
First Steps to Domesticate Hairy Stork’s-Bill (Erodium crassifolium) as a Commercial Pharmaceutical Crop for Arid Regions


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ecosystem and Plant Material
2.2. Field Experiments
2.3. Fertigation
2.4. Plant Density Trials (2009–2011)
2.5. Ecotype Examination (2016–2018)
2.6. Fertilizer Trials (2016–2018)
2.7. Source-Sink Manipulation (2016–2018)
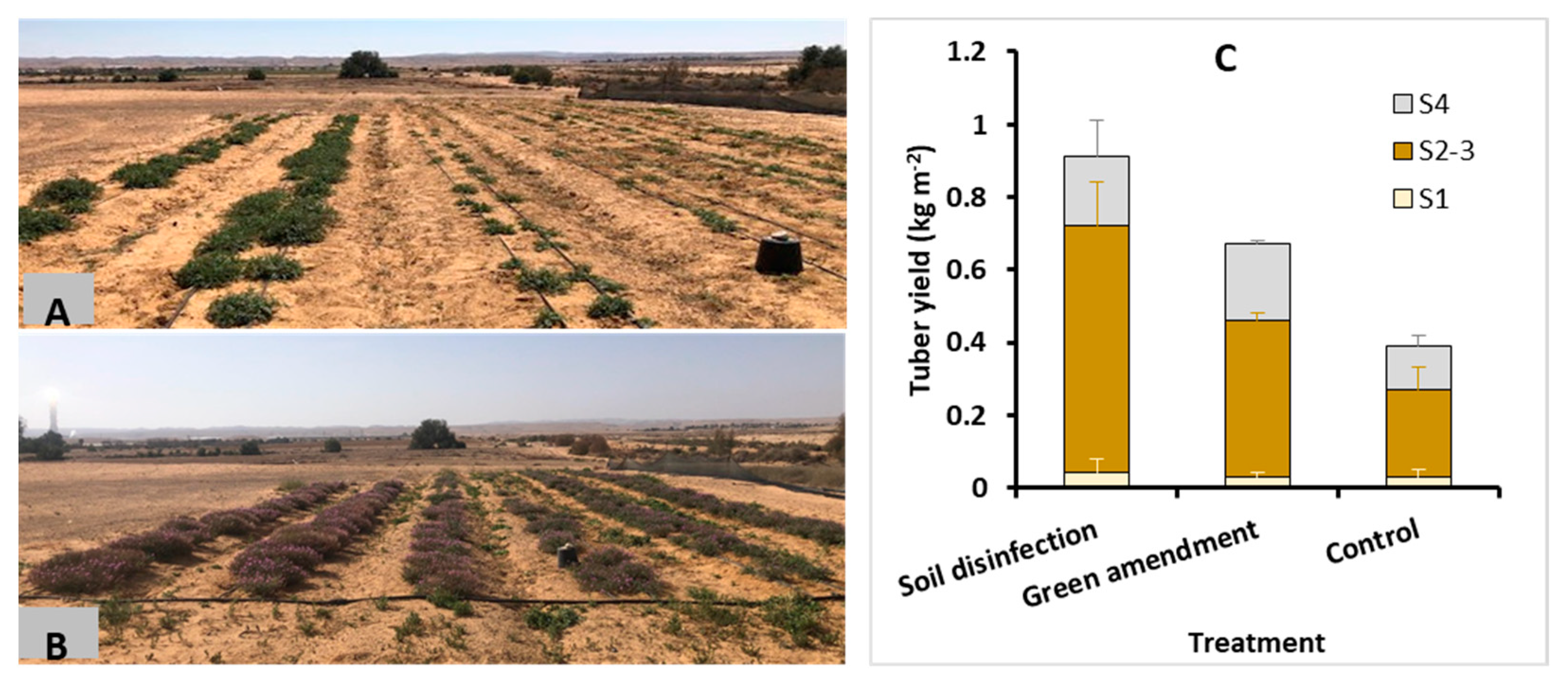
2.8. Soil Recovery (2020–2021)
2.9. Measurements
2.10. Ethanolic Extraction (EE) of HSB Tubers
2.11. Human Cell Culture and IL-8 Determination
3. Results and Discussion
3.1. Water Availability Determines Crop Productivity
3.2. Crop Density
3.3. Mineral Nutrition
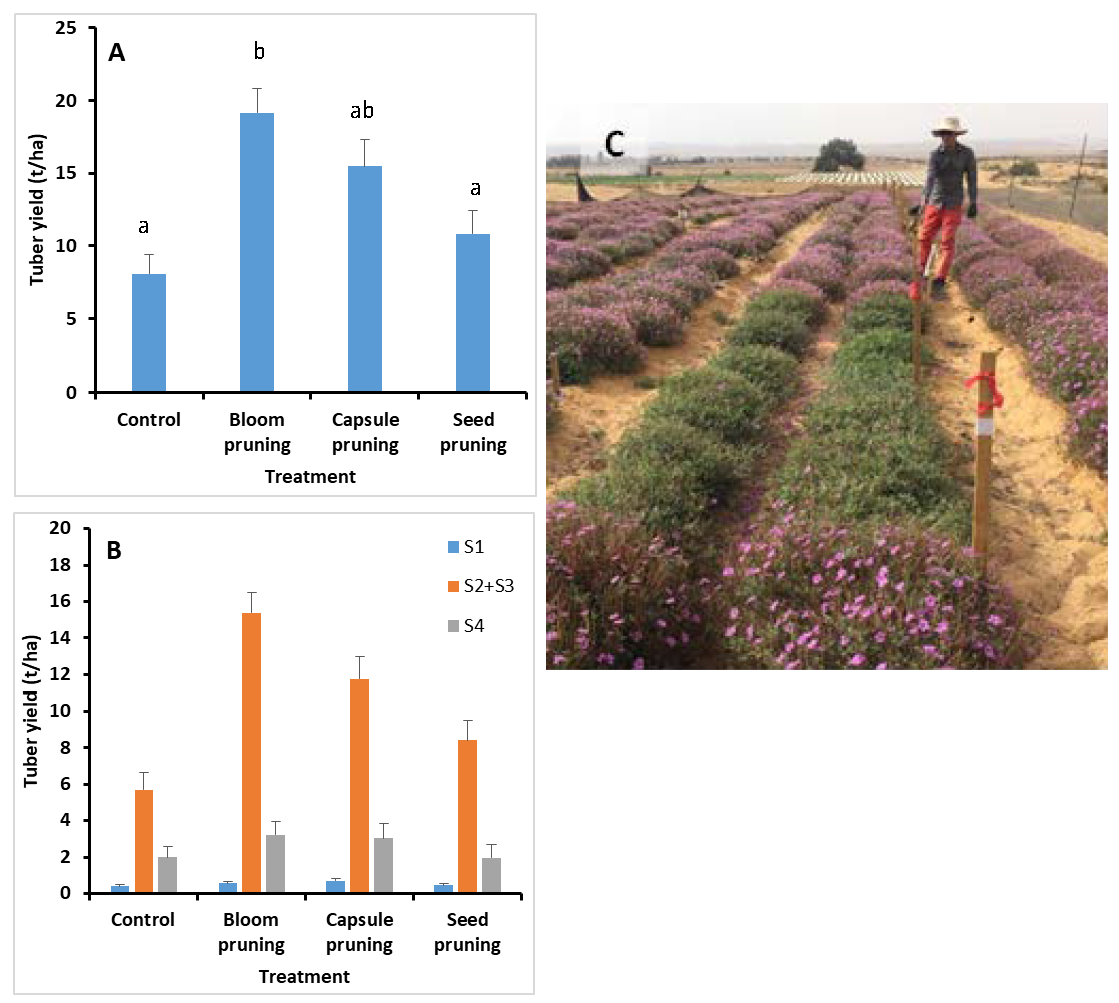
3.4. Source-Sink Manipulations
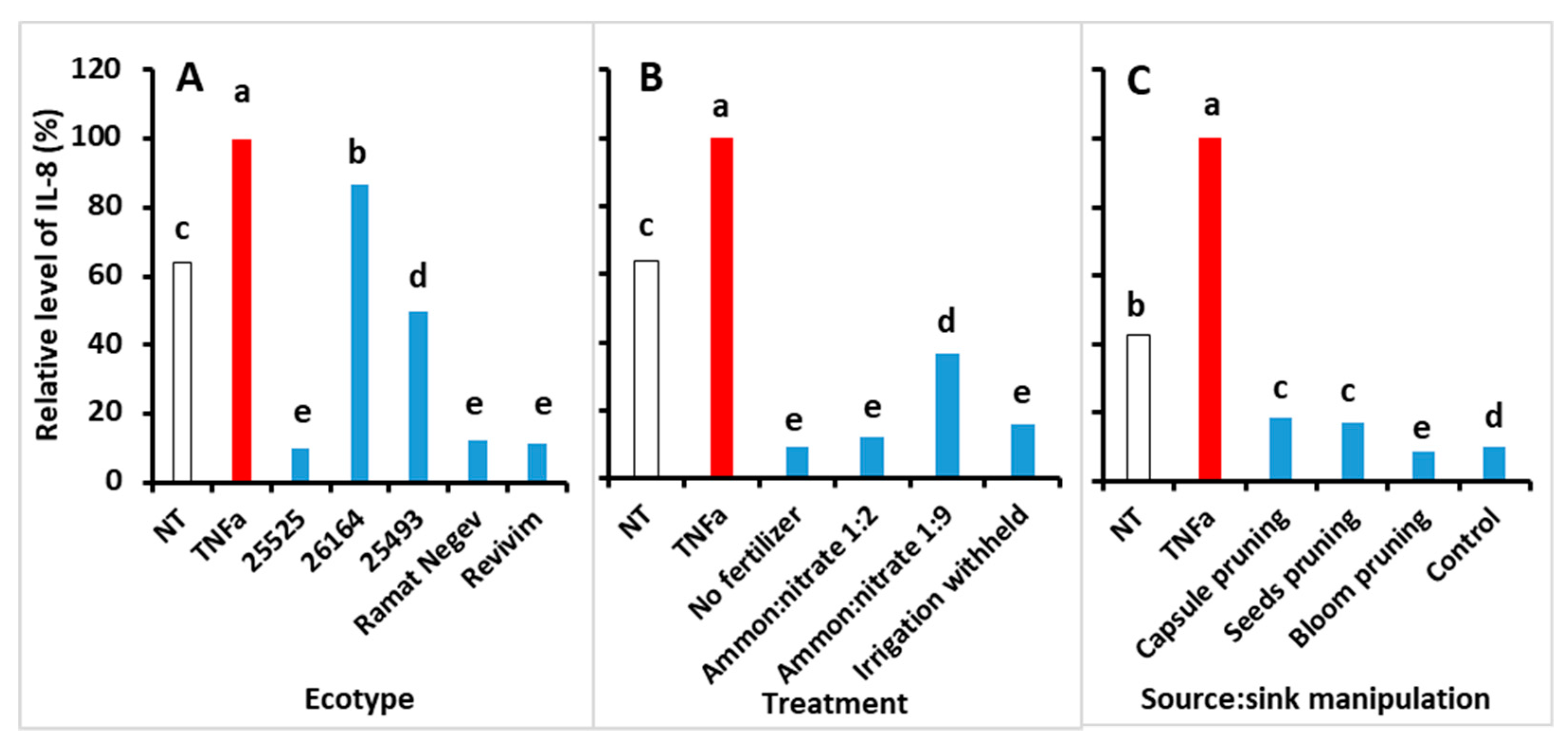
3.5. Anti-Inflammatory Activity
3.6. Challenges and Solutions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brullo, S.; Giusso del Galdo, G.; Guarino, R. Phytosociological notes on Lygeum Spartum grasslands from Crete. Lazaroa 2002, 23, 65–72. [Google Scholar]
- Bergmeier, E.; Kypriotakis, Z.; Jahn, R.; Böhling, N.; Dimopoulos, P.; Raus, T.; Tzanoudakis, D. Flora and phytogeographical significance of the islands Chrisi, Koufonisi and nearby islets (S Aegean, Greece). Willdenowia 2001, 31, 329–356. [Google Scholar] [CrossRef] [Green Version]
- El-Mokasabi, F.M. Floristic composition and traditional uses of plant species at Wadi Alkuf, Al-Jabal Al-Akhder, Libya. Am.-Eurasian J. Agric. Environ. Sci. 2014, 14, 685–697. [Google Scholar]
- Sharawy, S.M.; Badr, A. Systematic revision of Erodium species in Egypt as reflected by variation in morphological characters and seed protein electrophoretic profiles. Int. J. Bot. 2008, 4, 225–230. [Google Scholar] [CrossRef]
- Bidak, L.M.; Kamal, S.A.; Halmy, M.W.; Heneidy, S.Z. Goods and services provided by native plants in desert ecosystems: Examples from the northwestern coastal desert of Egypt. Glob. Ecol. Conserv. 2015, 3, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Wahab, R.H. Condition assessment of plant diversity of Gebel Maghara, North Sinai, Egypt. Catrina 2008, 3, 21–40. [Google Scholar]
- Hand, R. Supplementary notes to the flora of Cyprus VIII. Willdenowia 2015, 45, 245–259. [Google Scholar] [CrossRef] [Green Version]
- Danin, A. Desert Vegetation of Israel and Sinai; Cana Publishing House: Jerusalem, Israel, 1983. [Google Scholar]
- Osman, A.K.; Al-Ghamdi, F.; Bawadekji, A. Floristic diversity and vegetation analysis of Wadi Arar: A typical desert Wadi of the Northern Border region of Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 554–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.; Koltai, H.; Selvaraj, G.; Mazuz, M.; Segoli, M.; Bustan, A.; Guy, O. Nutritive and medicinal potential of tubers from cultivated Hairy stork’s-bill (Erodium crassifolium L’Hér). Plants 2020, 9, 1069. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H.; Kapulnik, Y.; Friedlander, M.; Mayzlish-Gati, E.; Nasser, A. Development of Therapeutic/Nutritional Products Based on Erodium Plant Extract. U.S. Patent 10,960,035, 30 March 2021. [Google Scholar]
- Yang, C.S.; Wang, H. Cancer preventive activities of tea catechins. Molecules 2016, 21, 1679. [Google Scholar] [CrossRef] [PubMed]
- Mangels, D.R.; Mohler, E.R., III. Catechins as potential mediators of cardiovascular health. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The role of catechins in cellular responses to oxidative stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing gallic acid for its broad-spectrum applications. Mini Rev. Med. Chem. 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparło, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Mazibuko-Mbeje, S.E. Inflammation and oxidative stress in an obese state and the protective effects of gallic acid. Nutrients 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, A.; Xiong, W.; Lin, H.; Xiao, W.; Huang, J.; Liu, Z. Catechins enhance skeletal muscle performance. Crit. Rev. Food Sci. Nutr. 2020, 60, 515–528. [Google Scholar] [CrossRef]
- Yadav, R.P.; Tarun, G. Versatility of turmeric: A review the golden spice of life. J. Pharmacogn. Phytochem. 2017, 6, 41–46. [Google Scholar]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Abbo, S.; Lev-Yadun, S.; Gopher, A. The ‘Human Mind’ as a common denominator in plant domestication. J. Exp. Bot. 2014, 65, 1917–1920. [Google Scholar] [CrossRef] [Green Version]
- Abbo, S.; Lev-Yadun, S.; Gopher, A. Plant domestication and crop evolution in the Near East: On events and processes. Crit. Rev. Plant Sci. 2012, 31, 241–257. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Zaady, E.; Shachak, M. Infiltration through three contrasting biological soil crusts in patterned landscapes in the Negev, Israel. Catena 2000, 40, 323–336. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Lazcano, C.; Christensen, T.H.; Muñoz, P.; Rieradevall, J.; Møller, J.; Antón, A.; Boldrin, A. Compost benefits for agriculture evaluated by life cycle assessment. A review. Agron. Sustain. Dev. 2013, 33, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Silbermann, A.; Zipilevich, Y. Irrigation and Fertilization of Spring Potatoes in the Negev; Publications of the Israeli Extension Services, MOAG: Volcani, Israel, 2003. (In Hebrew) [Google Scholar]
- Katan, J.; Gamliel, A. Soil solarization–30 years on: What lessons have been learned? In Recent Developments in Management of Plant Diseases; Springer: Dordrecht, The Netherlands, 2010; pp. 265–283. [Google Scholar]
- Gajanayake, B.; Reddy, K.R. Sweet potato responses to mid- and late-season soil moisture deficits. Crop. Sci. 2016, 56, 1865–1877. [Google Scholar] [CrossRef]
- Sonnewald, U.; Fernier, A.R. Next-Generation strategies for understanding and influencing source sink relations in crop plants. Curr. Opin. Plant Biol. 2018, 43, 63–70. [Google Scholar] [CrossRef]
- Kipanyula, M.J.; Etet, P.F.S.; Vecchio, L.; Farahna, M.; Nukenine, E.N.; Kamdje, A.H.N. Signaling pathways bridging microbial-triggered inflammation and cancer. Cell. Signal. 2013, 25, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Omar, Z.Z. First description of White Grub Beetle, Maladera Insanabilis Brenske, 1984 (Coleoptera Melolonthidae: Melolonthinae) from Erbil Governorate, Kurdistan Region-Iraq. Plant Arch. 2019, 19, 3991–3994. [Google Scholar]
- Ahrens, D.; Arnone, M.; Massa, B. Maladera insanabilis (Brenske, 1894), specie invasiva nella regione Mediterraneae sua distribuzione in Libia (Coleoptera scarabaeidae Sericini). Nat. Sicil. S. 2006, 5, 349–357. [Google Scholar]
- Pathania, M.; Chandel, R.S.; Verma, K.S.; Mehta, P.K. Diversity and population dynamics of phytophagous scarabaeid beetle (Coleoptera: Scarabaeoidae) in different landscape of Himachal Pradesh, India. Arthropods 2015, 4, 46–68. [Google Scholar]
- Baloda, A.S.; Jakhar, B.L.; Saini, K.K.; Yadav, T. Efficacy of insecticides as standing crop treatment against white grubs in groundnut crop. J. Entomol. Zool. Stud. 2021, 9, 973–975. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Season | Month | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| October | November | December | January | February | March | April | May | |||
| Minimum temperature (°C) | 2009/10 | 15.7 | 9.1 | 7.3 | 7.0 | 8.7 | 10.4 | 10.5 | 12.7 | |
| 2010/11 | 15.6 | 11.2 | 6.1 | 4.3 | 7.0 | 5.6 | 9.8 | 13.2 | ||
| 2011/12 | 12.0 | 5.0 | 2.7 | 1.8 | 4.7 | 6.1 | 9.6 | 13.4 | ||
| 2012/13 | 14.2 | 11.4 | 5.6 | 5.4 | 7.0 | 9.1 | 10.5 | 14.6 | ||
| 2013/14 | 10.2 | 10.8 | 5.2 | 4.6 | 4.2 | 9.2 | 10.8 | 14.7 | ||
| 2014/15 | 12.4 | 10.3 | 6.6 | 4.7 | 5.2 | 8.2 | 9.0 | 13.7 | ||
| 2015/16 | 16.2 | 12.1 | 3.7 | 3.8 | 5.9 | 8.7 | 11.5 | 14.4 | ||
| 2016/17 | 13.6 | 10.3 | 3.9 | 2.4 | 3.2 | 7.3 | 9.5 | 12.8 | ||
| 2017/18 | 12.4 | 7.7 | 6.8 | 5.7 | 8.2 | 8.4 | 10.7 | 17.0 | ||
| 2018/19 | 16.0 | 10.3 | 6.5 | 3.0 | 5.4 | 6.2 | 8.9 | 13.4 | ||
| 2019/20 | 15.0 | 10.4 | 5.1 | 4.4 | 6.2 | 7.5 | 10.4 | 13.7 | ||
| 2020/21 | 16.3 | 10.1 | 7.0 | 5.7 | 6.8 | 7.0 | 9.7 | 14.4 | ||
| Mean | 14.1 | 9.9 | 5.5 | 4.4 | 6.0 | 7.8 | 12.7 | 14.0 | ||
| SD | 2.00 | 1.90 | 1.47 | 1.49 | 1.61 | 1.45 | 0.78 | 1.16 | ||
| Maximum temperature (°C) | 2009/10 | 32.1 | 23.7 | 20.8 | 21.5 | 22.4 | 25.9 | 28.4 | 32.1 | |
| 2010/11 | 32.8 | 29.6 | 22.5 | 19.2 | 19.9 | 23.0 | 26.9 | 30.8 | ||
| 2011/12 | 29.1 | 21.8 | 19.8 | 16.5 | 18.5 | 21.0 | 29.2 | 32.0 | ||
| 2012/13 | 31.1 | 25.5 | 20.4 | 18.1 | 20.6 | 26.3 | 27.4 | 33.2 | ||
| 2013/14 | 28.8 | 26.7 | 18.7 | 19.4 | 21.1 | 24.1 | 29.6 | 31.5 | ||
| 2014/15 | 29.3 | 23.6 | 21.9 | 17.2 | 19.2 | 23.7 | 27.0 | 32.8 | ||
| 2015/16 | 30.9 | 24.6 | 18.7 | 17.0 | 22.7 | 24.8 | 31.3 | 32.4 | ||
| 2016/17 | 31.5 | 25.3 | 17.8 | 17.4 | 18.6 | 24.1 | 28.6 | 32.9 | ||
| 2017/18 | 29.4 | 24.5 | 22.2 | 17.5 | 23.3 | 28.0 | 29.0 | 34.4 | ||
| 2018/19 | 30.5 | 24.4 | 19.4 | 18.6 | 19.4 | 21.0 | 26.5 | 34.7 | ||
| 2019/20 | 30.9 | 26.9 | 20.3 | 16.3 | 18.7 | 22.8 | 27.5 | 33.3 | ||
| 2020/21 | 32.4 | 24.5 | 22.2 | 19.7 | 21.7 | 23.7 | 29.6 | 34.5 | ||
| Mean | 30.7 | 25.1 | 20.4 | 18.2 | 20.5 | 24.0 | 28.4 | 32.9 | ||
| SD | 1.35 | 1.97 | 1.57 | 1.53 | 1.71 | 2.05 | 1.41 | 1.22 | ||
| Sum | ||||||||||
| Precipitation (mm month−1) | 2009/10 | 32.7 | 4.5 | 16.7 | 59.7 | 30.3 | 23.0 | 0.2 | 1.2 | 168.3 |
| 2010/11 | 0.0 | 0.9 | 2.8 | 5.5 | 12.7 | 2.3 | 3.1 | 1.3 | 28.6 | |
| 2011/12 | 0.4 | 5.6 | 0.0 | 17.2 | 8.4 | 27.6 | 0.0 | 0.1 | 59.3 | |
| 2012/13 | 0.0 | 17.9 | 4.2 | 80.1 | 15.2 | 0.1 | 0.0 | 10.3 | 127.8 | |
| 2013/14 | 10.6 | 2.3 | 25.8 | 4.6 | 4.1 | 47.1 | 0.1 | 12.1 | 106.7 | |
| 2014/15 | 6.1 | 12.6 | 15.4 | 36.1 | 49.3 | 10.6 | 10.8 | 0.0 | 140.9 | |
| 2015/16 | 19.2 | 21.8 | 12.5 | 16.4 | 21.0 | 4.1 | 23.7 | 0.0 | 118.7 | |
| 2016/17 | 6.2 | 0.4 | 20.6 | 8.1 | 17.3 | 1.2 | 2.3 | 0.0 | 56.1 | |
| 2017/18 | 0.1 | 4.8 | 3.7 | 54.6 | 26.3 | 0.5 | 1.7 | 0.3 | 92.0 | |
| 2018/19 | 0.5 | 42.5 | 4.3 | 0.5 | 31.5 | 25.1 | 2.5 | 0.0 | 106.9 | |
| 2019/20 | 13.1 | 0.4 | 3.4 | 37.1 | 40.3 | 39.9 | 1.1 | 6.9 | 142.2 | |
| 2020/21 | 0.1 | 9.1 | 6.5 | 5.5 | 16.6 | 0.8 | 0.0 | 0.0 | 38.6 | |
| Mean | 7.4 | 10.2 | 9.7 | 27.1 | 22.8 | 15.2 | 3.8 | 2.7 | 98.8 | |
| SD | 10.2 | 12.3 | 8.3 | 26.2 | 13.3 | 16.8 | 6.9 | 4.4 | 44.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guy, O.; Cohen, S.; Koltai, H.; Mazuz, M.; Segoli, M.; Bustan, A. First Steps to Domesticate Hairy Stork’s-Bill (Erodium crassifolium) as a Commercial Pharmaceutical Crop for Arid Regions. Agronomy 2021, 11, 1715. https://doi.org/10.3390/agronomy11091715
Guy O, Cohen S, Koltai H, Mazuz M, Segoli M, Bustan A. First Steps to Domesticate Hairy Stork’s-Bill (Erodium crassifolium) as a Commercial Pharmaceutical Crop for Arid Regions. Agronomy. 2021; 11(9):1715. https://doi.org/10.3390/agronomy11091715
Chicago/Turabian StyleGuy, Ofer, Shabtai Cohen, Hinanit Koltai, Moran Mazuz, Moran Segoli, and Amnon Bustan. 2021. "First Steps to Domesticate Hairy Stork’s-Bill (Erodium crassifolium) as a Commercial Pharmaceutical Crop for Arid Regions" Agronomy 11, no. 9: 1715. https://doi.org/10.3390/agronomy11091715
APA StyleGuy, O., Cohen, S., Koltai, H., Mazuz, M., Segoli, M., & Bustan, A. (2021). First Steps to Domesticate Hairy Stork’s-Bill (Erodium crassifolium) as a Commercial Pharmaceutical Crop for Arid Regions. Agronomy, 11(9), 1715. https://doi.org/10.3390/agronomy11091715