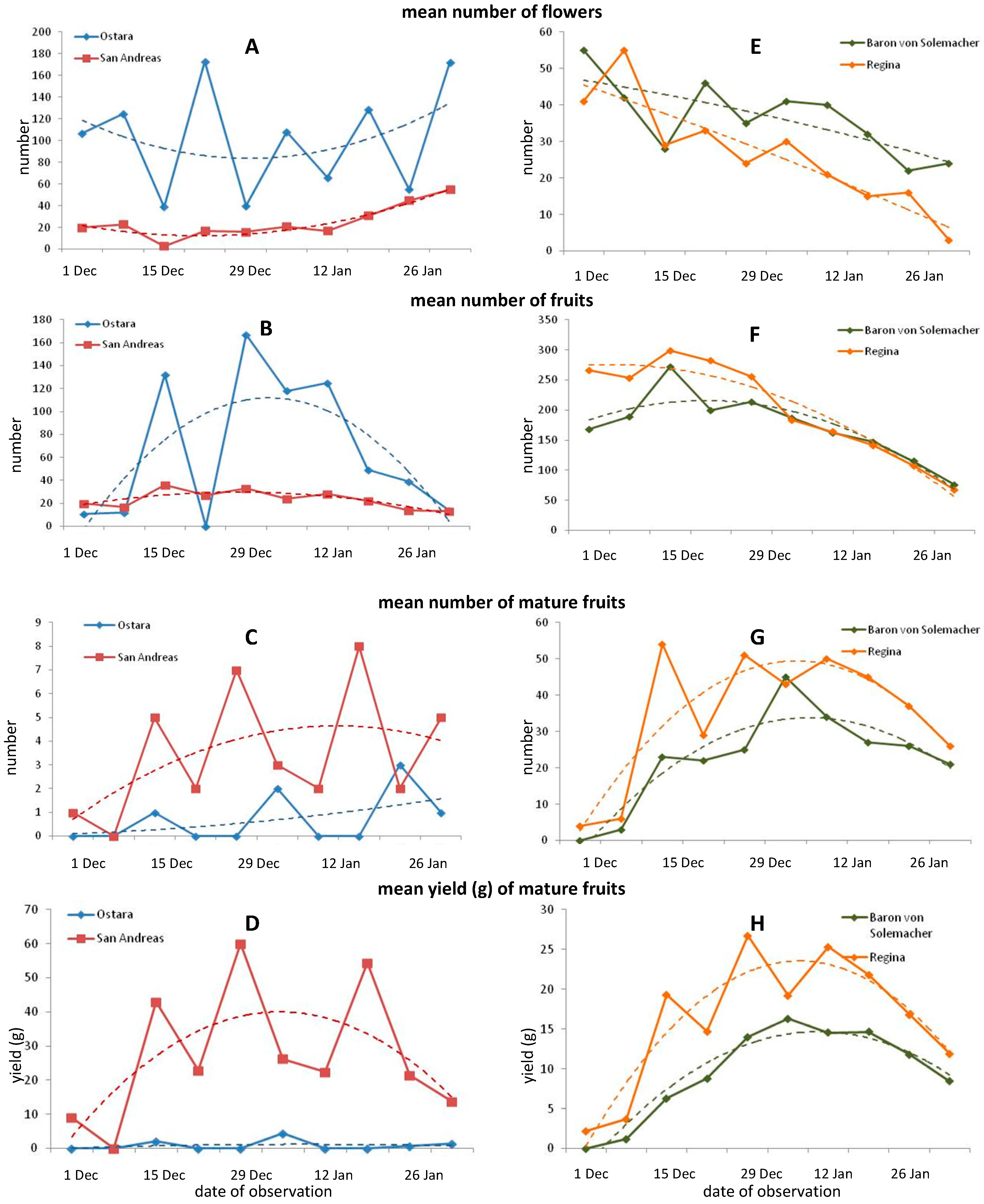
3.1. The Dynamics of Flowering and Yielding, and Biometrical Leaf Characteristic of Strawberry and Wild Strawberry
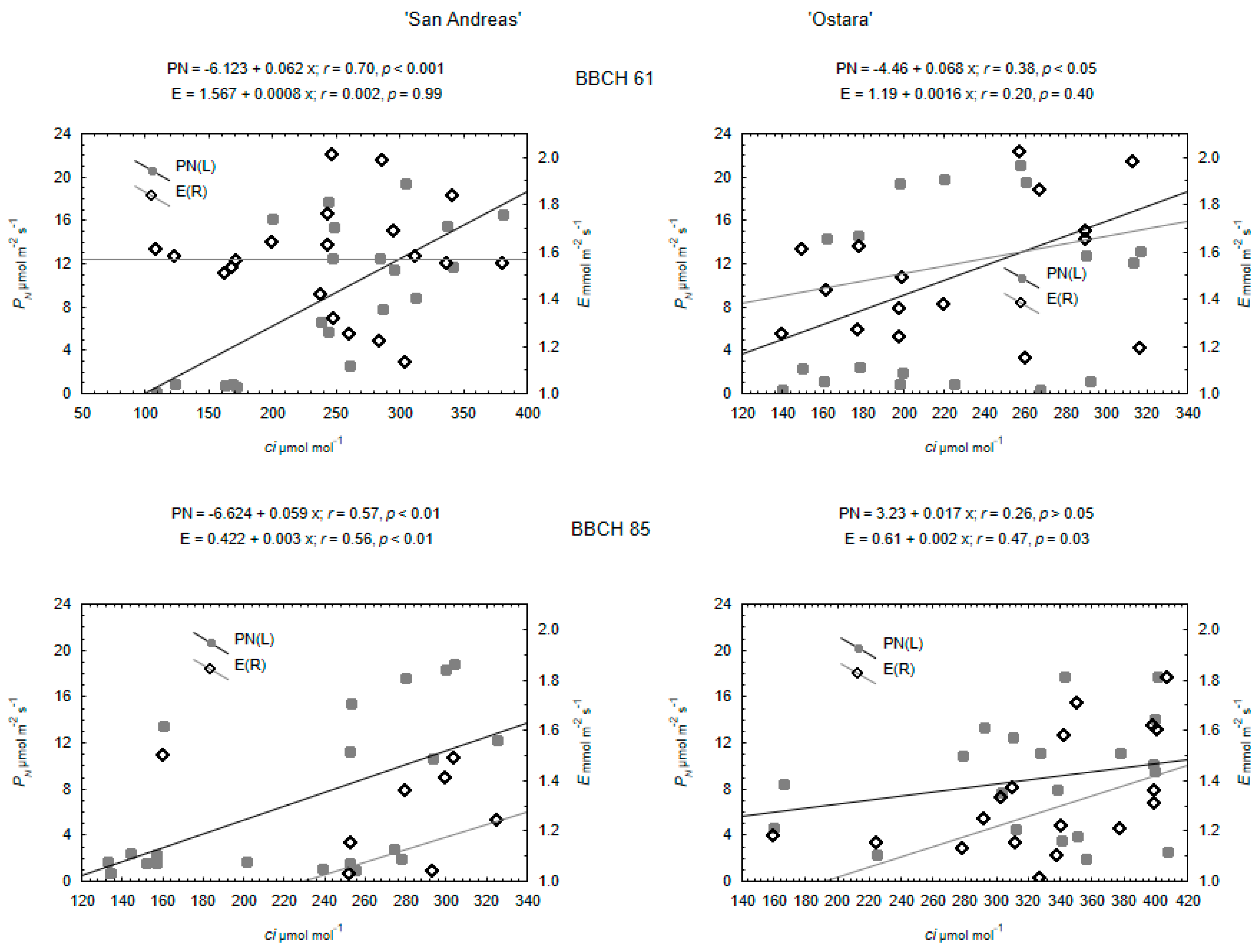
3.2. Physiological Response of Strawberry and Wild Strawberry
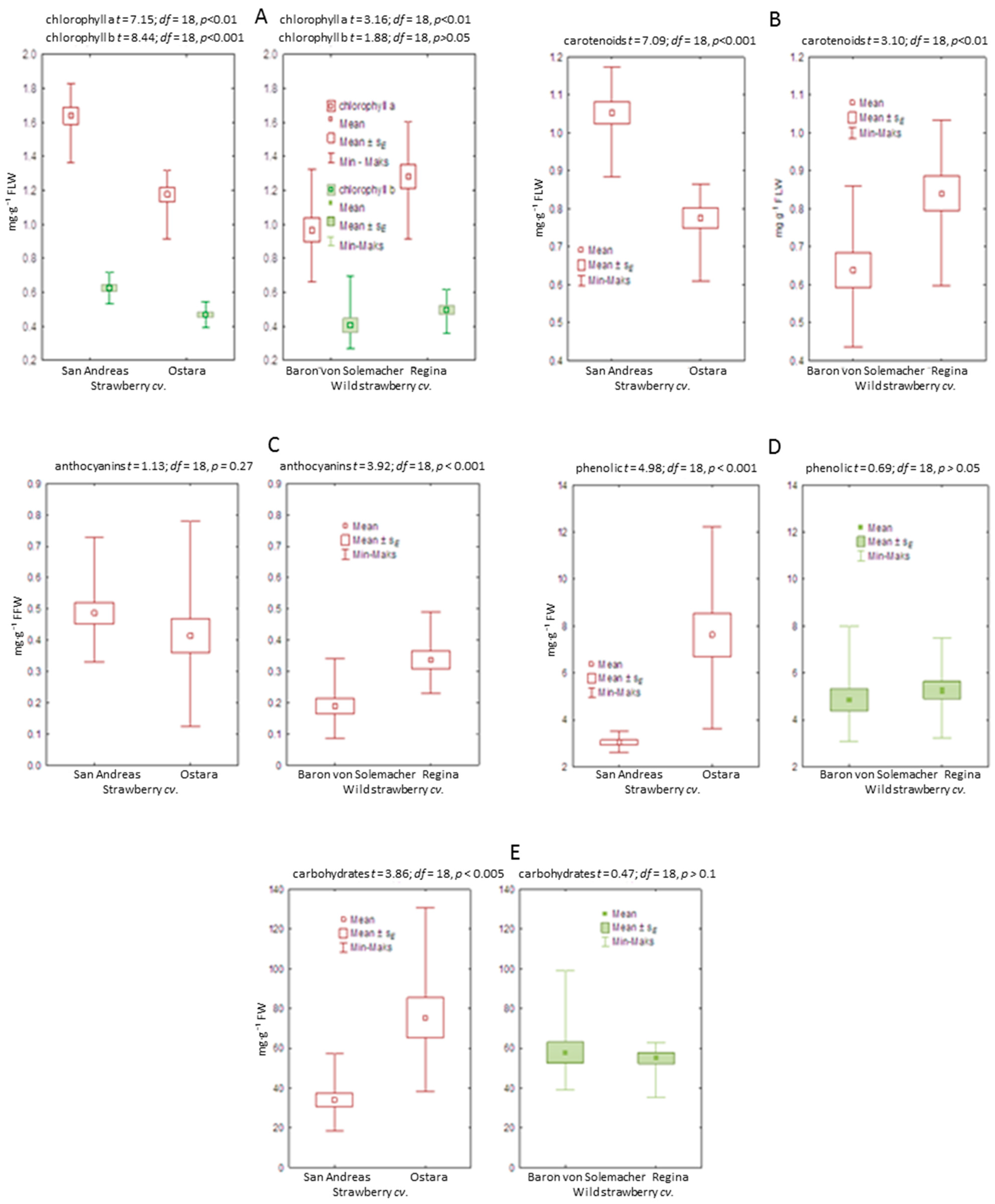
3.3. Biochemical Analysis of Leaves and Fruits
4. Discussion
4.1. The Dynamics of Flowering and Yielding, and Biometrical Leaf Characteristic of Strawberry and Wild Strawberry in Gutter Cultivation
4.2. Physiological Response of Strawberry and Wild Strawberry
4.3. Biochemical Analysis of Leaves and Fruits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef] [Green Version]
- Martins de Lima, J.; Welter, P.D.; Soares dos Santos, M.F.; Kavcic, W.; Costa, B.M.; Fagherazzi, A.F.; Nerbass, F.R.; Kretzschmar, A.A.; Rufato, L.; Baruzzi, G. Planting density interferes with strawberry production efficiency in Southern Brazil. Agronomy 2021, 11, 408. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 16 July 2021).
- Wang, S.Y.; Lin, H.S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef]
- Ulrich, D.; Olbricht, K. Diversity of volatile patterns on sixteen Fragaria vesca L. accessions in comparison to cultivars of Fragaria × ananassa. J. Appl. Bot. Food Qual. 2013, 86, 37–46. [Google Scholar] [CrossRef]
- Mozafari, A.; Havas, S.; Ghaderi, N. Application of iron nanoparticles and salicylic acid in in vitro cultures of strawberries (Fragaria × ananassa Duch.) to cope with drought stress. Plant Cell Tiss. Org. Cult. 2018, 132, 511–523. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; Lima, E.P.; D’Agostini, M.; Stockmans De Nardi, F.; Tretin, T.; Grando Dornelles, A.; Huzar-Novakowiski, J.; Calvete, E.O. Horticultural potential of nine strawberry cultivars by greenhouse production in Brazil: A view through multivariate analysis. Sci. Hortic. 2021, 279, 109738. [Google Scholar] [CrossRef]
- Andersen, Ø.M.; Fossen, T.; Torskangerpoll, K.; Fossen, A.; Hauge, U. Anthocyanin from strawberry (Fragaria ananassa) with the novel aglycone, 5-carboxypyranopelargonidin. Phytochemistry 2004, 65, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Palha, M.G.S. Strawberry growth and development in the mild winter european regions. Int. J. Fruit Sci. 2005, 5, 83–90. [Google Scholar] [CrossRef]
- Heide, O.M.; Stavang, J.A.; Sønsteby, A. Physiology and genetics of flowering in cultivated and wild strawberries—A review. J. Hortic. Sci. Biotechnol. 2013, 88, 1–18. [Google Scholar] [CrossRef]
- Khoshnevisan, B.; Shariati, H.M.; Rafii, S.; Mousazadeh, H. Comparison of energy consumption and GHG emissions of open field and greenhouse strawberry production. Renew. Sustain. Energy Rev. 2014, 29, 316–324. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; Ferreira, A.F.B.; Costa, R.C.; Tretin, N.; Tretin, T.; Calvete, E.O. Phyllochron, and root system development of six strawberry cultivars with different photoperiodic flowering responses. Sci. Agrar. Parana. 2020, 19, 368–373. [Google Scholar]
- Miranda, F.R.; Silva, V.B.; Santos, F.S.R.; Rossetti, G.; Silva, C.F.B. Production of strawberry cultivars in closed hydroponic systems and coconut fibre substrate. Rev. Ciênc. Agron. 2014, 45, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Galati, A.; Sabatino, L.; Prinzivalli, C.S.; D’Anna, F.; Scalenghe, R. Strawberry fields forever: That is, how many grams of plastics are used to grow a strawberry? J. Environ. Manag. 2020, 276, 111313. [Google Scholar] [CrossRef]
- Ahn, M.G.; Kim, D.S.; Ahn, S.R.; Sim, H.S.; Kim, S.; Kim, S.K. Characteristics and trends of strawberry cultivars throughout the cultivation season in a greenhouse. Horticulturae 2021, 7, 30. [Google Scholar] [CrossRef]
- Claire, D.; Watters, N.; Gendron, L.; Boily, C.; Pépin, S.; Caron, J. High productivity of soilless strawberry cultivation under rain shelters. Sci. Hortic. 2018, 232, 127–138. [Google Scholar] [CrossRef]
- Costa, R.C.; Calvete, E.O.; Mendonca, H.F.C.; Cecatto, A.P. Phenology, phyllochron, and gas exchanges in frigo and fresh strawberry (Fragaria × ananassa Duch.) plants of cv. Albion. Aust. J. Crop. Sci. 2014, 8, 901–908. [Google Scholar]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Michalska, A.; Carlen, C.; Heritier, J.; Andlauer, W. Profiles of bioactive compounds in fruits and leaves of strawberry cultivars. J. Berry Res. 2017, 7, 71–84. [Google Scholar] [CrossRef]
- Aarti, P.D.; Tanaka, R.; Tanaka, A. Effects of oxidative stress on chlorophyll biosynthesis in cucumber (Cucumis sativus) cotyledons. Phys. Plant 2006, 128, 186–197. [Google Scholar] [CrossRef]
- Ruiz-Sola, M.; Rodríguez-Concepción, M. Carotenoid biosynthesis in Arabidopsis: A colorful pathway. TAB 2012, 10, 158–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Bhusal, S.J.; Yoona, T.-M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Vemmos, S.N.; Petri, E.; Stournaras, V. Seasonal changes in photosynthetic activity and carbohydrate content in leaves and fruit of three fig cultivars (Ficus carica L.). Sci. Hortic. 2013, 169, 198–207. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2014, 79, 597–606. [Google Scholar] [CrossRef]
- Savchenko, T.; Tikhonov, K. Oxidative Stress-Induced Alteration of Plant Central Metabolism. Life 2021, 11, 304. [Google Scholar] [CrossRef]
- İnanç, A.L. Chlorophyll: Structural properties, health benefits and its occurrence in virgin olive oils. Akad. Gıda 2011, 9, 26–32. [Google Scholar]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar] [CrossRef] [PubMed]
- Masny, A.; Żurawicz, E. Season extension possibilities in two polish june-bearing strawberry cultivars. Acta Sci. Pol. Hortorum Cultus 2015, 14, 115–127. [Google Scholar]
- Lema-Rumińska, J.; Zalewska, M. Studies on flower pigments of chrysanthemum mutants: Nero and Wonder groups. Acta Sci. Pol. Hortorum Cultus 2004, 3, 125–135. [Google Scholar]
- Wettstein, D. Chlorophyll-letale und der submikroskopische Formwechsel der Plastiden. Exp. Cell Res. 1957, 12, 427–506. [Google Scholar] [CrossRef]
- Harborne, J.B. Comparative biochemistry of the flavonoids. Phytochemistry 1967, 6, 1569–1573. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterisazacion by UV-VIS Spectroscopy. In Current Protocols in Food Analytical Chemistry; Wiley: Hoboken, NJ, USA, 2001; Units F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Trevelyan, W.E.; Forrest, R.S.; Harrison, J.S. Determination of yeast carbohydrates with the anthrone reagent. Nature 1952, 170, 626–627. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. In Current Protocols in Food Analytical Chemistry; Wrolstad, R.E., Ed.; John Wiley & Sons: New York, NY, USA, 2001; pp. I1.1.1–I1.1.8. [Google Scholar] [CrossRef]
- Stadler, C. Effect of light intensity on yield of winter grown strawberries in Iceland. DGG Proc. 2017, 7, 1–5. [Google Scholar] [CrossRef]
- Hidaka, K.; Ito, E.; Sago, Y.; Yasutake, D.; Miyoshi, Y.; Kitano, M.; Miyauchi, K.; Okimura, M.; Imai, S. High yieldsof strawberry by applying vertically-moving beds on the basis of leaf photosynthesis. Environ. Control. Biol. 2012, 50, 143152. [Google Scholar] [CrossRef]
- Tang, Y.; Ma, X.; Li, M.; Wang, Y. The effect of temperature and light on strawberry production in a solar greenhouse. Solar Energy 2020, 195, 318–328. [Google Scholar] [CrossRef]
- Hidaka, K.; Dan, K.; Imamura, H.; Miyoshi, Y.; Takayama, T. Effect of supplemental lighting from different light sources on growth and yield of strawberry. Environ. Control. Biol. 2013, 51, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Nin, S.; Petrucci, W.A.; Giordani, E.; Marinelli, C. Soilless systems as an alternative to wild strawberry (Fragaria vesca L.) traditional open-field cultivation in marginal lands of the Tuscan Apennines to enhance crop yield and producers’ income. J. Hortic. Sci. Biotech. 2017, 93, 1–13. [Google Scholar] [CrossRef]
- Mott, K.A.; Parkhurst, D.F. Stomatal responses to humidity in air and helox. Plant Cell Environ. 1991, 14, 509–515. [Google Scholar] [CrossRef]
- Sperry, J.S.; Pockman, W.T. Limitation of transpiration by hydraulic conductance and xylem cavitation in Betula occidentalis. Plant Cell Environ. 1992, 16, 279–287. [Google Scholar] [CrossRef]
- Messinger, S.M.; Buckley, T.N.; Mott, K.A. Evidence for involvement of photosynthetic processes in the stomatal response to CO2. Plant Physiol. 2006, 140, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, K.I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keutgen, N.; Chen, K.; Lenz, F. Responses of strawberry leaf photosynthesis, chlorophyll fluorescence and macronutrient contents to elevated CO2. J. Plant Physiol. 1997, 150, 395–400. [Google Scholar] [CrossRef]
- Koziołek, C.; Grams, T.E.E.; Schreiber, U.; Matyssek, R.; Fromm, J. Transient knockout of photosynthesis mediated by electrical signals. New Phytol. 2004, 161, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Hinckley, T.M.; Braatne, J.H. Stomata. In Plant—Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 323–355. [Google Scholar]
- Nardini, A.; Salleo, S.; Raimondo, F. Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L. Trees 2003, 17, 529–534. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.H.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Aaby, K.; Mazur, S.P.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria × ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef]
- Krüger, E.; Will, F.; Kumar, K.; Celejewska, K.; Chartier, P.; Masny, A.; Mott, D.; Petit, A.; Savini, G.; Sønsteby, A. Influence of Post-Flowering Climate Conditions on Anthocyanin Profile of Strawberry Cultivars Grown from North to South Europe. Appl. Sci. 2021, 11, 1326. [Google Scholar] [CrossRef]
- Josuttis, M.; Dietrich, H.; Treutter, D.; Will, F.; Linnemannstöns, L.; Krüger, E. Solar UVB response of bioactives in strawberry (Fragaria × ananassa Duch.): A comparison of protected and open-field cultivation. J. Agric. Food Chem. 2010, 58, 12692–12702. [Google Scholar] [CrossRef]
- Bojarska, J.E.; Czaplicki, S.; Zarecka, K.; Zadernowski, R. Związki fenolowe owoców wybranych odmian truskawki. Żywność Nauka Technol. Jakość 2006, 2, 20–27. [Google Scholar]
- Cocetta, G.; Casciani, D.; Bulgari, R.; Musante, F.; Kołton, A.; Rossi, M.; Ferrante, A. Light use efficiency for vegetables production in protected and indoor environments. Eur. Phys. J. Plus 2017, 132, 43. [Google Scholar] [CrossRef]
- Aaby, K.; Remberg, S.V. Strawberry phenolics and impact of ripening. In Processing and Impact an Active Components in Food; Preedy, V., Ed.; Elsevier: London, UK, 2015; pp. 157–164. [Google Scholar] [CrossRef]
- Trouvelot, S.; Heloir, M.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trda, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horacio, P.; Martinez-Noel, G. Sucrose signaling in plants: A world yet to be explored. Plant Signal. Behav. 2013, 8, e23316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunachowicz, H.; Przygoda, B.; Nadolna, I.; Iwanow, K. Tabele Składu i Wartości Odżywczej Żywności; PWZL Wydawnictwo Lekarskie: Warsaw, Polska, 2017; p. 628. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Area (cm2) | Horizontal Width (cm) | Vertical Length (cm) | Perimeter (cm) | FW (g) | DW (g) |
|---|---|---|---|---|---|---|
| San Andreas | 37.99 ± 24.58 a | 7.57 ± 2.49 a | 11.32 ± 3.48 a | 41.94 ± 14.97 a | 3.01 ± 0.85 a | 0.93 ± 0.26 a |
| Ostara | 24.91 ± 7.46 b | 6.09 ± 1.40 a | 7.41 ± 2.14 b | 31.29 ± 7.08 b | 1.10 ± 0.30 b | 0.36 ± 0.10 b 1 |
| Cultivar | Area (cm2) | Horizontal Width (cm) | Vertical Length (cm) | Perimeter (cm) | FW (g) | DW (g) |
|---|---|---|---|---|---|---|
| Baron von Solemacher | 10.53 ± 4.58 b | 3.56 ± 1.12 b | 5.35 ± 1.35 b | 20.74 ± 5.72 b | 0.59 ± 0.18 a | 0.18 ± 0.04 a |
| Regina | 18.41 ± 4.28 a | 5.74 ± 1.05 a | 9.07 ± 1.82 a | 37.98 ± 7.79 a | 0.54 ± 0.15 a | 0.17 ± 0.04 a 1 |
| Cultivar | Stage | gs (mol·m−2·s−1) | Ci (μmol·mol−1) | PN (μmol·m−2·s−1) | E (mmol·m−2·s−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| San Andreas | 1 | 0.100 ± 0.005 | a | 324.3 ± 16.3 | a | 16.9 ± 6.28 | a | 1.569 ± 0.052 | a |
| 2 | 0.059 ± 0.005 | b | 227.4 ± 12.7 | b | 9.13 ± 4.93 | b | 0.993 ± 0.065 | b | |
| Mean | 0.080 ± 0.030 | A | 275.8 ± 12.8 | A | 13.0 ± 3.96 | A | 1.281 ± 0.062 | A | |
| Ostara | 1 | 0.100 ± 0.047 | a | 247.8 ± 16.5 | a | 9.29 ± 6.09 | a | 1.552 ± 0.102 | a |
| 2 | 0.080 ± 0.026 | a | 227.5 ± 14.6 | a | 5.22 ± 2.51 | b | 1.267 ± 0.070 | a | |
| Mean | 0.090 ± 0.039 | A | 237.5 ± 11.0 | B | 7.25 ± 3.27 | B | 1.409 ± 0.065 | A 1 | |
| Cultivar | Stage | gs (mol·m−2·s−1) | Ci (μmol·mol−1) | PN (μmol·m−2·s−1) | E (mmo·m−2·s−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Baron von Solemacher | 1 | 0.144 ± 0.012 | a | 249.8 ± 13.1 | a | 5.80 ± 2.36 | a | 1.947 ± 0.109 | a |
| 2 | 0.062 ± 0.004 | b | 245.2 ± 15.3 | a | 5.02 ± 1.91 | a | 1.087 ± 0.056 | b | |
| Mean | 0.103 ± 0.009 | A | 247.5 ± 9.95 | B | 5.41 ± 1.07 | B | 1.517 ± 0.092 | A | |
| Regina | 1 | 0.145 ± 0.015 | a | 292.9 ± 8.65 | a | 7.20 ± 1.24 | a | 1.963 ± 0.119 | a |
| 2 | 0.055 ± 0.026 | b | 245.3 ± 18.8 | b | 7.13 ± 2.90 | a | 0.998 ± 0.056 | b | |
| Mean | 0.100 ± 0.004 | A | 269.1 ± 10.9 | A | 7.17 ± 0.96 | A | 1.480 ± 0.101 | A 1 | |
| Cultivar | Fruits | Leaves | ||||
|---|---|---|---|---|---|---|
| Anthocyanins | Phenolics | Carbohydrates | Carotenoids | Chlorophyll a | Chlorophyll b | |
| (mg·g−1 FW) | ||||||
| San Andreas | 0.486 ± 0.034 a | 3.04 ± 0.11 b | 34.07 ± 3.49 b | 1.052 ± 0.029 a | 1.639 ± 0.049 a | 0.623 ± 0.019 a |
| Ostara | 0.414 ± 0.054 a | 7.62 ± 0.91 a | 75.35 ± 10.11a | 0.775 ± 0.025 b | 1.175 ± 0.041 b | 0.467 ± 0.014 b 1 |
| Cultivar | Fruits | Leaves | ||||
|---|---|---|---|---|---|---|
| Anthocyanins | Phenolics | Carbohydrates | Carotenoids | Chlorophyll a | Chlorophyll b | |
| (mg·g−1 FW) | ||||||
| Baron von Solemacher | 0.190 ± 0.023 b | 4.85 ± 0.47 a | 57.86 ± 5.17 a | 0.638 ± 0.046 b | 0.965 ± 0.071 a | 0.404 ± 0.041 a |
| Regina | 0.337 ± 0.028 a | 5.26 ± 0.37 a | 55.10 ± 2.73 a | 0.840 ± 0.045 a | 0.495 ± 0.025 b | 0.337 ± 0.026 a 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lema-Rumińska, J.; Kulus, D.; Tymoszuk, A.; Miler, N.; Woźny, A.; Wenda-Piesik, A. Physiological, Biochemical, and Biometrical Response of Cultivated Strawberry and Wild Strawberry in Greenhouse Gutter Cultivation in the Autumn-Winter Season in Poland—Preliminary Study. Agronomy 2021, 11, 1633. https://doi.org/10.3390/agronomy11081633
Lema-Rumińska J, Kulus D, Tymoszuk A, Miler N, Woźny A, Wenda-Piesik A. Physiological, Biochemical, and Biometrical Response of Cultivated Strawberry and Wild Strawberry in Greenhouse Gutter Cultivation in the Autumn-Winter Season in Poland—Preliminary Study. Agronomy. 2021; 11(8):1633. https://doi.org/10.3390/agronomy11081633
Chicago/Turabian StyleLema-Rumińska, Justyna, Dariusz Kulus, Alicja Tymoszuk, Natalia Miler, Anita Woźny, and Anna Wenda-Piesik. 2021. "Physiological, Biochemical, and Biometrical Response of Cultivated Strawberry and Wild Strawberry in Greenhouse Gutter Cultivation in the Autumn-Winter Season in Poland—Preliminary Study" Agronomy 11, no. 8: 1633. https://doi.org/10.3390/agronomy11081633
APA StyleLema-Rumińska, J., Kulus, D., Tymoszuk, A., Miler, N., Woźny, A., & Wenda-Piesik, A. (2021). Physiological, Biochemical, and Biometrical Response of Cultivated Strawberry and Wild Strawberry in Greenhouse Gutter Cultivation in the Autumn-Winter Season in Poland—Preliminary Study. Agronomy, 11(8), 1633. https://doi.org/10.3390/agronomy11081633