1. Introduction
Within North America, the geographic range of native
Vaccinium species ranges from Canada through Mexico, encompassing a broad spectrum of different climatic growing conditions. In the southeastern United States, select native
Vaccinium species including
V. angustifolium Aiton,
V. arboreum Marsh,
V. constablaei Gray,
V. corymbosum L.,
V. darrowii Camp,
V. elliottii Chapm.,
V. myrtilloides Michx.,
V. pallidum Aiton,
V. tenellum Aiton,
V. simulatum Small,
V. stamineum L., and
V. virgatum Aiton have been utilized in breeding populations for specific adaptations and trait introgression. Introgression of native species into cultivated germplasm has adapted
Vaccinium germplasm to a range of climates and growing conditions for cultivated blueberry, resulting in both low-chill and high-chill adapted cultivars [
1,
2]). Breeding for low-chill and high-chill cultivation has enabled blueberry production expansion, increasing the geographic range and facilitating fresh market availability year-round due to longer production windows across regions. Furthermore, blueberry production has gained traction due to increased consumer and commercial attention due to recent reports regarding the health benefits of blueberry total anthocyanin content and anthocyanin composition, including aglycone, sugar moiety, and acyl group [
3].
Genetic differences signify phenotypic variance and are observed as fruit flavor or size differences between different blueberry accessions. Variability in geographic and climatic growing conditions between environments may further affect fruit and plant phenotypic expression. A comparative study of geographically diverse locations found that blueberry cv. ‘Brightwell’ (
V. virgatum) grown in higher altitude environments reported increased total soluble solids (TSS), flavonoids, phenols, proanthocyanidins, and anthocyanins in China [
4]. In addition to altitude, environmental differences encompass climatic factors, including temperature and precipitation. Increased TSS can be attributed to decreased fruit volume and moisture loss resulting from reduced rainfall events [
5]. Additionally, decreased precipitation during blueberry fruit ripening resulted in small, firm fruit, which correlated with increased soluble solids and acidity [
6]. Water deficit treatments also decreased fruit antioxidant levels in blueberry [
7]. Alternatively, increased TSS and decreased fruit weight resulted from warmer temperatures in tomato [
8]. In addition, temperature also plays a role in anthocyanin content. Genetic control of apple anthocyanin biosynthesis was found to be regulated by temperature [
9]. Hot climates decreased the expression of anthocyanin biosynthesis genes, which failed to accumulate red fruit pigmentation in apples grown in hot climates compared to those grown under temperate climate conditions [
9].
Phenotypic expression and observed variation of plant growth and development are functions of both genetic background (G) and environment (E) and their interaction (G × E) [
10]. Temporal variation can also impact plant and fruit quality phenotypic variation. Temporal regulation includes the effects of the year (Y) and harvest time (H), wherein biotic and abiotic events between years or maturation time between harvests are involved in fruit production and quality differences. Seasonal temperature differences over the years can significantly affect the fruit production of blueberries [
11,
12,
13]. Winter temperatures control dormancy release. Thus, the earlier onset of warmer temperatures exemplified by the three-week earlier snowmelt in
Vaccinium myrtillus in Norway in 2011 resulted in two-week earlier blooming compared with the 2010 production season [
12]. Furthermore, severe weather events such as spring cold spells can cause significant cold injury to low-chill and precocious blueberry cultivars, impacting yield [
14]. In addition to cold-related events, yearly differences in temperature fluctuations and precipitation events, including heatwaves, drought, and flooding, affect developing fruits’ quality [
15,
16]. Within a harvest season, weekly climate changes and increased growing degree days (GDD) can impact blueberry fruit maturation rates between sequential harvests within the same plant; exemplified by the and cause observed variation in phenolic, acid, and total soluble solid composition between harvests [
17]. Within a harvest season, weekly climate changes and increased growing degree days (GDD) can impact blueberry fruit maturation rates between sequential harvests within the same plant; exemplified by the and cause observed variation in phenolic, acid, and total soluble solid composition between harvests [
17]. By studying the impact of these interactions on phenotypic traits, we can evaluate factors affecting consistency in genotype performance.
In studying the effects of environment, years, harvests, and genotype on phenotypic traits, our research aimed to assess genotypic performance in select environments and evaluate phenotypic consistency. The cultivars used in this study are highbush blueberry ‘Echota’, ‘O’Neal’, ‘Reveille’, ‘Summit’, and ‘Sunrise’ (
Supplementary Figure S1). The southern highbush (SHB) cultivars ‘O’Neal’ and ‘Summit’ have
V. darrowii in their pedigrees; ‘Summit’ and ‘Reveille’ also have low-chill
V. virgatum introgression; ‘Reveille’ has
V. tenellum in its pedigree history. ‘Echota’ is a high-chill northern highbush (NHB) derived from a native low-chill North Carolina
V. corymbosum accession (NC102). ‘Sunrise’ is a modern northern highbush cultivar developed from high-chill
V. corymbosum parents with pedigrees containing
V. angustifolium; as such, ‘Sunrise’ does not have low-chill species in its pedigree history. Using these selected cultivars in both North Carolina and Oregon environments, our research informs on the stability of fourteen phenotypic traits. The phenotypic stability of a trait across environments, years, and harvests indicates a greater emphasis on the genetic control of the trait, which is essential for blueberry genotype uniformity for commercial production. The objectives of this study were to evaluate the genetic consistency of five blueberry cultivars and assess them for phenotypic variation between two environments, two years of research, two sequential harvests per year, and their interactions. Further, we aimed to evaluate the correlation between phenotypic traits. The results of this study may impact blueberry breeding program decision-making for target environments and further identify blueberry production benchmarks for specified traits.
4. Discussion
As blueberry production expands, it encompasses increased geographic and climatic diversity. Regions with compatible climates for both northern and southern highbush genotypes accommodate extended harvest windows. However, genotype response to diverse environments is largely uncharacterized. The main objective of the present study was to determine the extent of variation on fruit quality traits of five blueberry genotypes resulting from different sources of variation, including environment, year, harvest, and their interactions. Understanding the sources attributed to phenotypic plasticity informs that trait’s genetic control and stability [
10]. This research evaluates sources of variation significant to phenotypic stability and genotypic performance in select environments. Additionally, our analysis over multiple environments, years, and harvests was previously uncharacterized in blueberry for this range of phenotypic traits.
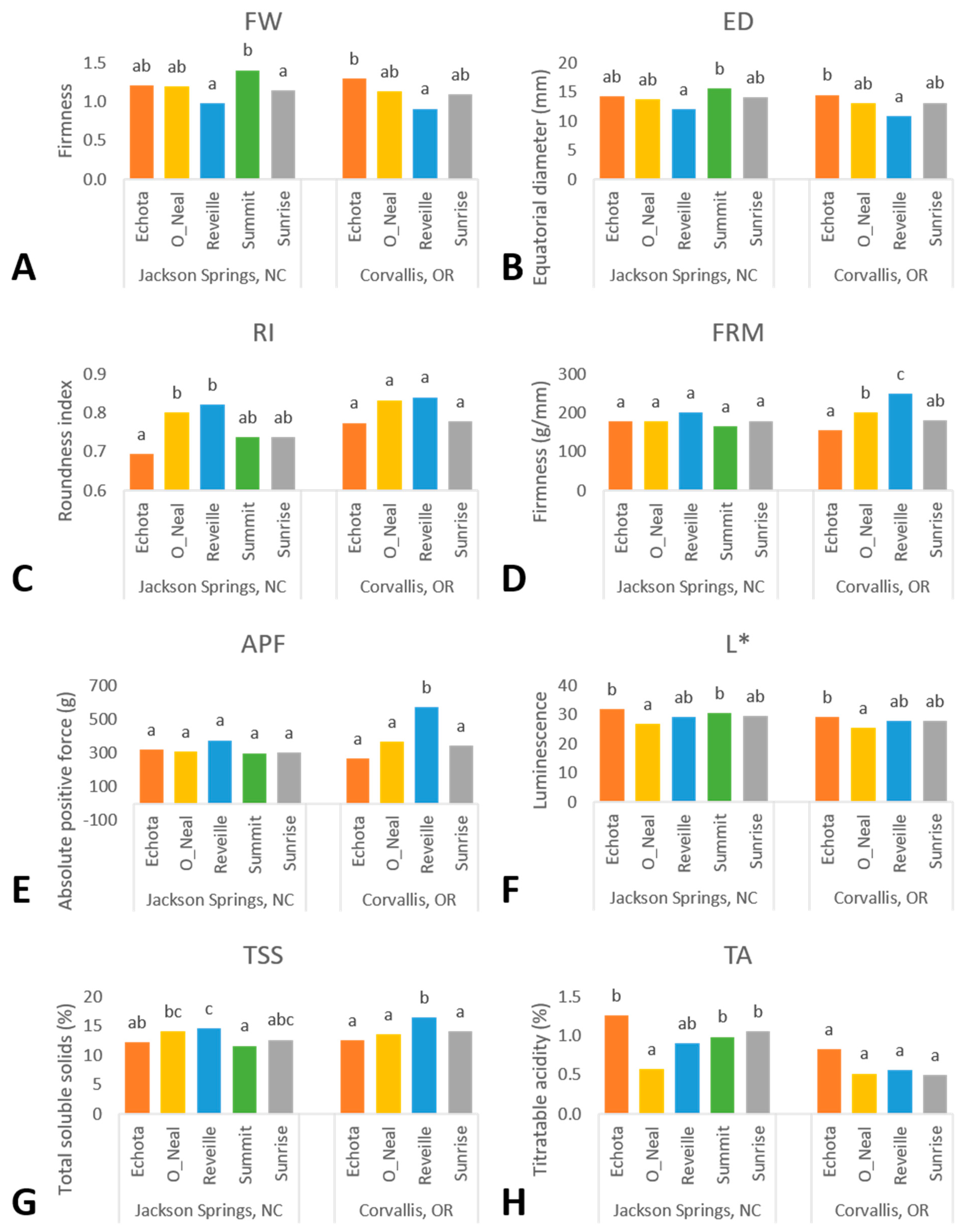
The selected environments saw differences in precipitation and air and soil temperatures. Corvallis had decreased net precipitation, air, and soil temperature, and increased evapotranspiration compared to the Jackson Springs environment in both years of study (
Table 8). However, our results indicated that E, G × E, G × E × Y, and G × E × H were not significant sources of variation for the majority of the phenotypic traits in either model. Yearly climatic differences in rainfall and temperature events suggested a greater influence on phenotypic expression. Precipitation and evapotranspiration are transitory effects that fluctuate year to year. Environmental differences are more evident in soil and air temperature and their effect on fruit maturation times. Air and soil temperatures warmed more gradually, and fruit matured later at the Corvallis location compared with Jackson Springs. While similar to Corvallis regarding maximum air and soil temperatures in March, the Jackson Springs environment experiences temperature increases in April that are not observed until May at the Corvallis location. The warmer air and soil temperatures at the Jackson Springs site agree with previous studies, which found that increased daytime soil and air temperatures advanced reproductive and vegetative growth of blueberry plants grown in high tunnels. Advancing the timeline of reproductive growth ultimately accelerated the fruit ripening by five weeks compared to blueberries under field conditions [
11]. Further, higher temperatures have been reported to induce pigment accumulation and accelerate ripening in blueberries [
25].
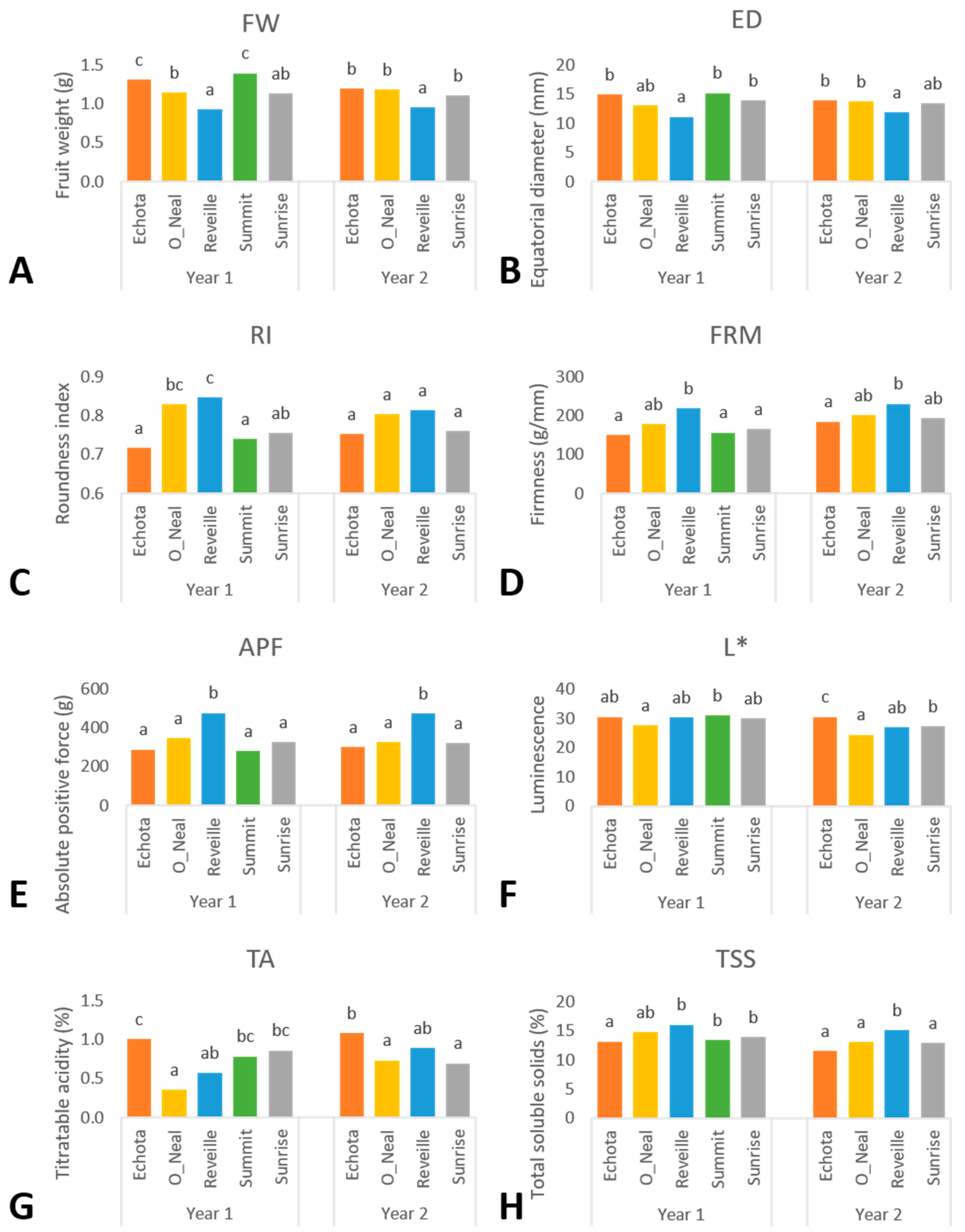
Yearly phenotypic variation is likely resultant from the differential precipitation and temperatures between years of analysis as observed within each environment—increased precipitation results in larger fruit in blueberry [
6]. Across environments, fruit size metrics were not significantly different; however, they varied in accordance with annual precipitation events within a location. Within the Corvallis environment, increased precipitation in 2019 from rainfall events resulted in larger fruit, as exemplified by the 14% increase in FW in 2019 compared to 2020, Where precipitation in the Jackson Springs environment decreased by 13% between 2019 and 2020, FW was affected by a 26% decrease. The soil type in Jackson Springs is sandy, and it was amended by pine bark to acidify the soil and increase organic matter. The plants were irrigated by a linear movement irrigation system every 7–10 days. However, the soil did not have much capacity to retain moisture even after incorporating pine bark. Further, PD, ED, RI, and DPF decreased in the Jackson Springs environment from 2019 to 2020 and increased in Corvallis. Fruit volume metrics are likely to increase due to precipitation events within a location [
6]. Conversely, water stress conditions, including high-temperature effects and drought conditions, negatively affect fruit volume metrics [
16,
26,
27]. Fruit volume metrics increased at the Jackson Springs and decreased at the Corvallis locations between the years of the study, which correspond to increased temperatures during harvest. While high-chill northern blueberry cultivars are negatively impacted at temperatures ≥ 30 °C, noted by a reduced photosynthetic rate [
28], low-chill-adapted blueberry cultivars have a higher estimated photosynthetic heat tolerance with no significant electrolytic leakage or superoxide radical accumulation observed at temperatures < 40 °C [
29]. While high-chill northern blueberry cultivars are negatively impacted at temperatures ≥ 30 °C, noted by a reduced photosynthetic rate [
28], low-chill-adapted blueberry cultivars have a higher estimated photosynthetic heat tolerance than northern high-chill-adapted cultivars with no significant electrolytic leakage or superoxide radical accumulation observed as a result of heat stress until at temperatures ≥ 40 °C [
29]. In our findings, the maximum temperatures during harvest (May–June at the Jackson Springs location and July–August at the Corvallis location) exceeded 40 °C at both locations in both years of study. Overall, fruit volume metrics were higher in years with a lower mean temperature during harvest. The decreased fruit volume metrics in the NHB cultivars ‘Echota’ and ‘Sunrise’ were related to increased annual temperatures between years for both environments; these observations may support decreased photosynthetic rates due to of heat stress. Coinciding with years of lower maximum air temperatures, ‘Sunrise’ and ‘Echota’ had increased fruit volume metrics in Corvallis in the first year and Jackson Springs in the second year of study. However, it is noteworthy that the increased fruit volume metrics at the Jackson Springs location were likely significantly influenced by the three-fold increase in precipitation in the second year of study. Further research examining the effects between NHB and optimal fruit production temperature would establish a baseline for comparisons for fruit volume metrics.
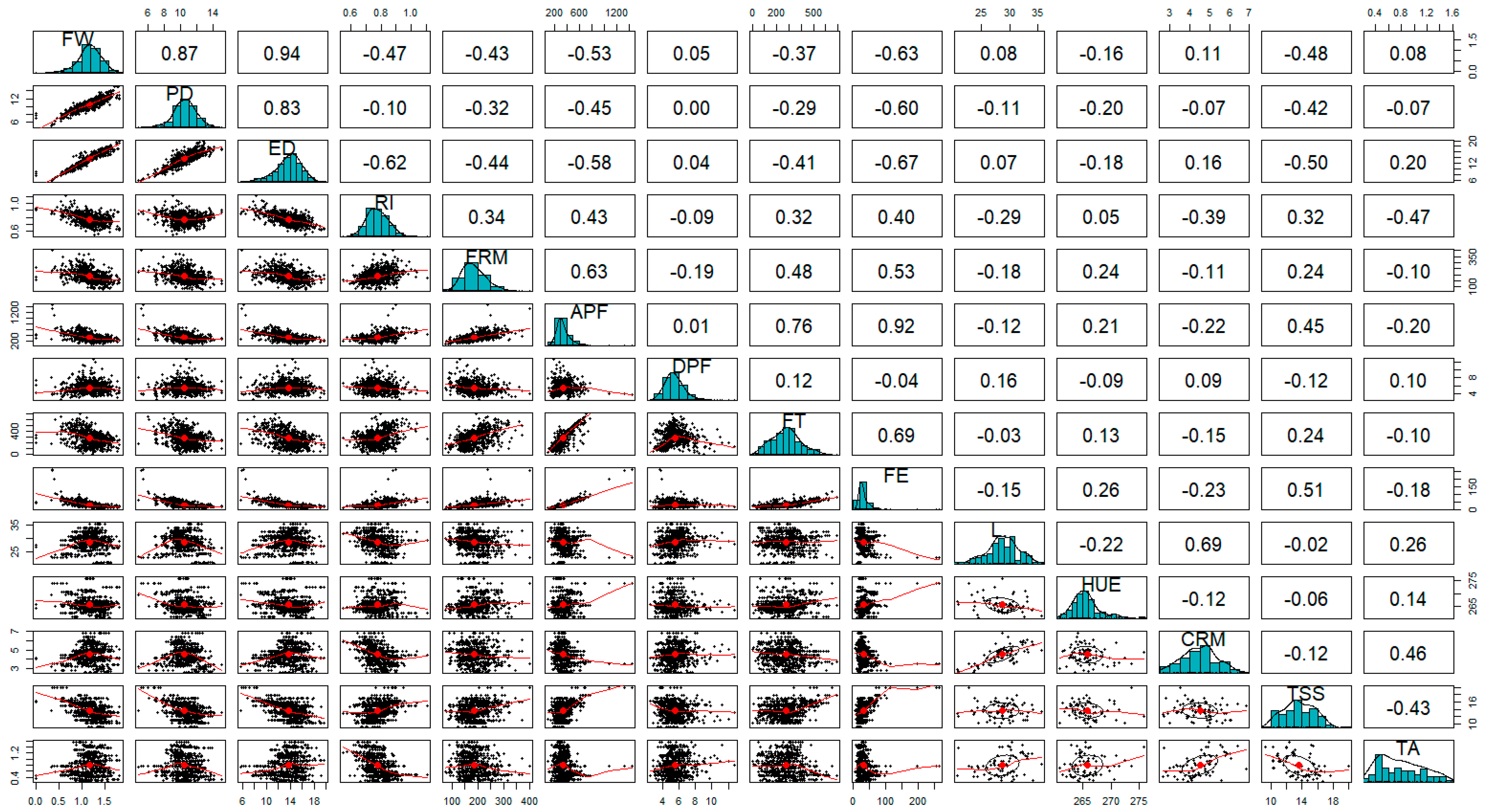
FW and ED had a moderate negative correlation with fruit firmness metrics, including FRM, APF, and FE (
r > 0.40). We observed decreases of 17%, 6%, and 15% in traits, respectively, from 2019 to 2020 in Corvallis. Conversely, fruit volume metrics of FW, PD, ED, and DPF increased 14%, 9%, 10%, and 10%, respectively, from 2019 to 2020 in Corvallis. Previous reports indicate that decreased precipitation corresponds to firmer fruit [
6]. While Corvallis plants are consistently irrigated twice a week, climate data reported minimal rainfall in July and August in both years of study and increased net evapotranspiration. Corresponding to increased evapotranspiration during harvest, FW, PD, and ED all decreased while FRM, APF, and FE all increased at Corvallis in 2020. At the Jackson Springs location, only APF and FE had increased values coinciding with increased evapotranspiration in 2019 compared with 2020. Contrasting with the expected increase in precipitation and reduced evapotranspiration in May and June 2020, FRM measurements were significantly higher in Jackson Springs. At the same time, FW also increased from 2019 to 2020, likely due to the three-fold increase in precipitation. Thus, both FW and FRM simultaneously increased in Jackson Springs between 2019 and 2020. FRM increased in ‘Echota’, ‘O’Neal’, and ‘Summit’, all of which increased FW. In contrast, ‘Reveille’ showed an increase in FW but a decrease in FRM. Only ‘Sunrise’ had a lower FW and higher FRM between years of the study in Jackson Springs. Contrary to environmental influences between the first and second years of study, ‘Echota’, ‘O’Neal’ and ‘Summit’ exhibited higher fruit volume-related metrics and lower FRM, APF, FT, and FE values. ‘Reveille’ reflected expectations of increased fruit volume metrics and decreased fruit firmness measurements at Jackson Springs from 2019 to 2020. The overall moderate negative correlation between FW-FRM and ED-FRM indicates that denser fruit with higher weight values and fruit with a larger equatorial diameter had lower fruit firmness. These correlations and an inverse relationship with precipitation and evapotranspiration suggest minimal temporal influence over these phenotypic traits.
While increased water uptake due to increased irrigation or precipitation in blueberry fruit positively correlates with fruit volume metrics, it negatively correlates with TSS [
5]. The Jackson Springs location faced lower precipitation in May in 2019 and higher rainfall in June than 2020. More importantly, evapotranspiration was nearly 40% and 6% greater in May and June 2019, respectively, compared with 2020 at Jackson Springs. Blueberry plant water loss through evaporation reduced fruit volume metrics, including FW, PD, and ED, correlated with increased TSS values (
r > 0.40). Corvallis experienced increased evaporation in 2020, which had decreased FW, PD, and ED and increased TSS.
Additionally, the increased air temperatures during harvest at each location may positively affect fruit TSS values. For example, at Jackson Springs, TSS values averaged across cultivars in 2019 were 21% greater than 2020, when May and June maximum air temperatures were 12.8% and 2.5% greater in 2019 than 2020. Comparatively, there was only a slight increase in both maximum air and soil temperature (<3% in July and August) and TSS (<2%) values in fruit collected from Corvallis in 2020. Corvallis blueberry harvest in July and August was hotter with cooler nighttime temperatures in 2020 compared with 2019; however, TSS showed a minimal increase. Contrary to our results, previous studies in blueberry report a negative correlation between decreasing light level or photosynthetic active radiation and canopy temperature with reduced fruit water content and fruit weight, while TSS increased [
30].
In contrast, another study found a higher air and soil temperatures due to growing blueberry plants in high tunnels had no reported effect on TSS compared with control blueberry plants in the field [
11]. These studies did not separate the light level and temperature treatment effects. Thus, conclusions may differ as there have not been studies that isolate the direct effect of air temperature and GDD without impacting light level and air movement on pre-harvest phenotypic characteristics in blueberry fruit. Similar to blueberry, red grapes are non-climacteric fruit that accumulates pigment during ripening. In grapes, high temperatures inhibit pigmentation and anthocyanin accumulation in select cultivars [
31]. In contrast to Jackson Springs blueberry observations, reduced temperatures in grape, with a mean temperature of 20.3 °C, reflected increased TSS values compared to a warmer (37 °C/32 °C day/night) temperature treatment [
32]. In a similar study, TSS was highest in interspecific wine grape hybrid held at a constant 25 °C for 15 to 37 days and lower at 20 °C and 30 °C, and TA was highest in fruit harvested from plants kept at 20 °C [
33]. A study on tomatoes also found that increasing temperature from 26° C to 32 °C corresponded with increased TSS values [
8].
Our results found a moderate negative correlation between TSS and TA. The relationship between decreased precipitation and fruit volume is positively correlated with TSS. However, we did not find any correlation between fruit volume metrics and TA. Interestingly while it has been shown that plants under decreased precipitation and increased water stress had higher TA and TSS [
5,
6], our results showed that there was a negative correlation between TSS and TA.
The HUE angle of blueberry defines the gradient of the blue color of the fruit on a color spectrum; values closer to 270° are considered darker blue, and values approaching 220° are lighter blue. CRM, on the other hand, indicates the level of color intensity or saturation. From 2019 to 2020, CRM values decreased in Jackson Springs and increased in Corvallis, coinciding with warmer harvest temperatures at each location. Chromaticity values have been related to pigment accumulation in fruit [
34]. Previous results found that post-harvest fruit and blueberry puree had higher chroma values at lower temperatures, agreeing with our findings at the Jackson Springs location; however, these results were not significant between 4 °C and room temperature, 75 °C, and 100 °C in a study of three NHB blueberry cultivars. Overall, L* and HUE decreased and increased, respectively, between years in our study and do not correspond to temperature or rainfall events in either environment. This observation was consistent across G × Y except for ‘Echota’, for which L* values slightly increased between years of study in Corvallis. Fruit color is a factor of fruit ripeness and maturation, which can be altered by harvest time and anthocyanin accumulation. As blueberry fruit ripens, anthocyanin content increases, turning from green to blue with intermediate red or blush hues. Further, the fruits of most blueberry cultivars produce a wax coating or bloom, which can preclude color determination. Fruit bloom affects L* in the light-dark color perception of the fruit and is impacted by fruit handling and weather, diminishing the bloom coating. As such, we would expect L* values to have increased variability.
As a whole, blueberry fruit ripening is indicated by fruit pigment accumulation for one month. Plant fruit ripening and the blueberry reproductive cycle are based upon genotype and environmental factors controlling budbreak. Fruit derived from the same plant but collected at a later harvest due to a later fruit maturation window will have accumulated more GDD and increased opportunities for rainfall events and temperature increases. As such, we would expect to see differences between harvests. Our results identified significant differences in both ‘O’Neal’ and ‘Reveille’ regarding fruit volume metrics between harvests, as well as differences in L*, HUE, CRM, and TSS in ‘Echota’, ‘Reveille’, and ‘Sunrise’ between harvests.
It was intriguing that we did not find any consistent effect between harvests within a genotype across environments or years for any phenotypic trait. However, we did observe that TSS increased between harvests for most genotypes on an environment and yearly basis. While previous findings reported blueberry fruit decreasing in CRM values and L* during ripening, our results did not find any consistent trend in either CRM or L* [
25]. However, a major difference between the previous study [
25] and ours is the evaluation of blueberry fruit ripening stages covering pigmentation. Our study looks at similarly ripe fruit and examines fruit variation between successive harvests. Variation in fruit measures within a plant between successive harvests signify differences in GDD and temperature events between harvest timepoints. As fruit matured and accumulated anthocyanins, Spinardi et al. (2019) found that fruit shifted in hue angle from 290° (violet) to 210° (blue) during ripening. In contrast, our findings did not see a shift in HUE between harvests, further supporting the uniform ripeness of fruit between harvests.
Intriguingly, we did not find any differences in fruit firmness metrics associated with either harvest timepoint. A previous study examined the metrics between blueberry fruit ripeness and canopy position found that fruit six days post-optimal-ripeness were significantly less firm than optimally-ripe fruit [
15]. While our study only evaluated ripe fruit, it is important to note that increased GDD did not impact fruit firmness metrics. While average harvest values of FRM and FT decreased between harvests, the difference was not significant. Further, fruit firmness metrics of APF and FE increased. Additional interactions with genotype, environment or year were not a significant source of variation with harvest effects for any measured fruit firmness metrics.
Genotype is an expected significant source of variation. Accordingly, all studied phenotypic traits displayed significant differences between genotypes apart from DPF, and HUE. Overall, ‘Reveille’ performed highest in fruit firmness metrics (FRM, APF, FT, and FE) across genotypes within either environment or year of study. ‘Summit’ consistently had higher fruit volume metric averages (FW, PD, and ED) across the evaluated genotypes in both the environments and years of the study. Similarly, ‘Echota’ had the second-highest values for fruit volume metrics, and, as with ‘Summit’, lower fruit firmness metrics were observed, including FRM, APF, PF, FT, and FE. ‘Echota’, followed by ‘Summit’, had the highest L* and CRM values rating across genotypes. Higher L* values signify lighter fruit skin coloration as a result of pigmentation or genotypic bloom production. The current market preference in blueberry visual appeal coincides with higher L* values and light blue blueberry fruit in addition to larger fruit size characteristics [
35] displayed by both ‘Echota’ and ‘Summit’. ‘O’Neal’ was the least acidic genotype in this study and had a rounder fruit similar to ‘Reveille’ yet had the lowest L* and CRM values. ‘O’Neal’ was average among the majority of studied traits, representing both medium volume and firmness. Similarly, ‘Sunrise’ represented the median value for nearly every phenotypic trait; this observation was consistent in both environments for all phenotypic traits except for DPF, wherein ‘Sunrise’ had the highest DPF value across genotypes in Corvallis.


 ,
, 




{kind=link}
{kind=link}
{kind=link}