1. Introduction
By increasing residential houses, due to the growing population in the developing countries, the human population pressure of water consumption on global freshwater resources will noticeably escalate. Almost half the human population on the globe do not have access to potable water [
1] and water scarcity for agricultural crops may be a critical issue in the twenty-first century [
2], as it is currently using 70% of the total available freshwater [
3,
4]. Furthermore, there is growing poverty and socio-economic issues in the developing countries where freshwater is crucial for food production to meet the quantitative and qualitative needs of the growing population [
5]. Therefore, it seems necessary to increase food production without a significant increase of freshwater availability and utilization. Most vegetables are sensitive to a saline environment, and yield decreases are noted with moderate to high levels of salt stress in the plant rooting zone [
6]. In order to minimize the reduction in crop yield, proper managing approaches are required to maintain the salt ions in the plant rooting zone below the critical level [
7]. Furthermore, in saline conditions, the quality and quantity of crops are unstable and can be maintained by minimizing the movement of salts into the plant growing media.
Malnutrition is one of the main problems of two-thirds of the population of the world, resulting in a shortage of one or more essential mineral elements in daily food [
8,
9]. Quality of food can be produced in the prevailing resources through proper management [
10]. Moreover, the concentration of these essential nutrients, i.e., vitamins and phytochemicals (polyphenol, carotenoids, phenolic acids, flavonoids, ascorbic acids, fibers, stilbenes/lignans) can be maintained in the food [
11]. Hence, agricultural products are the basic source of nutrients for human beings when introducing any new strategy for optimizing, improving, or sustaining crop production.
In general, the quantity and quality of vegetables are influenced by environmental stresses such as salinity and drought [
12]. Salt sensitive plants under a salinity-stressed environment often tend to reduce water consumption, and consequently, retard the nutrient absorption systems [
13,
14]. Most of the conventional crops are not tolerant to elevated salinity stress, despite this, several are growing in up to 1–30% seawater concentrations [
15]. Seawater may be the greatest alternative water source for irrigation as it is available on a large scale [
16] and also a rich source of most of the nutrients [
17] that are equally beneficial for plants and human. Thus, an integrated approach in the form of blending seawater with fresh water will be imperative for the sustainable advancement of the agricultural sector [
15]. Seawater application to growing crops has long been under investigation, but its pure application to agricultural crops is impossible and may be needed to check its feasibility in a closed environment (hydroponic crop), as it will not aggravate the salinization problem in the crop root zone, which is a major threat to its application to crops [
1]. Thus, seawater use can give us two benefits at the same time: the decreasing dependence of the agriculture sector on freshwater and growing crops utilizing the rich source of nutrients.
The earlier findings assumed that the application of seawater to crop plants up to a certain concentration does not affect crop quality and productivity [
3,
14,
18,
19,
20]. Moreover, the maximum threshold level of seawater may change according to the nature of growing plant species. The chloride concentration in the seawater did not influence the uptake of nitrogen in lettuce and potato crops [
2]. Moreover, the crop performance and higher N uptake significantly reduced the uptake of chloride concentration [
1]. In addition, past studies have indicated that the concentration of secondary metabolites and the organoleptic value of vegetables are better in a saline environment (seawater or NaCl) as seen in broccoli [
21], onion [
22], tomato [
23], and lettuce [
3,
20]. Those that are organoleptic with higher nutritional values are used in the prevention of human disease [
24]. Thus, the achievable growth and development of other vegetables should be managed by adding a certain level of salt concentration.
All these results regarding the use of seawater on vegetables recommend that the use of the diluted seawater may improve the bio-fortification of horticultural crops and enhance the nutrients status of the food [
2]. Although the previous literature offers the effects of a saline environment on over 130 crop species, information about several other crops with the use of seawater are missing [
25], especially focusing on the nutritional and quality aspects.
In the present study, an experiment was planned to examine the effects of different seawater treatments (seawater and freshwater blend) on the three most consumable summer horticultural crops: eggplant (Solanum melongena L. CV. PP long), pepper (Capsicum annuum L. CV. NARC-4) and tomato (Solanum lycopersicum L. CV. Rio grandi). The present study explored the effects of (i) growing of eggplant, tomato and pepper using seawater and freshwater blend, and (ii) seawater in the growing media on crop productivity, green pigments, gas exchange parameters, mineral elements, quality, MDA, ASA and phenols.
2. Materials and Methods
2.1. Site Description
The study was conducted at the Department of Environmental Sciences, COMSATS University Islamabad, Vehari Campus, Vehari Punjab, Pakistan (30.0318° N, 72.3145° E) during the autumn season (2019) to study the effect of different Indian seawater concentrations on the three commonly used summer vegetables. The plant analyses were performed at the postgraduate laboratory of the department.
2.2. Plant Material, Experimental Design and Growth Conditions
The three most common commercial subtropical vegetables were selected for the present study; eggplant (
Solanum melongena L. CV. PP long), pepper (
Capsicum annuum L. CV. NARC-4) and tomato (
Solanum lycopersicum L. CV. Rio grandi). The consumption of these crops is increased in Pakistan during the summer season. The experiment was performed in half-strength hydroponic culture as explained by Hoagland and Arnon [
26]. The crop cycle length (sowing and harvesting) was managed as was routinely performed in soil cultivation. All the chemicals for hoagland solution were taken from Sigma-Aldrich, St. Louis, MO, USA distributer, Punjab, Pakistan.
The nursery of each vegetable was sown on 1 February 2019 at the experimental site of Department of Environmental Sciences, COMSATS University Islamabad, Vehari Campus, Vehari, Punjab, Pakistan and transplanted on 10 March 2019. The crop nursery for each vegetable (40-day-old seedlings) were transplanted into hydroponic culture on the same day at morning time. For acclimatization of newly transplanted plants, the plants were grown for 10 days after the transplantation in the nutrient culture without seawater treatments. If any plant was dead, then it was replaced from the extra plant stock. During the experiment, plants were grown in normal humidity (average 40%–50%), natural light intensity (280 watt/m; without artificial light) and temperature (30 °C/18 °C day/night). The experimental setup consisted of 26 plastic tubs (V. 5 L each) having 4 plants in each and total number of plants were 16 of each vegetable in each replication (total 48 plants per replication). All the hydroponic units were continuously aerated with the help of small aquarium pumps that were purchased from the local market of Burewala, Punjab, Pakistan.
The seawater applied to the experimental plants was collected from the Indian Ocean (approximately 5 Km away from Port Bin Qasim Karachi, Pakistan) 10 days before the initiation of trial, and stored in a sterile tank at ≈ 35–40 °C. Chemical characteristics of the used seawater are presented in
Table 1. Na
+ and K
+ concentrations were measured with the help of flame photometer (Perkin Elmer; Singapore). The pH and EC were measured by using a portable pH meter (Hanna Instruments, China). NO
2, NO
3, phosphate and silicate were determined with a spectrophotometer (Perkin Elmer; Singapore). Four different seawater treatments were applied to the plants, i.e., CT (control; distilled water along with nutrient solution); A: (5% seawater + nutrient solution); B: (10% seawater + nutrient solution); C: (20% seawater + nutrient solution). Moreover, the pH and EC of the growing media was measured on a daily basis using the portable pH meter and maintained the pH 6.0 ± 0.05 during the entire period of the experiment. The hydroponic media was replaced after every two weeks, such that the level of nutrients was maintained during the whole duration of experiment. The physicochemical characteristics of the hydroponic media (nutrients solution+ seawater concentrations) are shown in
Table 2.
2.3. Plant Growth, Biomass, Water Use Efficiency and Water Potential Analysis
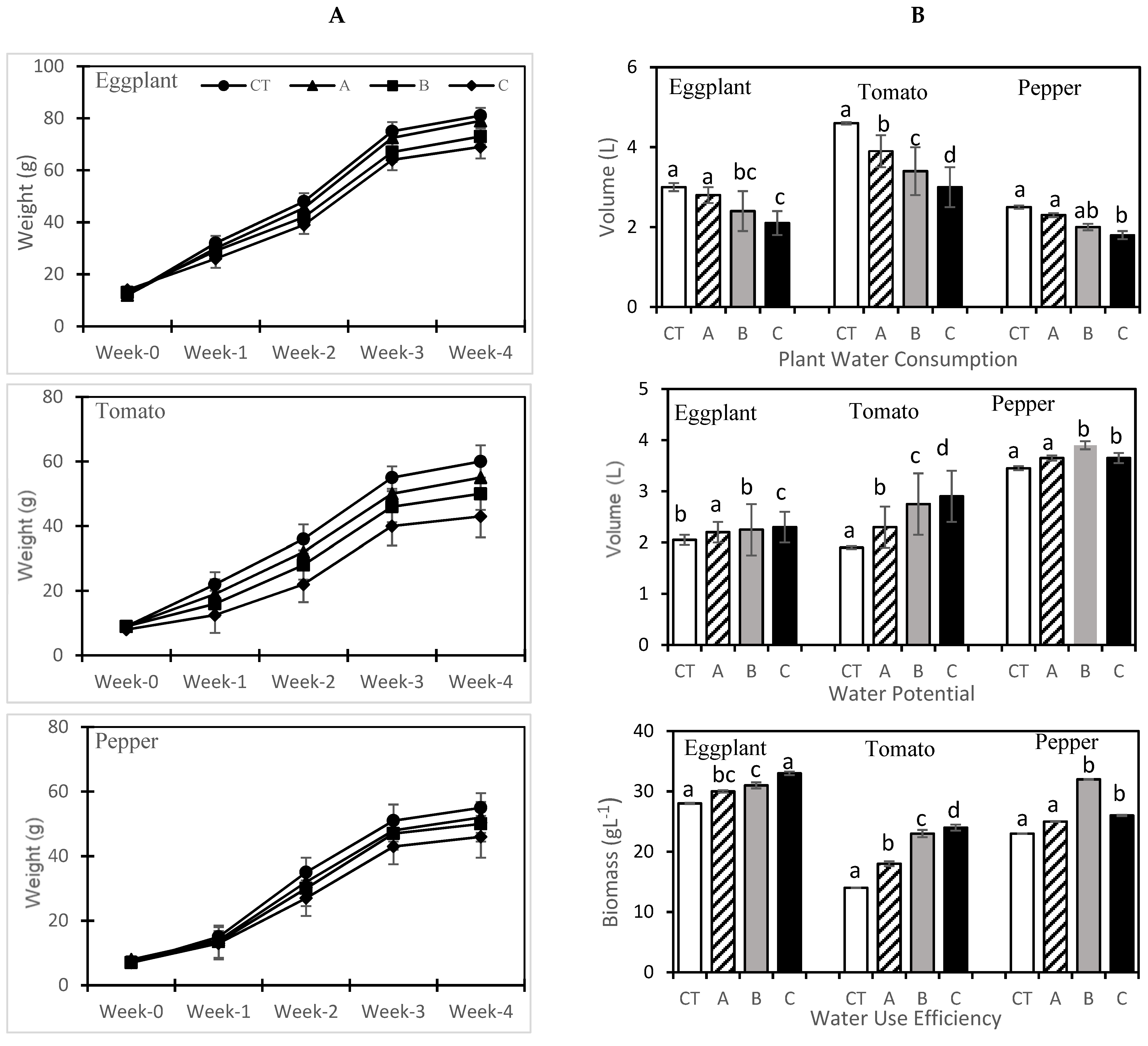
Growth characteristics of each crop was measured weekly by weighing the entire pot including the plant by subtracting the pot weight and nutrient media, thus obtaining the entire plant weight (g). At the harvesting time, fresh leaf samples of each crop were collected from all four replicates per treatment, frozen into the liquid nitrogen at the greenhouse, and stored into freezer at the laboratory for further analysis i.e., pigment analysis, antioxidant activities and enzymatic studies. Subsequently, shoot fresh weight of all crop plants were taken individually by cutting the root portion from the main plant body. The entire plant was placed into a warm air oven at 70 ± 2 °C (Wise Ven, Wisd Laboratory Instruments, Korea) until constant weight was achieved, and thus the whole plant dry mass was recorded.
Crop evapotranspiration (ET) was determined by weighing the solution volume of each treatment (weekly basis). We have assumed zero water loss apart from the plant body (plant evapotranspiration) as the experiment was a closed system with a thermopore sheet from the corner of pots (pots from corner side were tightly packed with thermopore sheet). The WU was calculated by using the following equation: WUE = whole plant dry biomass yield (g)/ET (L). WP was calculated by the following equation: WP = fresh marketable biomass (g)/ET(L). The marketable yield (g/plant) was noted by weighing the fruits yield of each vegetable at harvesting.
2.4. Determination of Gas-Exchange Parameters
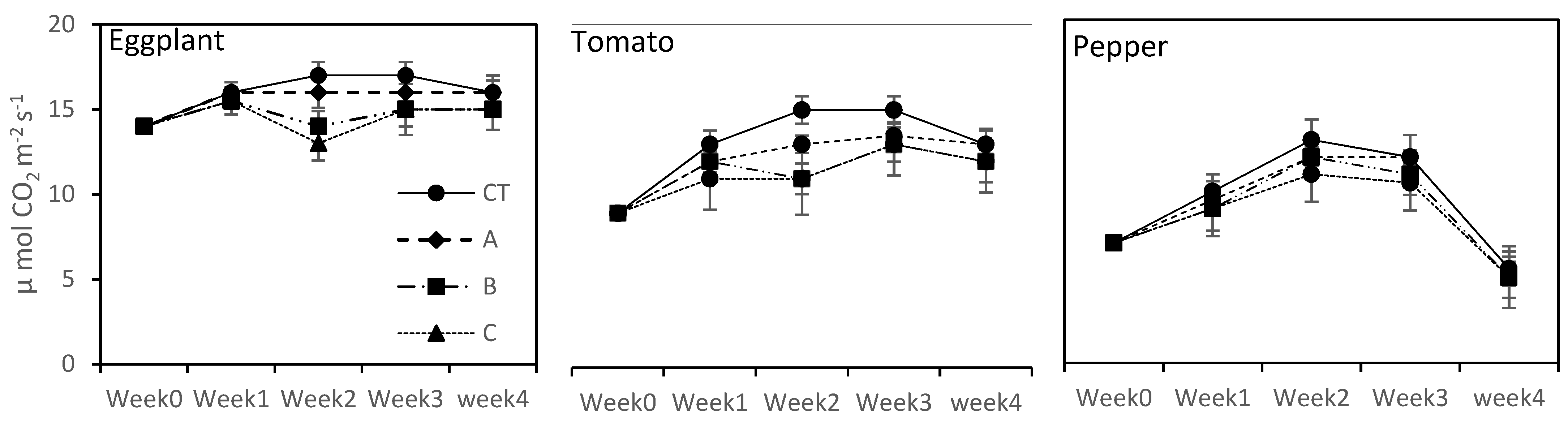
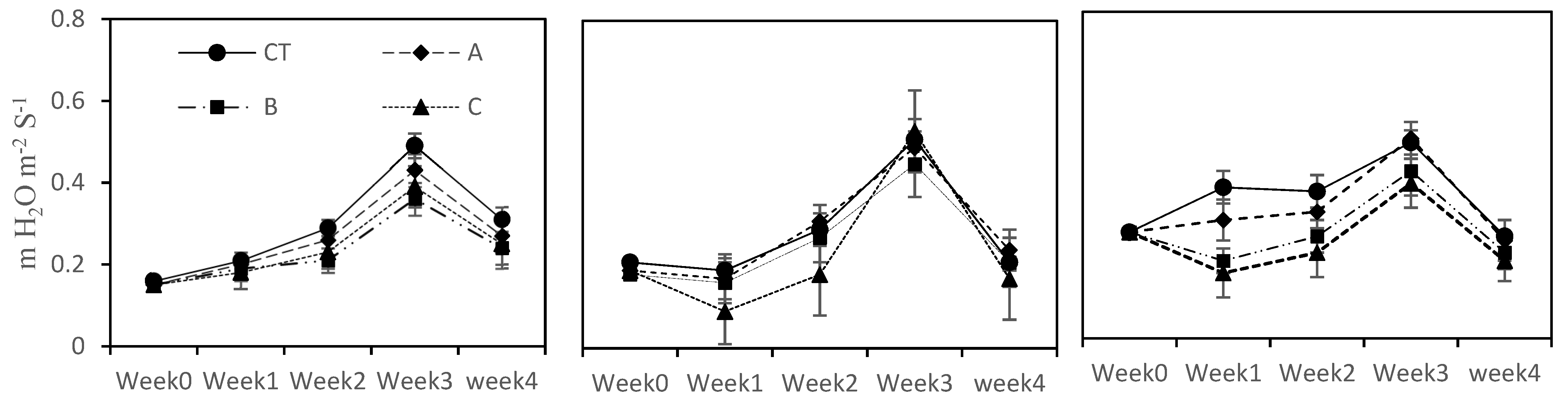
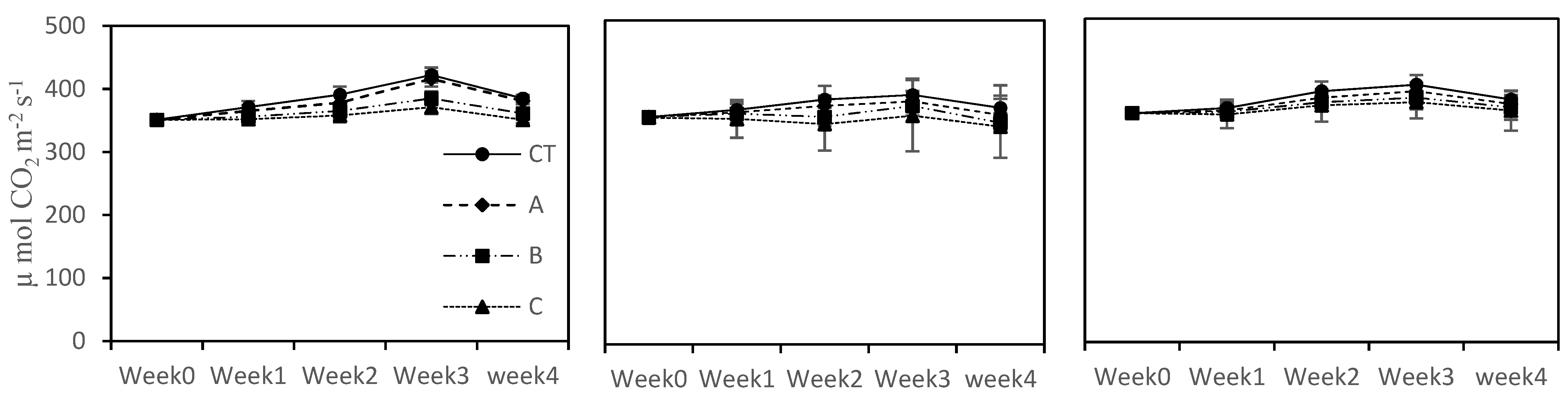
Plant leaf gas exchange parameters, i.e., photosynthetic rate (Pn), transpiration rate (E) and internal rate CO2 (Ci), were determined by using IRGA (infrared gas analyzer). These measurements were taken from each plant of each treatment in each crop. For this purpose, the second youngest lush green leave (upper surface of leave) was taken and was placed on the sensor for stable readings. The gas exchange parameters were taken when each vegetable completed its vegetative stage. All the readings were taken on a clear sunny day with ambient humidity and temperature.
Photosynthetic pigments, i.e., chlorophyll a (chla), chlorophyll b (chlb), carotenoid (car) contents and total free porphyrins, were measured from each treatment of all crops. Total porphyrin content was the sum of protophyrin, Mg- protophyrin and proto-chlorophyllide. The plant leaf extract (homogenized with 90% acetone (v/v)) was centrifuged for 5 min and absorbance of the extract was recorded at 663, 648 and 470 nm, while case of total porphyrins was determined at 575, 590 and 690 nm using a spectrophotometer (Perkin Elmer Lambda 25, Singapore). Total pigment concentration was determined by following the methods of Lichtenthaler [
27] and Chi-Ming Yang [
28] explained below:
Chlorophyll a (mg/g FW) = (11.75 × A663 − 2.35 × A645) × 50/500;
Chlorophyll b (mg/g FW) = (18.61 × A645 − 3.96 × A663) × 50/500;
Carotenoid (mg/g FW) = ((1000 × A470) − (2.27 × Chl a) − (81.4 × Chl b)/227) × 50/500.
2.5. Determination of Fruit Quality Parameters
Titratable acidity was measured by following the titrimetric method and was expressed in term of citric acid (%). Total sugar contents in the fruits were measured using the Luff-Schoorl method [
29]. The plant protein contents were determined by measuring N concentration and protein was found by multiplying N with 6.25 factor [
30]. For the analysis of vitamin C, the titration method was used [
31]. Nitrate was determined by measuring the wavelength of plant extract at 410 nm using the spectrophotometer [
32]. Total oxalate was measured by following the method of Baker [
33].
2.6. Determination of Mineral Elements, ASA and MDA Contents
The concentrations of Na
+, K
+ and Ca
+2 were determined by digesting the plant leaf material (0.1 g per treatment of each vegetable) in the presence of HNO
3 (5 mL; 67%) and deionized water (5 mL). After mineralization of the sample, the volume of the extract was increased up to 25 mL by adding more deionized water. The concentration of Na
+, K
+ and Ca
+2 was measured by running the extract on flame photometer (Perkin Elmer Lambda 25, Singapore). The standards were run before the start of Na
+, K
+ and Ca
+2 determination. The concentrations of Fe, Mg
+2, Zn
+2 and Cu
+2 were measured by following the method of Pignatelli et al. [
34].
Ascorbate peroxidase activity (ASA) was calculated by using the method by Amako et al. [
35]. The proline content analysis was performed from each vegetable leaf. The lipid peroxidation was recorded in term of malondialdehyde (MDA) contents, and readings were taken at 532 and 600 nm. The MDA concentrations were calculated by following the equation developed by Heath and Packer [
36]:
2.7. Statistical Analysis
Four replicates of each treatment were used for all vegetables. The completely randomized design (CRD) was used to design the experiment. The means and standard errors (shown in graphs) were calculated using Excel (Microsoft Office 2007). Analysis of variance was performed using Minitab’s ANOVA and General Linear Model GLM (version 15). The graphs are constructed by using the Origin Pro software (V. 2017).
4. Discussion
The present research findings indicate that all three vegetables reduced their biomass yield at C (20% SFW) treatment but showed a nonsignificant difference at control and A (5% SFW) treatments when measured at the time of harvesting. These results are in agreement with those of Atzori, Guidi Nissim, Caparrotta, Masi, Azzarello, Pandolfi, Vignolini, Gonnelli and Mancuso [
3] and Mahjoor et al. [
37], who all recorded similar results with no statistical difference between CT and low seawater and freshwater blend treatments. The excessive concentration of sodium chloride in seawater may be the reason for stress on vegetables. During the study, it was seen that the plants, i.e., eggplant and pepper, managed its growth in the presence of higher seawater concentrations by reducing the freshwater consumption compared to control, while the tomato plants were severely disturbed by higher seawater treatment (C (20% SFW) treatment). The stressed plants were unable to absorb and translocate the water properly. Despite this, WUE and WP were increased in all three vegetables (
Figure 2). A review of the literature [
38,
39,
40] presented that at higher salt concentrations, WUE is decreased. In contrast to earlier studies, our results showed the increase of both WUE and WP in stress conditions with a significant decrease of water consumption for all three vegetables. It is observed in the literature that sensitive crops decreased water uptake to avoid further salt stress, while the tolerant crops used the water at the constant rate, including at higher levels of salinity [
2,
3]. That was also noted in the case of our study results.
The photosynthetic rate (Pn) was not negatively affected in the long term by higher concentrations of seawater in the cases of eggplant and pepper, showing a somewhat photosynthetic machinery adaptation at 15% and 20% seawater treatments as compared to tomato plants. Although all the vegetables showed poor growth under a saline environment as compared to CT, this factor could not restrict the plant leaf gas exchange and stomatal activities. These results are in line with those of Wilson et al. [
41], who noted that restricted plant growth did not hinder plant photosynthesis activities under different salinity conditions. Similarly, at higher salinity levels (C (20% SFW) treatment), the leaf pigment concentrations were not affected as compared to CT and at lower seawater concentrations (A; 5% SFW) and B; 10% SFW treatments). Our results are harmonious with those noted by Santos [
42]. The results of the current study suggest that at the three examined seawater blends, negligible reduction in the leaf pigments concentration was noted, which are not strong enough to decrease or stop the photosynthesis apparatus. Our findings are realized and concur to imply that the tested seawater concentrations can be used as growth media to grow these three tested crops.
The data showed that at an increasing order of seawater concentrations reduced the fruit quantity and quality of tomato, eggplant and pepper. The fruit yield of CT was comparable to a treatment (5% SFW) in the cases of eggplant and pepper, but fruit yield was decreased at 5% SFW treatments in the case of tomato, which showed sensitivity toward the increasing salinity environment. The results are consistent with those of Rameshwaran, Tepe, Yazar and Ragab [
19] and Shahbaz, et al. [
43]. This may be attributable to less water consumption under saline conditions and consequently less water and nutrient movement toward the fruit portion. The low concentration of SFW did not decrease the growth and biomass of the plants. In addition, the mineral contents present in the seawater may trigger the growth of plants [
1]. Our study results showed a strong relationship between plant water consumption and plant yield. This indicates that eggplant and pepper can easily adjust their metabolism between growth and salt stress to such an extent where their growth may not hamper [
3].
Titratable acidity (TA) was not influenced significantly in response to increasing seawater concentrations in eggplant but saw a significant change in tomato and pepper plants. These results are supported by Fernández-Garcí, et al. [
44], who recorded that treatments with the saline environment did not change TA in the fruits. This may be due to better acclimatization of eggplant in saline conditions with the passage of time. The increase in TA concentration resulted in considerable changes in fruit flavor [
45]. The reaction of all three vegetables toward total sugar, vitamin-C and NO
3-N was observed to be significant to increasing seawater, and their concentration was decreased at higher seawater concentrations (C (20% SFW) treatment). Generally, sugar contents are increased with the increase of salinity, but in our study, it decreased at higher levels of salinity (
Table 5). The possible reason for low sugar content may be due to less formation of hydrocarbon-synthesizing enzymes, which is responsible for the formation of sugar contents in plants. Similar results were noted by Navarro, et al. [
46] and stated that it could be due to an increase of ionic strength in the growing media, resulting a in the cost of higher fruit respiration [
47]. Vitamin C, or ascorbic acid, has the ability to lower salt stress in plants [
48,
49].
The depressing effect of increased seawater treatments was seen in the form of lower NO
3- concentration in the tissues of all three vegetables, but a more promising effect was recorded at 20% SFW [
50]. The statistically nondifference was noted between CT and A (5% SFW) treatments. The lower nitrate level in the plant tissues indicated that nutrient-induced salinity may create a hurdle toward intake, absorption and translocation in the plant body and it may be related to less water uptake by plants [
51]. Total oxalate is a major component of plant fruit palatability, and its higher concentrations may produce a bitter taste. Data presented in
Table 5,
Table 6 and
Table 7 indicate that at higher seawater concentrations (20% SFW), the oxalate level was increased in the fruits of all three vegetables [
52,
53]. Furthermore, no significant difference was seen among CT, A (5% SFW) and B (10% SFW) in the eggplant and pepper fruits, but significant variation was found between CT/A and B treatments in the fruits of the tomato plants. These results are consistent with those of Turhan, et al. [
14], who observed that total oxalate increased in the plants at higher seawater levels but did not significantly change at lower levels of seawater (2.5% and 5% seawater and freshwater blend). However, Carvalho, et al. [
54] noted the opposite results in purslane and stated that a sharp decrease in total oxalate concentration was observed over an increase in a saline environment in the purslane plants. Further, the transcriptional regulatory mechanism and modern biotechnological behind the overall improvement of physiological and nutritional attributes is also important in sustainability perspectives [
55,
56,
57].
Phenolic contents (PC) in all three vegetables indicated the positive correlation between salinity stress levels and sensitivity of plants. The superior level of PC in the case of our experiment may be due to osmotic and ionic stress on the plant metabolic system [
4]. In certain plants, such a stress increases or decreases the cell supporting the secondary metabolites, i.e., polyphenols [
5]. This results in disintegration of cell components or cytosol or serious damage to the cell membrane [
6]. Thus, the low and middle level of SFW did not considerably change in the concentrations of PC, MDA and ASA in eggplant and pepper plants as compared to tomato plants, therefore showing their possible cultivation at the tested low SFW concentrations. Additionally, the appropriate water and soil amendments such as leaching, organic matters, gypsum and lime may reduce the accumulation of salts and hereafter reduce the damaging impact of seawater on the crops.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}