
Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Plots and Experimental Treatments
2.2. Collecting Samples in the Field
2.3. Preparation of Soil and Root Samples
2.4. Preparing Root Samples for Illumina Sequencing
2.5. Element Analyses in Soil and Roots
2.6. Statistical Analyses
3. Results
3.1. Soil Properties and Root Chemistry Are Not Affected by Reduced Management Intensity
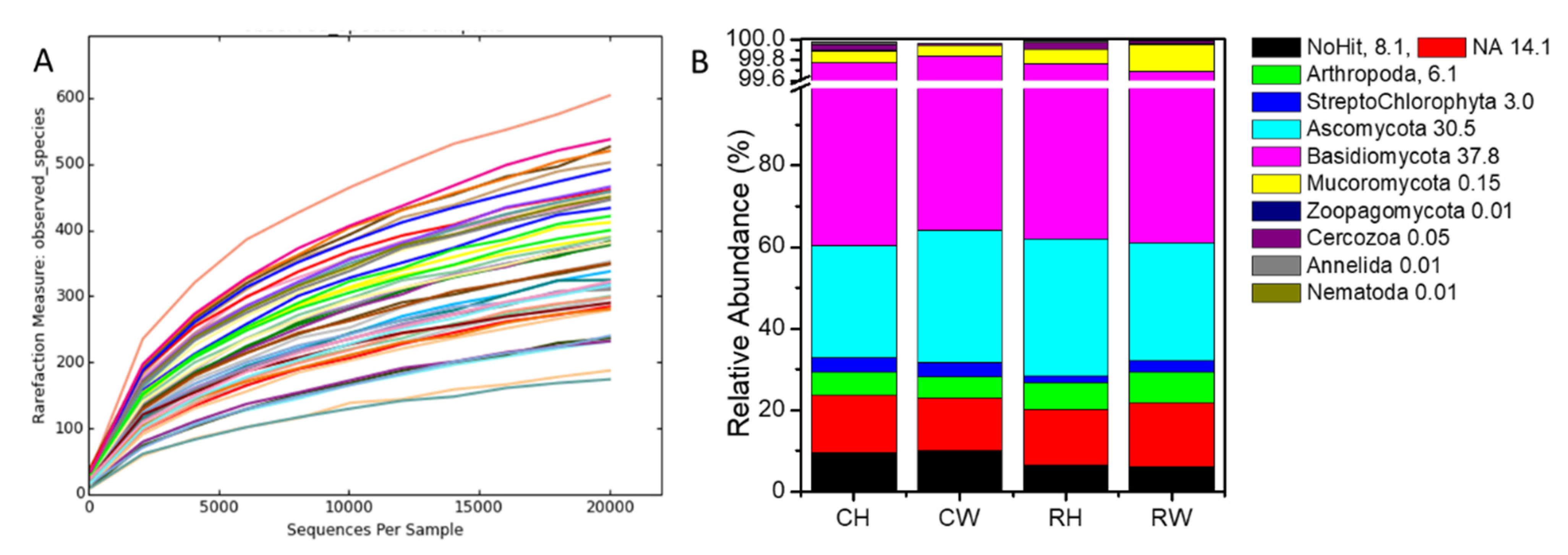
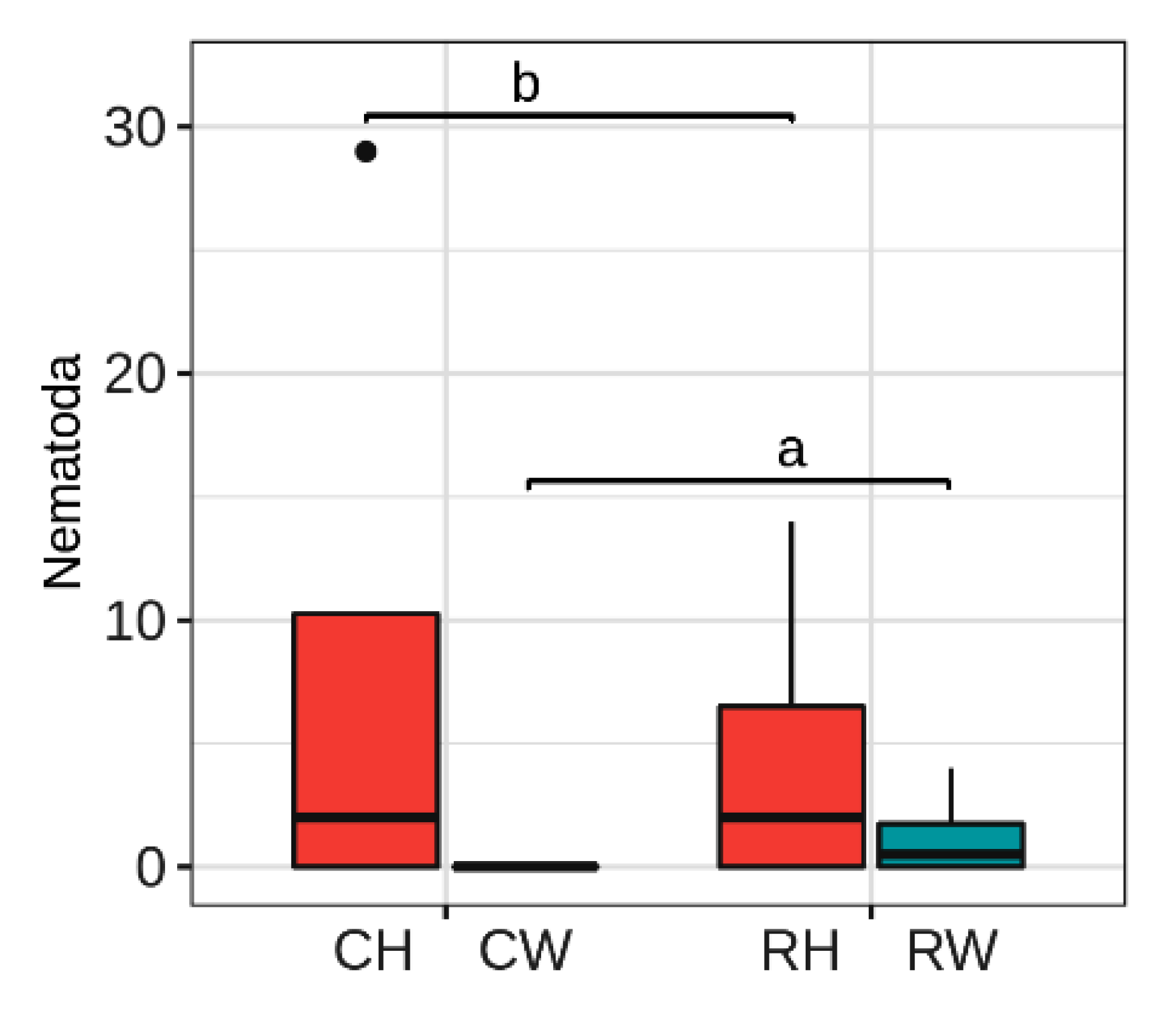
3.2. Diversity of Root-Associated Biota Is Not Affected by Reduced Management Intensity, but Nematodes Respond to Weeding
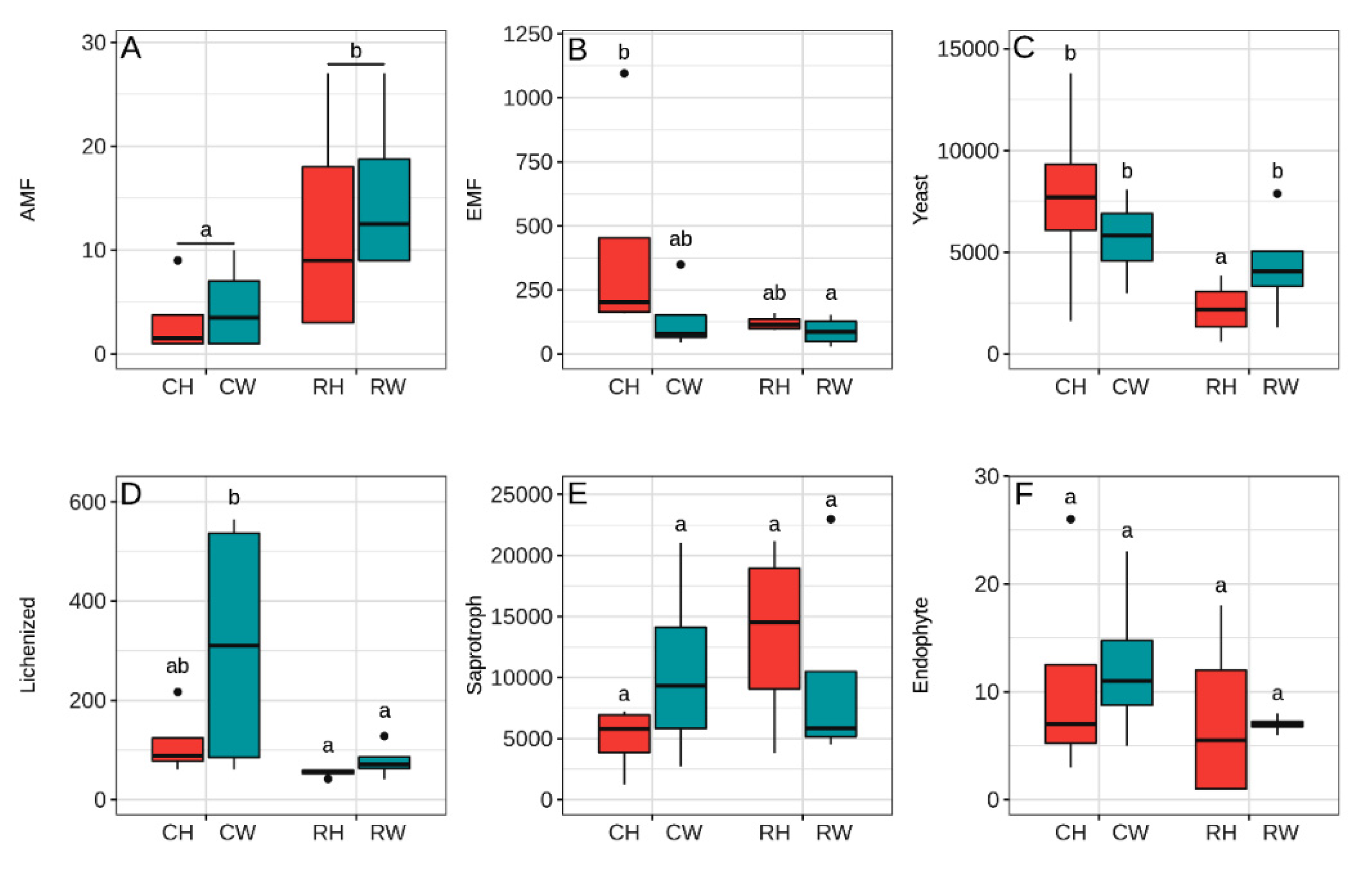
3.3. Fungal Guilds Are Shifted by Reduced Fertilizer and Weeding
4. Discussion
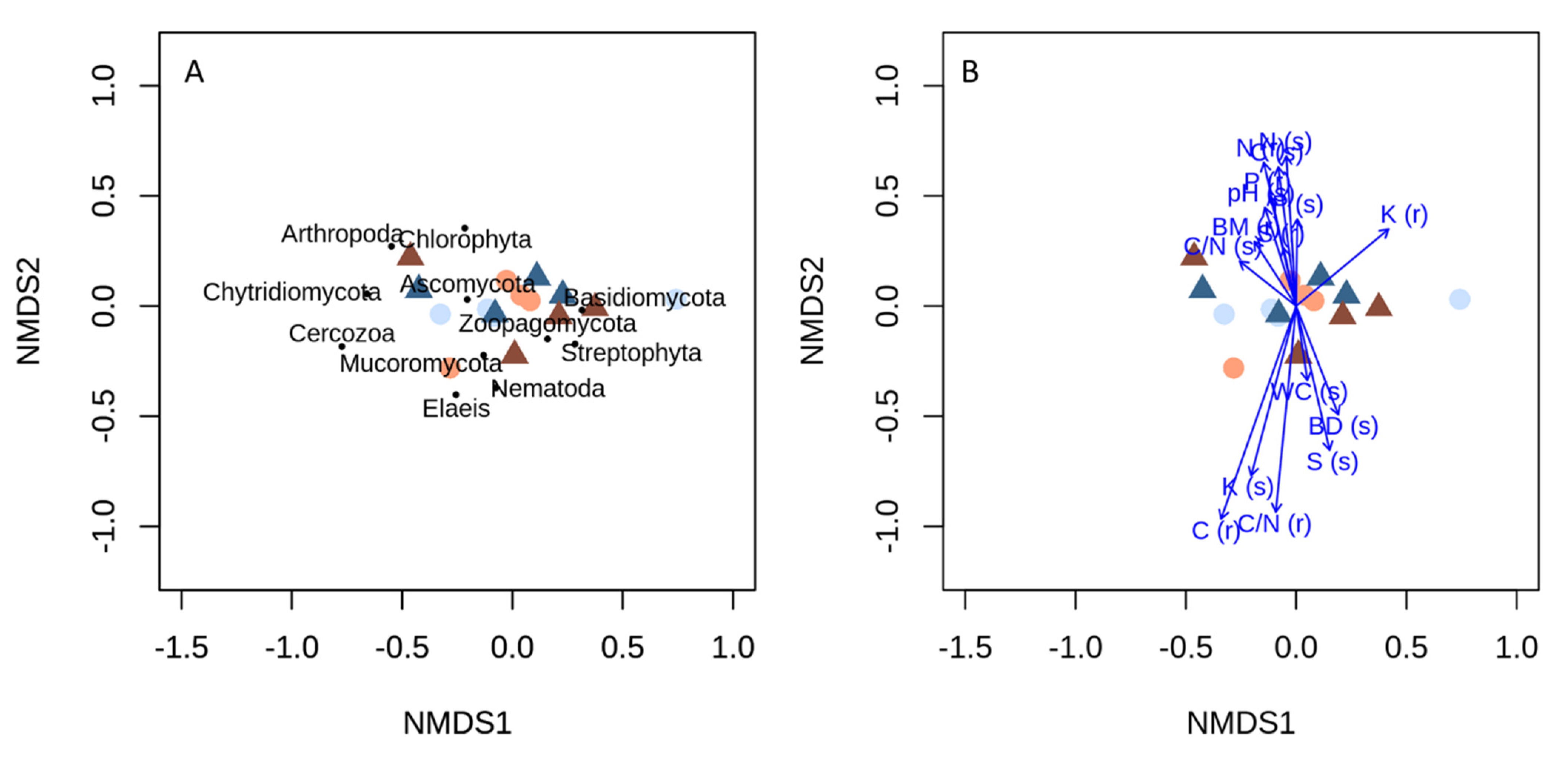
4.1. Reduced Management Intensity Does Not Affect Root and Soil Chemistry or Diversity of Root-Associated Phyla
4.2. Fungal Guilds Show Differentiated Patterns to Reduced Fertilizer Application and Weeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibbs, H.K.; Ruesch, A.S.; Achard, F.; Clayton, M.K.; Holmgren, P.; Ramankutty, N.; Foley, J.A. Tropical Forests Were the Primary Sources of New Agricultural Land in the 1980s and 1990s. Proc. Natl. Acad. Sci. USA 2010, 107, 16732–16737. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Rudel, T.K.; Aide, T.M.; Defries, R.; Emerson, R. A Contemporary Assessment of Change in Humid Tropical Forests. Conserv. Biol. J. Soc. Conserv. Biol. 2009, 23, 1386–1395. [Google Scholar] [CrossRef]
- Miller, D.C.; Ordoñez, P.J.; Brown, S.E.; Forrest, S.; Nava, N.J.; Hughes, K.; Baylis, K. The Impacts of Agroforestry on Agricultural Productivity, Ecosystem Services, and Human Well-Being in Low-and Middle-Income Countries: An Evidence and Gap Map. Campbell Syst. Rev. 2020, 16, e1066. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, C.; Paltseva, J.; Searle, S. Ecological Impacts of Palm Oil Expansion in Indonesia. 2016. Available online: https://theicct.org/publications/ecological-impacts-palm-oil-expansion-indonesia (accessed on 11 January 2022).
- Khatiwada, D.; Palmén, C.; Silveira, S. Evaluating the Palm Oil Demand in Indonesia: Production Trends, Yields, and Emerging Issues. Biofuels 2021, 12, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Euler, M.; Schwarze, S.; Siregar, H.; Qaim, M. Oil Palm Expansion among Smallholder Farmers in Sumatra, Indonesia. J. Agric. Econ. 2016, 67, 658–676. [Google Scholar] [CrossRef]
- Wijaya, A.; Sugardiman Budiharto, R.A.; Tosiani, A.; Murdiyarso, D.; Verchot, L.V. Assessment of Large Scale Land Cover Change Classifications and Drivers of Deforestation in Indonesia. ISPRS Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, XL-7/W3, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Kubitza, C.; Krishna, V.V.; Urban, K.; Alamsyah, Z.; Qaim, M. Land Property Rights, Agricultural Intensification, and Deforestation in Indonesia. Ecol. Econ. 2018, 147, 312–321. [Google Scholar] [CrossRef]
- Grass, I.; Kubitza, C.; Krishna, V.V.; Corre, M.D.; Mußhoff, O.; Pütz, P.; Drescher, J.; Rembold, K.; Ariyanti, E.S.; Barnes, A.D.; et al. Trade-offs between Multifunctionality and Profit in Tropical Smallholder Landscapes. Nat. Commun. 2020, 11, 1186. [Google Scholar] [CrossRef] [Green Version]
- Foster, W.A.; Snaddon, J.L.; Turner, E.C.; Fayle, T.M.; Cockerill, T.D.; Ellwood, M.D.F.; Broad, G.R.; Chung, A.Y.C.; Eggleton, P.; Khen, C.V.; et al. Establishing the Evidence Base for Maintaining Biodiversity and Ecosystem Function in the Oil Palm Landscapes of South East Asia. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3277–3291. [Google Scholar] [CrossRef] [Green Version]
- Drescher, J.; Rembold, K.; Allen, K.; Beckschäfer, P.; Buchori, D.; Clough, Y.; Faust, H.; Fauzi, A.M.; Gunawan, D.; Hertel, D. Ecological and Socio-Economic Functions across Tropical Land Use Systems after Rainforest Conversion. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150275. [Google Scholar] [CrossRef] [PubMed]
- Fitzherbert, E.B.; Struebig, M.J.; Morel, A.; Danielsen, F.; Brühl, C.A.; Donald, P.F.; Phalan, B. How Will Oil Palm Expansion Affect Biodiversity? Trends Ecol. Evol. 2008, 23, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Rembold, K.; Mangopo, H.; Tjitrosoedirdjo, S.S.; Kreft, H. Plant Diversity, Forest Dependency, and Alien Plant Invasions in Tropical Agricultural Landscapes. Biol. Conserv. 2017, 213, 234–242. [Google Scholar] [CrossRef]
- Allen, K.; Corre, M.D.; Tjoa, A.; Veldkamp, E. Soil Nitrogen-Cycling Responses to Conversion of Lowland Forests to Oil Palm and Rubber Plantations in Sumatra, Indonesia. PLoS ONE 2015, 10, e0133325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röll, A.; Niu, F.; Meijide, A.; Ahongshangbam, J.; Ehbrecht, M.; Guillaume, T.; Gunawan, D.; Hardanto, A.; Hertel, D.; Kotowska, M.M.; et al. Transpiration on the Rebound in Lowland Sumatra. Agric. For. Meteorol. 2019, 274, 160–171. [Google Scholar] [CrossRef]
- Guillaume, T.; Kotowska, M.M.; Hertel, D.; Knohl, A.; Krashevska, V.; Murtilaksono, K.; Scheu, S.; Kuzyakov, Y. Carbon Costs and Benefits of Indonesian Rainforest Conversion to Plantations. Nat. Commun. 2018, 9, 2388. [Google Scholar] [CrossRef] [PubMed]
- Sahner, J.; Budi, S.W.; Barus, H.; Edy, N.; Meyer, M.; Corre, M.D.; Polle, A. Degradation of Root Community Traits as Indicator for Transformation of Tropical Lowland Rain Forests into Oil Palm and Rubber Plantations. PLoS ONE 2015, 10, e0138077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senior, M.J.M.; Hamer, K.C.; Bottrell, S.; Edwards, D.P.; Fayle, T.M.; Lucey, J.M.; Mayhew, P.J.; Newton, R.; Peh, K.S.-H.; Sheldon, F.H.; et al. Trait-Dependent Declines of Species Following Conversion of Rain Forest to Oil Palm Plantations. Biodivers. Conserv. 2013, 22, 253–268. [Google Scholar] [CrossRef]
- Clough, Y.; Krishna, V.V.; Corre, M.D.; Darras, K.; Denmead, L.H.; Meijide, A.; Moser, S.; Musshoff, O.; Steinebach, S.; Veldkamp, E.; et al. Land-Use Choices Follow Profitability at the Expense of Ecological Functions in Indonesian Smallholder Landscapes. Nat. Commun. 2016, 7, 13137. [Google Scholar] [CrossRef]
- Krashevska, V.; Klarner, B.; Widyastuti, R.; Maraun, M.; Scheu, S. Impact of Tropical Lowland Rainforest Conversion into Rubber and Oil Palm Plantations on Soil Microbial Communities. Biol. Fertil. Soils 2015, 51, 697–705. [Google Scholar] [CrossRef]
- Schneider, D.; Engelhaupt, M.; Allen, K.; Kurniawan, S.; Krashevska, V.; Heinemann, M.; Nacke, H.; Wijayanti, M.; Meryandini, A.; Corre, M.D.; et al. Impact of Lowland Rainforest Transformation on Diversity and Composition of Soil Prokaryotic Communities in Sumatra (Indonesia). Front. Microbiol. 2015, 6, 1339. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, N.; Schneider, D.; Sahner, J.; Ballauff, J.; Edy, N.; Barus, H.; Irawan, B.; Budi, S.W.; Qaim, M.; Daniel, R.; et al. Intensive Tropical Land Use Massively Shifts Soil Fungal Communities. Sci. Rep. 2019, 9, 3403. [Google Scholar] [CrossRef] [Green Version]
- Kerfahi, D.; Tripathi, B.M.; Lee, J.; Edwards, D.P.; Adams, J.M. The Impact of Selective-Logging and Forest Clearance for Oil Palm on Fungal Communities in Borneo. PLoS ONE 2014, 9, e111525. [Google Scholar] [CrossRef] [PubMed]
- Schulz, G.; Schneider, D.; Brinkmann, N.; Edy, N.; Daniel, R.; Polle, A.; Scheu, S.; Krashevska, V. Changes in Trophic Groups of Protists with Conversion of Rainforest into Rubber and Oil Palm Plantations. Front. Microbiol. 2019, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkelmann, D.; Schneider, D.; Engelhaupt, M.; Heinemann, M.; Christel, S.; Wijayanti, M.; Meryandini, A.; Daniel, R. How Rainforest Conversion to Agricultural Systems in Sumatra (Indonesia) Affects Active Soil Bacterial Communities. Front. Microbiol. 2018, 9, 2381. [Google Scholar] [CrossRef] [PubMed]
- McGuire, K.L.; D’Angelo, H.; Brearley, F.Q.; Gedallovich, S.M.; Babar, N.; Yang, N.; Gillikin, C.M.; Gradoville, R.; Bateman, C.; Turner, B.L.; et al. Responses of Soil Fungi to Logging and Oil Palm Agriculture in Southeast Asian Tropical Forests. Microb. Ecol. 2015, 69, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Corley, R.H.V.; Tinker, P.B.H. The Oil Palm; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 978-0-470-75036-0. [Google Scholar]
- Goh, K.J. Fertilizer Recommendation Systems for Oil Palm: Estimating the Fertilizer Rates. 2004. Available online: http://www.oneoilpalm.com/wp-content/uploads/2015/09/Fertilizer-recommendation-systems-for-oil-palm-estimating-the-fertilizer-rates.pdf (accessed on 7 January 2022).
- Darras, K.F.A.; Corre, M.D.; Formaglio, G.; Tjoa, A.; Potapov, A.; Brambach, F.; Sibhatu, K.T.; Grass, I.; Rubiano, A.A.; Buchori, D.; et al. Reducing Fertilizer and Avoiding Herbicides in Oil Palm Plantations—Ecological and Economic Valuations. Front. For. Glob. Chang. 2019, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Luke, S.H.; Advento, A.D.; Aryawan, A.A.K.; Adhy, D.N.; Ashton-Butt, A.; Barclay, H.; Dewi, J.P.; Drewer, J.; Dumbrell, A.J.; Eycott, A.E.; et al. Managing Oil Palm Plantations More Sustainably: Large-Scale Experiments within the Biodiversity and Ecosystem Function in Tropical Agriculture (BEFTA) Programme. Front. For. Glob. Chang. 2020, 2, 75. [Google Scholar] [CrossRef] [Green Version]
- Bruggen, A.V.V.; He, M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.; Morris, J.G. Environmental and Health Effects of the Herbicide Glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, S.W.; Ewel, J.J.; Haggar, J.P. Enhancing Nutrient Retention in Tropical Tree Plantations: No Short Cuts. Ecol. Appl. 2004, 14, 28–46. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, Biogeochemistry and Ecological Relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Pierret, A.; Doussan, C.; Capowiez, Y.; Bastardie, F.; Pagès, L. Root Functional Architecture: A Framework for Modeling the Interplay between Roots and Soil. Vadose Zone J. 2007, 6, 269–281. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The Rhizosphere: A Playground and Battlefield for Soilborne Pathogens and Beneficial Microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Elshafie, H.S.; Camele, I. Structural and Functional Organization of the Root System: A Comparative Study on Five Plant Species. Plants Basel Switz. 2020, 9, E1338. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Simon, L.; Lalonde, M.; Bruns, T. Specific Amplification of 18S Fungal Ribosomal Genes from Vesicular-Arbuscular Endomycorrhizal Fungi Colonizing Roots. Appl. Environ. Microbiol. 1992, 58, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR Primers for the Detection and Identification of Arbuscular Mycorrhizal Fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR a Fast and Accurate Illumina Paired-End ReAd MergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, R.H.; Tedersoo, L.; Ryberg, M.; Kristiansson, E.; Hartmann, M.; Unterseher, M.; Porter, T.M.; Bengtsson-Palme, J.; Walker, D.M.; de Sousa, F.; et al. A Comprehensive, Automatically Updated Fungal ITS Sequence Dataset for Reference-Based Chimera Control in Environmental Sequencing Efforts. Microbes Environ. 2015, 30, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE Database for Molecular Identification of Fungi—Recent Updates and Future Perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2015. Available online: http://CRAN.R-project.org/package=vegan (accessed on 11 January 2022).
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of Total, Organic, and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Formaglio, G.; Veldkamp, E.; Damris, M.; Tjoa, A.; Corre, M.D. Mulching with Pruned Fronds Promotes the Internal Soil N Cycling and Soil Fertility in a Large-Scale Oil Palm Plantation. Biogeochemistry 2021, 154, 63–80. [Google Scholar] [CrossRef]
- Ashton-Butt, A.; Aryawan, A.A.; Hood, A.S.; Naim, M.; Purnomo, D.; Wahyuningsih, R.; Willcock, S.; Poppy, G.M.; Caliman, J.P.; Turner, E.C.; et al. Understory Vegetation in Oil Palm Plantations Benefits Soil Biodiversity and Decomposition Rates. Front. For. Glob. Chang. 2018, 1, 10. [Google Scholar] [CrossRef]
- Silver, W.L.; Thompson, A.W.; Reich, A.; Ewel, J.J.; Firestone, M.K. Nitrogen Cycling in Tropical Plantation Forests: Potential Controls on Nitrogen Retention. Ecol. Appl. 2005, 15, 1604–1614. [Google Scholar] [CrossRef]
- Ballauff, J.; Zemp, D.C.; Schneider, D.; Irawan, B.; Daniel, R.; Polle, A. Legacy Effects Overshadow Tree Diversity Effects on Soil Fungal Communities in Oil Palm-Enrichment Plantations. Microorganisms 2020, 8, 1577. [Google Scholar] [CrossRef]
- Turley, N.E.; Bell-Dereske, L.; Evans, S.E.; Brudvig, L.A. Agricultural Land-Use History and Restoration Impact Soil Microbial Biodiversity. J. Appl. Ecol. 2020, 57, 852–863. [Google Scholar] [CrossRef]
- Corre, M.D.; Veldkamp, E.; Arnold, J.; Wright, S.J. Impact of Elevated N Input on Soil N Cycling and Losses in Old-Growth Lowland and Montane Forests in Panama. Ecology 2010, 91, 1715–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, S.J.; Matson, P.A. Nitrogen Oxide Emissions after Nitrogen Additions in Tropical Forests. Nature 1999, 400, 152–155. [Google Scholar] [CrossRef]
- Berkelmann, D.; Schneider, D.; Hennings, N.; Meryandini, A.; Daniel, R. Soil Bacterial Community Structures in Relation to Different Oil Palm Management Practices. Sci. Data 2020, 7, 421. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode Community Structure as a Bioindicator in Environmental Monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Krashevska, V.; Kudrin, A.A.; Widyastuti, R.; Scheu, S. Changes in Nematode Communities and Functional Diversity with the Conversion of Rainforest into Rubber and Oil Palm Plantations. Front. Ecol. Evol. 2019, 7, 487. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Ongena, M.; Wang, Q.; Guan, D.; Cao, F.; Jiang, X.; Li, J. Chronic Fertilization of 37 Years Alters the Phylogenetic Structure of Soil Arbuscular Mycorrhizal Fungi in Chinese Mollisols. AMB Express 2018, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; Manoharan, L.; Rosenstock, N.P.; Olsson, P.A.; Hedlund, K. Long-Term Agricultural Fertilization Alters Arbuscular Mycorrhizal Fungal Community Composition and Barley (Hordeum Vulgare) Mycorrhizal Carbon and Phosphorus Exchange. New Phytol. 2017, 213, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Feng, J.; Han, M.; Zhu, B. Responses of Arbuscular Mycorrhizal Fungi to Nitrogen Addition: A Meta-Analysis. Glob. Change Biol. 2020, 26, 7229–7241. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling Mycorrhiza-Induced Resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.-A.; Bessis, A.; Pozo, M.J. Microbial Priming of Plant and Animal Immunity: Symbionts as Developmental Signals. Trends Microbiol. 2014, 22, 607–613. [Google Scholar] [CrossRef]
- Starmer, W.T.; Lachance, M.-A. Chapter 6—Yeast Ecology. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 65–83. ISBN 978-0-444-52149-1. [Google Scholar]
- Ballauff, J.; Schneider, D.; Edy, N.; Irawan, B.; Daniel, R.; Polle, A. Shifts in Root and Soil Chemistry Drive the Assembly of Belowground Fungal Communities in Tropical Land-Use Systems. Soil Biol. Biochem. 2021, 154, 108140. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Comandini, O.; Kuyper, T.W. Ectomycorrhizal Fungal Diversity: Seperating the Wheat from the Chaff. Fungal Divers. 2008, 33, 1–45. [Google Scholar]
- Mohan Kumar, B. Litter Dynamics in Plantation and Agroforestry Systems of the Tropics—A Review of Observations and Methods. In Ecological Basis of Agroforestry; Rani Batish, D., Kumar Kohli, R., Jose, S., Pal Singh, H., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 181–216. ISBN 978-1-4200-4327-3. [Google Scholar]
- Tuovila, H.; Davey, M.; Yan, L.; Huhtinen, S.; Rikkinen, J. New Resinicolous Chaenothecopsis Species from China. Mycologia 2014, 106, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Rikkinen, J.; Poinar, G. A New Species of Resinicolous Chaenothecopsis (Mycocaliciaceae, Ascomycota) from 20 Million Year Old Bitterfeld Amber, with Remarks on the Biology of Resinicolous Fungi. Mycol. Res. 2000, 104, 7–15. [Google Scholar] [CrossRef]
- Beimforde, C.; Tuovila, H.; Schmidt, A.R.; Lee, W.G.; Gube, M.; Rikkinen, J. Chaenothecopsis Schefflerae (Ascomycota: Mycocaliciales): A Widespread Fungus on Semi-Hardened Exudates of Endemic New Zealand Araliaceae. N. Z. J. Bot. 2017, 55, 387–406. [Google Scholar] [CrossRef] [Green Version]
- Haney, R.L.; Senseman, S.A.; Hons, F.M.; Zuberer, D.A. Effect of Glyphosate on Soil Microbial Activity and Biomass. Weed Sci. 2000, 48, 89–93. [Google Scholar] [CrossRef]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Glyphosate Effects on Soil Rhizosphere-Associated Bacterial Communities. Sci. Total Environ. 2016, 543, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Arfarita, N.; Imai, T.; Prasetya, B. Potential Use of Soil-Born Fungi Isolated from Treated Soil in Indonesia to Degrade Glyphosate Herbicide. J. Degrad. Min. Lands Manag. 2014, 1, 63–68. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
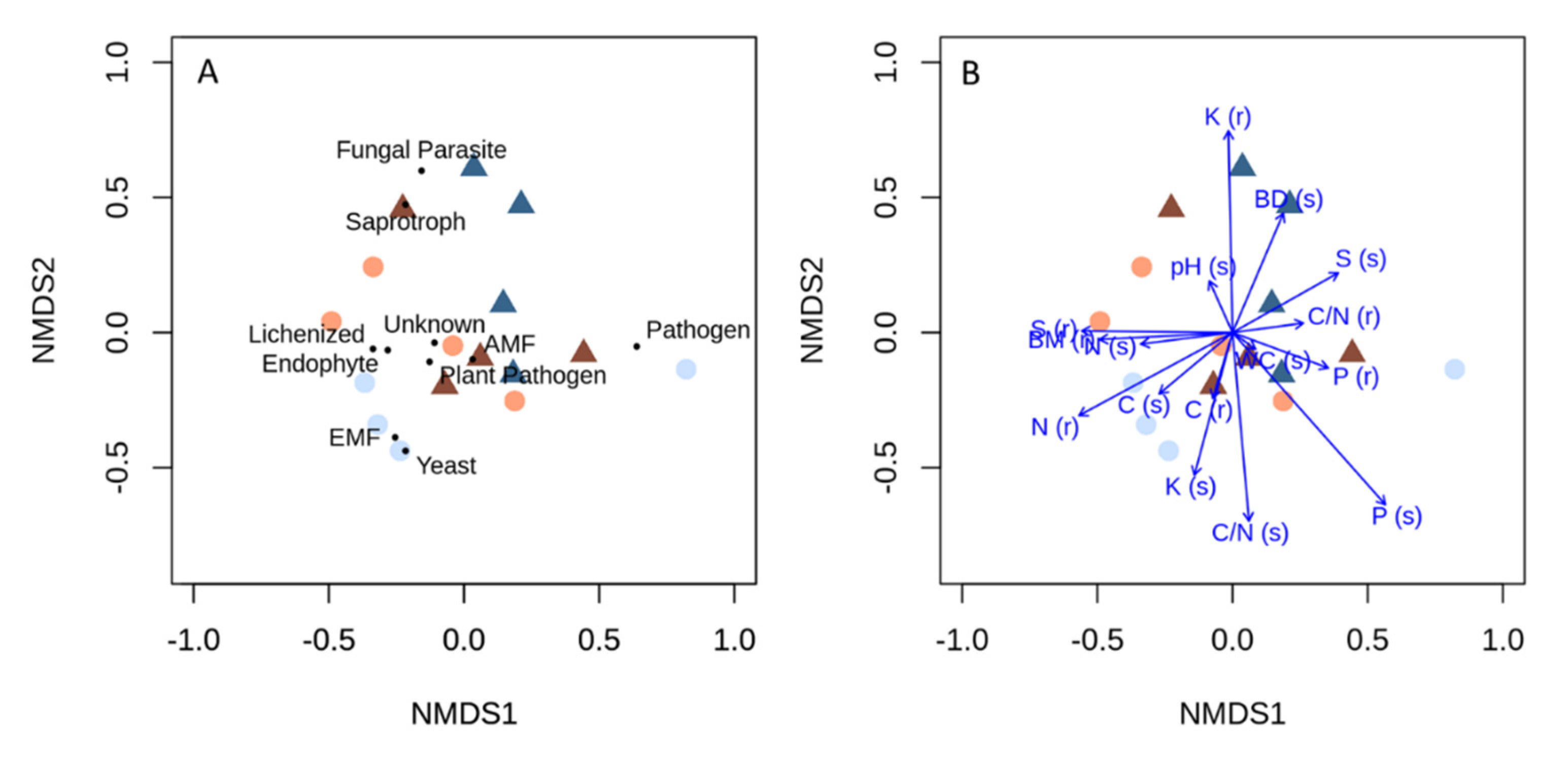{kind=link}
| Parameter | CH | CW | RH | RW | pf | Pw | Pfxw |
|---|---|---|---|---|---|---|---|
| C | 13.19 ± 1.15 | 12.55 ± 2.05 | 13.22 ± 1.60 | 14.15 ± 0.91 | 0.586 | 0.923 | 0.600 |
| N | 0.823 ± 0.069 | 0.808 ± 0.132 | 0.849 ± 0.095 | 0.858 ± 0.045 | 0.678 | 0.970 | 0.899 |
| P | 0.144 ± 0.033 | 0.164 ± 0.044 | 0.101 ± 0.019 | 0.186 ± 0.050 | 0.777 | 0.179 | 0.402 |
| S | 0.044 ± 0.002 | 0.054 ± 0.005 | 0.050 ± 0.005 | 0.052 ± 0.004 | 0.636 | 0.138 | 0.322 |
| K | 0.029 ± 0.005 | 0.032 ± 0.007 | 0.024 ± 0.002 | 0.025 ± 0.003 | 0.172 | 0.586 | 0.853 |
| Na | 0.042 ± 0.001 | 0.083 ± 0.023 | 0.058 ± 0.007 | 0.059 ± 0.008 | 0.754 | 0.094 | 0.124 |
| Mg | 0.139 ± 0.023 | 0.137 ± 0.019 | 0.144 ± 0.032 | 0.163 ± 0.033 | 0.573 | 0.761 | 0.685 |
| Ca | 0.395 ± 0.057 | 0.513 ± 0.095 | 0.410 ± 0.082 | 0.489 ± 0.081 | 0.961 | 0.224 | 0.808 |
| Mn | 0.005 ± 0.001 | 0.005 ± 0.001 | 0.005 ± 0.001 | 0.006 ± 0.001 | 0.854 | 0.364 | 0.535 |
| Fe | 1.007 ± 0.055 | 1.086 ± 0.037 | 1.168 ± 0.051 | 1.110 ± 0.058 | 0.076 | 0.837 | 0.185 |
| Al | 5.99 ± 0.36 | 6.57 ± 0.49 | 6.28 ± 0.58 | 6.64 ± 0.57 | 0.724 | 0.357 | 0.828 |
| C/N | 16.07 ± 0.383 | 14.29 ± 1.350 | 15.43 ± 0.405 | 16.45 ± 0.400 | 0.324 | 0.618 | 0.072 |
| Soil pH | 6.76 ± 0.05 | 6.84 ± 0.03 | 6.85 ± 0.02 | 6.85 ± 0.04 | 0.227 | 0.300 | 0.300 |
| MC | 20.06 ± 0.50 | 20.70 ± 0.76 | 20.21 ± 0.98 | 19.45 ± 0.71 | 0.471 | 0.943 | 0.362 |
| SBD | 1.31 ± 0.02 | 1.28 ± 0.03 | 1.33 ± 0.03 | 1.30 ± 0.03 | 0.506 | 0.415 | 0.969 |
| Parameter | CH | CW | RH | RW | Pf | Pw | Pfxw |
|---|---|---|---|---|---|---|---|
| C | 427.1 ± 19.7 | 440.1 ± 22.7 | 457.9 ± 30.2 | 479.3 ± 28.5 | 0.179 | 0.506 | 0.871 |
| N | 5.91 ± 0.26 | 6.57 ± 0.49 | 5.86 ± 0.24 | 5.73 ± 0.24 | 0.180 | 0.429 | 0.233 |
| P | 0.321 ± 0.016 | 0.332 ± 0.043 | 0.400 ± 0.095 | 0.434 ± 0.075 | 0.170 | 0.729 | 0.856 |
| S | 0.680 ± 0.047 | 0.731 ± 0.052 | 0.651 ± 0.027 | 0.690 ± 0.035 | 0.404 | 0.285 | 0.882 |
| K | 0.318 ± 0.055 | 0.356 ± 0.052 | 0.391 ± 0.073 | 0.412 ± 0.116 | 0.415 | 0.708 | 0.914 |
| Na | 0.340 ± 0.033 | 0.318 ± 0.043 | 0.221 ± 0.026 | 0.291 ± 0.046 | 0.059 | 0.534 | 0.231 |
| Mg | 0.723 ± 0.085 | 0.724 ± 0.083 | 0.847 ± 0.109 | 0.965 ± 0.145 | 0.100 | 0.586 | 0.592 |
| Ca | 1.749 ± 0.188 | 1.892 ± 0.328 | 4.362 ± 2.313 | 2.891 ± 0.930 | 0.159 | 0.601 | 0.525 |
| Mn | 0.039 ± 0.012 | 0.033 ± 0.005 | 0.047 ± 0.018 | 0.037 ± 0.016 | 0.676 | 0.591 | 0.894 |
| Fe | 6.124 ± 2.041 | 5.127 ± 0.803 | 7.149 ± 2.983 | 3.868 ± 1.301 | 0.953 | 0.282 | 0.564 |
| Al | 11.87 ± 0.73 | 13.65 ± 1.04 | 12.33 ± 0.97 | 10.49 ± 1.19 | 0.184 | 0.981 | 0.077 |
| C/N | 73.3 ± 4.5 | 72.0 ± 7.1 | 80.3 ± 7.2 | 86.1 ± 7.3 | 0.122 | 0.738 | 0.596 |
| RB | 0.211 ± 0.032 | 0.252 ± 0.072 | 0.169 ± 0.040 | 0.165 ± 0.025 | 0.167 | 0.685 | 0.628 |
| Index | CH | CW | RH | RW | Pf | Pw | Pfxw |
|---|---|---|---|---|---|---|---|
| Shannon | 3.45 ± 0.46 | 3.65 ± 0.18 | 3.29 ± 0.25 | 3.49 ± 0.23 | 0.606 | 0.510 | 0.996 |
| Richness | 790 ± 87 | 856 ± 27 | 737 ± 48 | 768 ± 54 | 0.249 | 0.423 | 0.772 |
| Evenness | 0.047 ± 0.012 | 0.047 ± 0.008 | 0.039 ± 0.007 | 0.045 ± 0.007 | 0.564 | 0.733 | 0.741 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryadin, A.R.; Janz, D.; Schneider, D.; Tjoa, A.; Irawan, B.; Daniel, R.; Polle, A. Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations. Agronomy 2022, 12, 199. https://doi.org/10.3390/agronomy12010199
Ryadin AR, Janz D, Schneider D, Tjoa A, Irawan B, Daniel R, Polle A. Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations. Agronomy. 2022; 12(1):199. https://doi.org/10.3390/agronomy12010199
Chicago/Turabian StyleRyadin, Aisjah R., Dennis Janz, Dominik Schneider, Aiyen Tjoa, Bambang Irawan, Rolf Daniel, and Andrea Polle. 2022. "Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations" Agronomy 12, no. 1: 199. https://doi.org/10.3390/agronomy12010199
APA StyleRyadin, A. R., Janz, D., Schneider, D., Tjoa, A., Irawan, B., Daniel, R., & Polle, A. (2022). Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations. Agronomy, 12(1), 199. https://doi.org/10.3390/agronomy12010199