1. Introduction
The germination rate of non-dormant seeds is controlled by multiple factors, with water availability generally considered to be one of the most important. Reduced water availability, for example in experiments utilizing PEG as an osmoticum, or in dry soils, reduce water uptake and seed germination rates [
1], with stronger solutions / drier soils repressing germination rate to a greater degree [
2]. Despite a general understanding of the link between reductions in water uptake and germination, a detailed physiological explanation of the mechanism(s) remains elusive. Plant hormones including abscisic acid (ABA) [
3], cytokinins, gibberellins and Indole-3-acetic acid (IAA) have been suggested to have a role [
4], although it is unclear whether these are a cause of, or a response to, changes in seed water content.
As with PEG, salt treatment generally reduces germination percentages and rates, relative to water controls. Salt is reported to affect seed germination by osmotic effects, ion toxicity effects, or a combination of the two [
4,
5,
6]. Differences in the germination rates and percentages between seeds incubated in salt and PEG are generally attributed to “ionic effects”, which can be either negative or positive. In some halophytic (salt-tolerant) species, incubation in high salt concentrations can arrest germination, although germination percentages can often recover when these seeds are transferred to pure water [
7]. It is unclear whether arrested germination is directly due to osmotic effects, or to some non-toxic germination repression caused by the salt, although it has been noted that high concentrations of Na
+ and Cl
− ions can inhibit critical metabolic steps in cell division [
8]. In some species, moderate salt levels have been shown to promote germination [
4], and it has been suggested that salt can be compartmentalized and used as a cellular osmotica, allowing seeds to germinate under osmotic conditions which would otherwise preclude it [
9,
10]. Compartmentalization of salt into vacuoles has been shown to aid the growth and survival of vegetative plants [
11,
12].
The most widely used germination model, the hydrothermal time model, posits a base water potential designated Ψ
b (50), at which half of the viable seeds in a population cannot germinate. The physiological basis of Ψ
b remains unclear, yet is presumably related to the seeds ability to take up water. While both salt and PEG are able to exert osmotic effects, their effect are often not identical. As PEG is either unable to cross cell membranes, or can do so only slowly, it tends to act purely as an osmotica. Conversely, as mentioned above, at least in some species, salt can be taken up and compartmentalized, allowing seeds to avoid the negative consequences of low water potential environments. Few studies have attempted to directly quantify the kinetics of seed water uptake and relate it to germination mechanics. McDonald et al. [
13] measured water uptake by maize seeds over a 48-h period, demonstrating the existence of two separate pathways of water uptake. The first pathway was rapid, and related to water uptake into the embryo, while the second involved a more gradual wetting of the endosperm, likely associated with starch breakdown. Other studies have used imaging techniques to investigate water uptake by seeds. Kikuchi et al. [
14] noted rapid uptake of water into the testa and cotyledons of kidney beans following a lag period of a few hours. Munz et al. [
15] revealed that water uptake followed a coordinated series of steps featuring multiple pools in oilseed rape seeds.
The main aim of this paper is to gain a better understanding of water uptake kinetics by seeds, particularly in relation to environmental water potential. Kinetic studies are frequently used to provide information about the regulation of complex systems, for example, enzymatic properties, biochemical pathways or sink / source relationships between plant organs. Here, we measured water uptake by wheat seeds and developed a mathematical model, which allowed us to compare the effects of salt and PEG on the rate of water uptake into different pools. Wheat (Triticum aesitvum L.) seeds were used due to their relatively large size, which allowed reliable estimates of water uptake rates based upon mass increase through time. Water uptake rates and contents were compared between seeds incubated in pure water, a −1MPa NaCl solution, and a −1 MPa PEG solution, both under constant conditions, and where the osmotic conditions were changed part-way through the experiment. Given the assumption that seeds can take up salt but not PEG, we hypothesized that water uptake rates would be lower in seeds incubated in PEG than in either the water or salt treatments, and that this would have a detectable effect on water uptake kinetics. We also hypothesized that when seeds were transferred between treatments, the initial treatment conditions would influence the final water content of the seeds.
2. Materials and Methods
All experiments described in this study used seeds from a single seed batch of a locally developed commercially available wheat variety, “Aoba”. Seeds between 40 and 50 mg were used, this representing approximately the middle 50th percentile, with larger and smaller seeds excluded.
Osmotica were made using either PEG 6000 or NaCl dissolved in milliQ grade water. PEG solutions were prepared according to Equation (5) from Michel [
16] (Equation (1), below), and checked using a WP4 Dewpoint Osmometer (Decagon, Pullman, WA, USA).
where [PEG] is the concentration of PEG required (g g
−1 H
2O) to give a desired water potential (Ψ), at a given temperature (T).
Salt solutions were made according to Equation (2), which was derived from a dataset of measured values of water potential in relation to salt concentration at a range of temperatures. The dataset measurements were found to correspond well with published values [
17].
where [NaCl] was the concentration of salt (g L
−1) required to produce a solution of a desired water potential (Ψ) at 5 °C.
Seeds were placed equidistant from each other in a grid pattern on Whatmann 3 mm CHR chromotography paper wetted with the test solution of interest and sealed in a 15 cm diameter petri-dish. A small volume of extra solution was placed in each container, to reduce matric effects caused by the filter paper, and to buffer the system against water uptake by the seeds. Additional solution was added, or the chromatography paper/solution replaced on an “as necessary” basis. It was calculated that water uptake by seeds would be able to alter the concentration of the solution by less than 3%, and the petri dishes were sealed to minimize evaporative losses. Changing old solution for fresh solution had little impact on seed water content, confirming that the osmotic potentials of the solutions did not change greatly throughout the experimental period. Each experiment was conducted only once, but they were part of a larger series of trails, with water uptake patterns consistent across all experiments.
2.1. Experiment 1
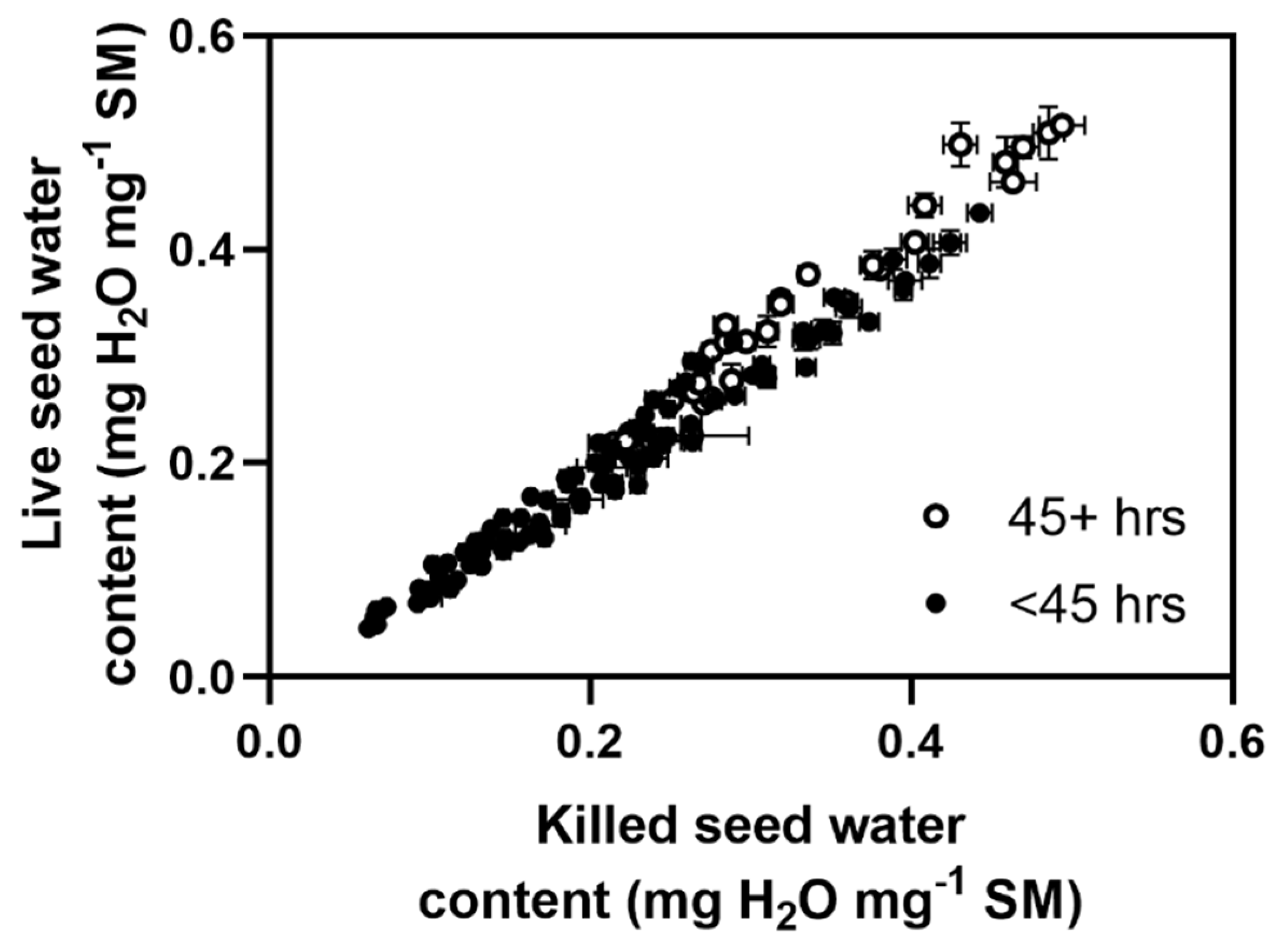
This experiment aimed to compare the water uptake kinetics of killed seeds, where the embryo had been destroyed with a hot needle, versus live seeds (
Figure 1). Killed seeds are preferable for measuring water uptake as they do not germinate, allowing longer water uptake time courses to be obtained. Thirty-two live and 32 killed seeds were used in each trial, which was conducted at 14 °C under a range of PEG-generated water potentials (0, −0.23, −0.35, −0.47. −0.58, −0.77, −1.05, −1.42, −1.80, −2.18 and −2.46 MPa) to promote a range of water uptake rates in the seeds. At the start of the experiment, then 3, 6, 9, 12, 20, 27, 36, 45, 56, 69 and 84 h after the start of incubation, individual seed fresh weight was recorded to the nearest 0.1 mg. After 12, 27, 45, and 69 h, six replicate seeds were removed from each treatment, weighed, dried and re-weighed. After 84 h the remaining eight seeds for each treatment were weighed and the experiment terminated. Ten additional control seeds were weighed then dried at 80 °C for seven days and re-weighed, without being immersed in any test solution. The dry mass of the test seeds (approximately totalling 750) was compared with the dry mass of the control seeds, with no significant differences in mass noted between seed lots, suggesting no loss of mass due to respiration. In subsequent experiments it was therefore assumed that seed dry weight remained relatively constant throughout the experiments, and it was not further tested.
2.2. Experiment 2
This experiment had two goals. The first was to characterize water uptake kinetics in wheat seeds incubated in deionized water, −1 MPa NaCl and −1MPa PEG solutions. This water potential was selected based upon a pre-experiment, where we observed that a −1 MPa water potential was sufficient to cause a reduction in seed germination rate, and in the case of PEG, to cause a significant reduction in final seed water content. The second goal was to investigate the effects on water uptake of transferring the seeds between either of the osmotica and deionized water, after 8 or 96 h. These time points were chosen as they represent the seeds taking water up into pool 2 and pool 3, thought to be the embryo and endosperm respectively. Killed seeds were used in this experiment, with 12 seeds per treatment. Seeds were individually weighed prior to the start of treatments, then periodically throughout the treatment period in order to estimate water uptake. Measurement times were adjusted to try and capture detailed information about the curve. For example, being more frequent when water uptake was most rapid or immediately following transfer between treatments, and less frequently when water uptake was less rapid. The exact time points can be found in the associated dataset. Each seed was dried using tissue paper, weighed to the nearest 0.1 mg, and returned to the same solution. This experiment was conducted at 5 °C, with the controls (i.e., seeds maintained in water, NaCl or PEG for the whole experiment, without being transferred) incubated for 338 h, while the reciprocal transfer treatments were terminated after 314 h.
2.3. Data Analysis
All seed water contents (SWC) were expressed as mg of water per mg of the initial seed mass (SM). Experiment one showed that the initial water content of the seeds was very uniform, allowing us to ignore this without significantly affecting the results.
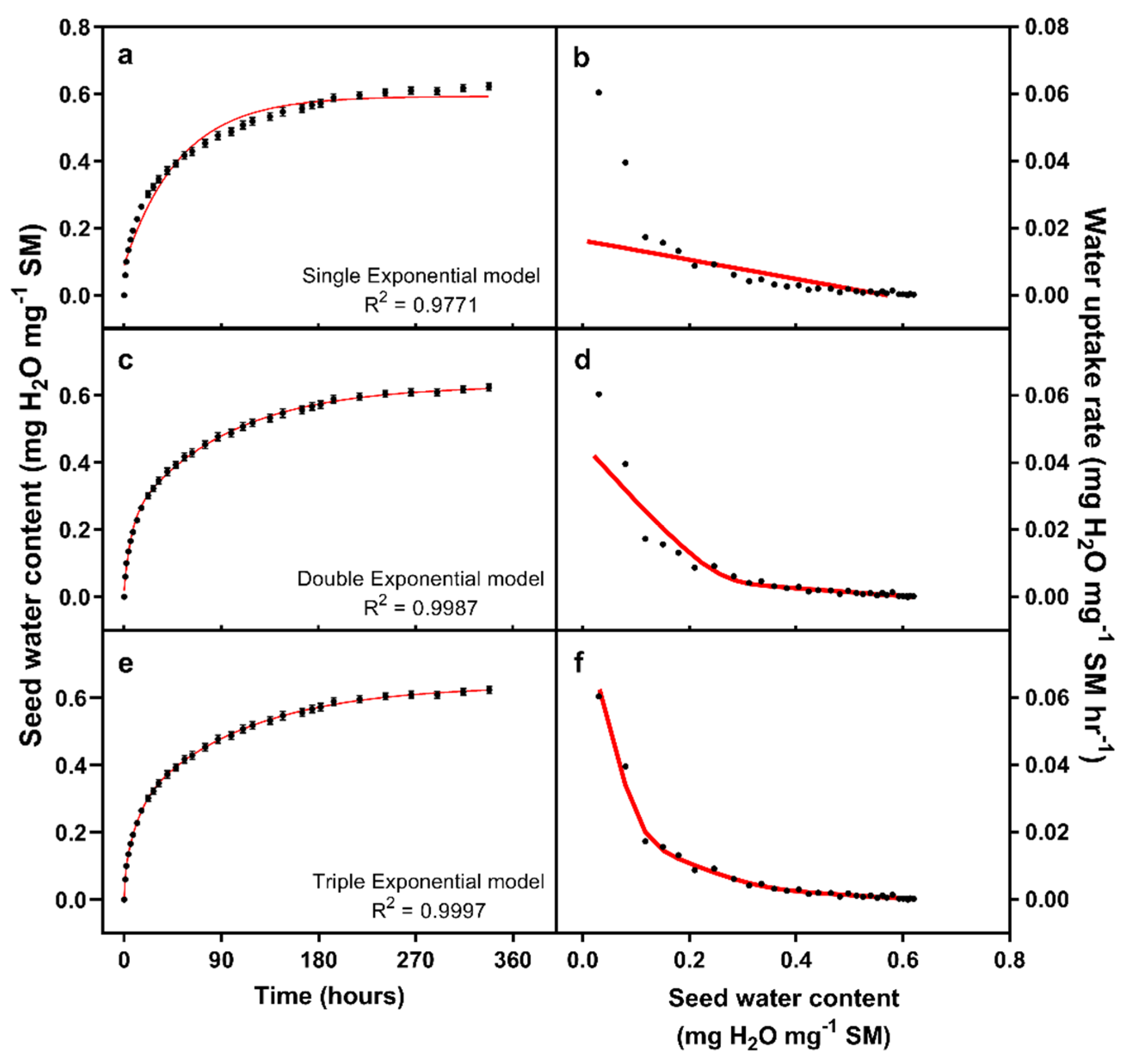
We used an exponential-rise-to maximum curve framework to model water uptake, following Finch-Savage et al. [
1]. This model type, with a single exponential rise-to-max curve shown in Equation (3), can be easily understood in mechanistic terms, with water passing into a single pool with a max (
m), over time (
t), at a rate determined by the rate constant (
K), proportional to the seed water content (
SWC).
In terms of the underlying biology, this type of model often describes diffusional processes, particularly where a membrane is involved.
Equation (4) shows a double exponential model, with two pools, each with a maximum value (
m1 and
m2), and a rate constant (
K1 and
K2). Within our context, we may understand these as “fast” and “slow” pools, respectively.
Equation (5) shows a triple exponential model, representing three pools, each with maxima and rate constants.
More complex exponential model implies multi-phasic water uptake, with water being absorbed into two or more pools. The maximum seed water content is the sum of m-values for each pool, while the rate of water flux into each pool would be described by a linear relationship between the seed water content and time, with the rate constant being the gradient of this relationship.
Curve fitting was conducted in PRISM v9.1.2 (Graphpad software, San Diego, CA, USA), using a least squares regression method. Normality of a residuals was tested by Shapiro–Wilk test, while a Runs test was used to test for systemic lack of fit between the curve and the data. The Extra-sum-of-squares F-test was used to determine which curve to fit. Briefly, the extra-sum of squares test compares whether a more complex model fits the data better than can be explained purely from increased model complexity, compared to a simpler model.
Seed water uptake rate was estimated as the mass difference between two adjacent time points (or curve-fit predictions), divided by the time difference. The seed water content was estimated as the mean seed water content of any two adjacent time points.
Differences in seed water content for different treatments were compared using the GLM command, with a post-hoc REGWF test to identify group differences in IBM SPSS v.27 (IBM corporation, Armonk, NY, USA). Data was tested for heteroskedasticity and log-transformed where necessary.
4. Discussion
Our data suggests the existence of three distinct pools into which water is absorbed by wheat seeds. The existence of multiple pools and multiphasic water uptake is consistent with previous studies in wild oat [
18], maize [
13] and soybean [
19]. The first pool is the smallest-comprising around 11–13% of the total capacity. It has a very high-rate constant, filling in just a few hours. This first pool is relatively unaffected by salt, but absorption of water was slowed by the PEG solution. It seems likely that this pool represents apoplastic components of the seed, such as cell walls [
20]. It has been estimated that the water potential of seeds can be as low as −400 MPa [
21], which is mainly considered to be attributable to matric effects.
The second pool comprises around 30% of the total capacity. The rate constant is around one-tenth that of the first pool, with a half-life of around eight hours in the water control treatment. Under optimal conditions, wheat and barley seeds can germinate in as little as 24 h, at which time we would expect the first pool to be completely saturated, and this second pool at 80–90% of its maximum capacity. This pool is seemingly weakly affected by salt, with the half-life increasing by around 50%, but no change in its maximum size. Given the influence of salt on water uptake to this pool, this suggests a more biological than physical explanation, with water crossing a membrane over which salt can slowly cross. We assume this pool to be cellular. Although the embryo is relatively small, accounting for only around 3% of the mass of the seed [
20], it has been shown to take up several times its own mass when incubated in water [
22].
The third pool is the largest, and slowest to fill. Its rate constant is an order of magnitude lower than for the second pool, taking 70 h to half-fill in the water controls, and nearly 100 h in the salt treatment. The rate constant is affected by salt, suggesting this to be a biological phenomenon, which we assume to be starch breakdown. Previous investigators have noted a reduction in the rate of α-amylase-mediated starch degradation to soluble sugars when seeds are incubated in saline solutions [
23,
24,
25], with these sugars acting as osmotica.
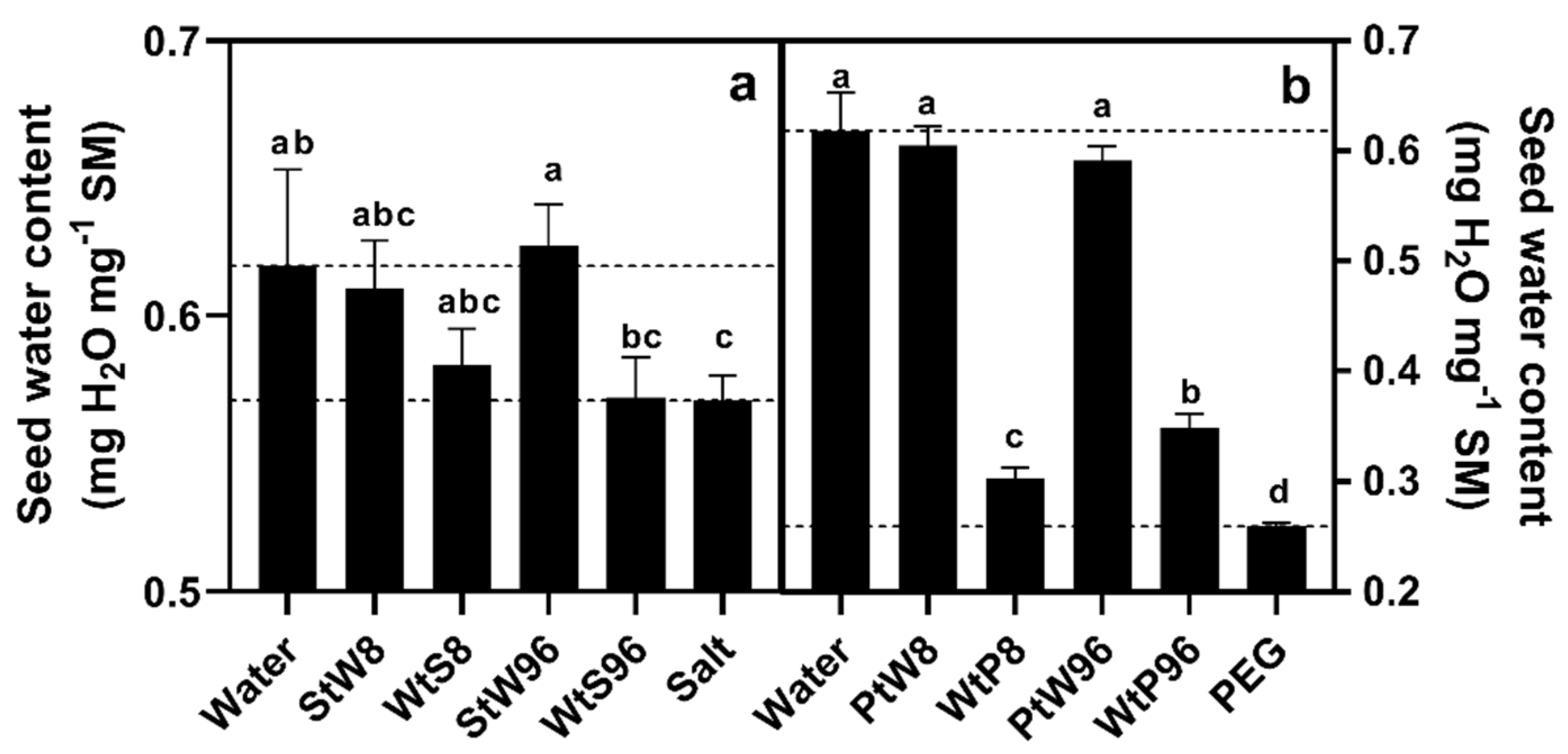
The water uptake curve of the PEG incubated seeds was best described by a double-exponential model. The maximum water content achieved by the PEG treatment was 41.4% that of the water control seeds. The rate constants were the same order of magnitude as those of the fast and medium pools identified in water control seeds, yet the values were lower than those for salt or water controls.
Switching seeds between the water and salt treatments led to the seeds following the water uptake curve of the solution they were exposed to. This was particularly clear in the seeds switched after 96 h, with the water to salt seeds achieving a maximum water content lower than the water controls and not significantly different with the salt controls. The salt to water seeds similarly achieved a maximum water content not different from the water controls, and significantly higher than the salt controls. In studies where germination is suppressed by salt, germination percentages often recover after the seeds are transferred to pure water [
7,
26]. This is often used to conclude that the retardation of germination is caused by osmotic effects. For this conclusion to be valid, we would expect to see similar reduction in germination percentage where PEG solutions were used. Zhang et al. [
10] showed in two barley genotypes that germination percentages were more strongly suppressed by PEG than salt, concluding that both salt and water were being taken up by the seeds. This corresponds strongly with our study, where NaCl incubated seeds achieved final water contents only 4.8% lower than water control seeds, suggesting that any osmotic effects are very mild. An alternate plausible explanation for salt suppression of germination may be that osmotic factors are relatively unimportant, with reductions in germination being a direct result of salt entering the embryo, which has been shown to inhibit seed germination at low concentrations and kill the seed at high concentrations [
27]. Transfer to pure water would presumably allow salt export from the seed.
Switching between water and PEG had a stronger effect. Transfer from water to PEG resulted in final water contents significantly higher than PEG controls, with this increase significantly greater in the 96-h transfer than the 8-h transfer seeds. This suggests that some changes occurred in the seeds during the period they were incubated in water, which could not be undone by the PEG solution. Particularly in the 96-h seeds, it seems evident that the third water pool is involved in this change, suggesting that starch breakdown accounts for this phenomenon. Previous studies have reported that seed priming-soaking seeds in water for between 10 and 24 h prior to planting leads to improvements in germination percentages and plant establishment, as well as the subsequent vigour, disease resistance and ultimately yield of crop plants in semi-arid tropical areas [
1,
28]. It seems plausible that improved water and carbohydrate availability during germination and seedling establishment may at least partially explain these observations [
29].
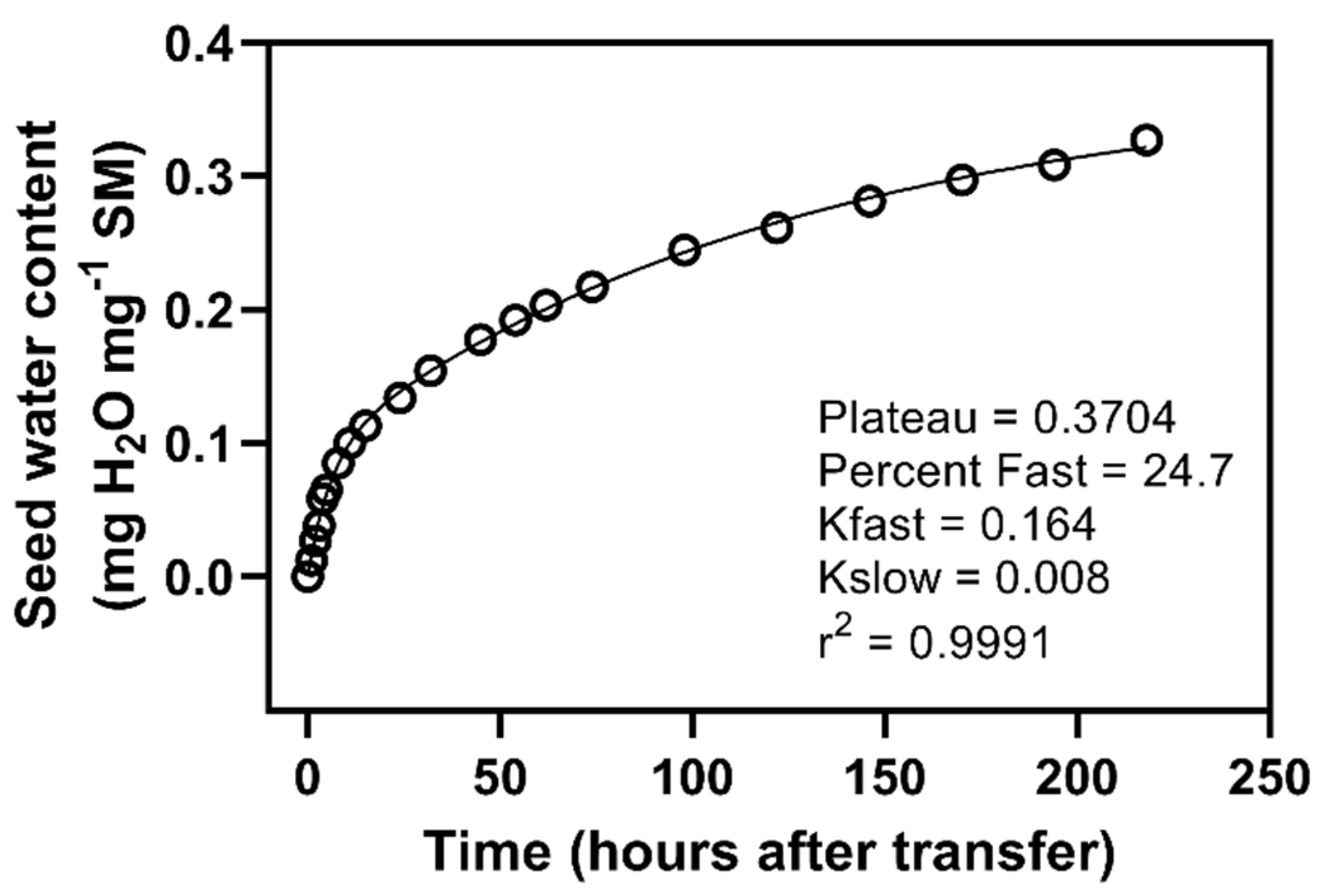
Transferring from PEG to water led to the seeds achieving maximum water contents not different from water control seeds. Isolating post-transfer water uptake data in the 96-h seeds allowed curve fitting, and an investigation of water uptake kinetics (
Figure 6). A double-exponential curve fitted the data well, with half-lives of around 4 and 86 h for the two pools. The faster pool represented around 25% of the total water uptake. The use of a double-exponential fit to this data suggests the existence of two pools. The larger of these pools has a rate constant (8.01 × 10
−3) consistent with the slow pool of water control seeds, which we hypothesize to be related to starch breakdown. The smaller pool has a rate constant (1.64 × 10
−1) intermediate between pools one and two, although more strongly influenced by the second. Our data appears to suggest that some amount of starch breakdown occurs even under the PEG control conditions, although water uptake was suppressed.
Within the context of our experiment, water uptake rates may be described by changes in the rate constants and sizes of the three pools. PEG appears to have a strong ability to alter both water uptake rate and the maximum water content, while salt caused declines in the rate constants of the second and third pool, with the maximum water content less affected. This may explain the greater germination percentages achieved in iso-osmotic salt solutions compared with PEG solutions [
10]. Our data furthermore suggests the existence of a minimum water content, under which the seeds cannot germinate. Speculatively, seed to seed variation may be explained by differences in factors which control water uptake, such as seed size, membrane permeability, or the activities of key enzymes.
In conclusion, using killed seeds, we characterised water uptake into at least three pools, comprising approximately 11%, 28%, and 60% of the total water uptake capacity, with half-lives of 0.7, 8, and 70 h in water, respectively. Incubating seeds in a −1 MPa salt solution slowed the rate of water uptake into the second and third pools but caused only a small decline in maximum water content. When the seeds were incubated in −1 MPa PEG, water uptake rates and the maximum water content were strongly suppressed. Transferring seeds between water and salt solutions had only a small effect on water uptake, while transferring between water and PEG had strong effects on water uptakes. Notably, seeds incubated in water then transferred to PEG exhibited maximum seed water contents higher than PEG controls, which may explain the benefits of pre-soaking seeds before planting.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}