

Heterologous Overexpression of ZmHDZIV13 Enhanced Drought and Salt Tolerance in Arabidopsis and Tobacco


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Bacterial Strain, Plasmid and Transformation
2.3. Measurement of Germination Rate and Root Traits and Evaluation of Drought Resistance
2.4. Measurement of Leaf Stomatal Density, Photosynthesis Rate, Transpiration Rate and Water-Use Efficiency
2.5. Measurement of MDA and Proline Contents
2.6. Gene Expression Analysis
2.7. Statistical Analysis
3. Results
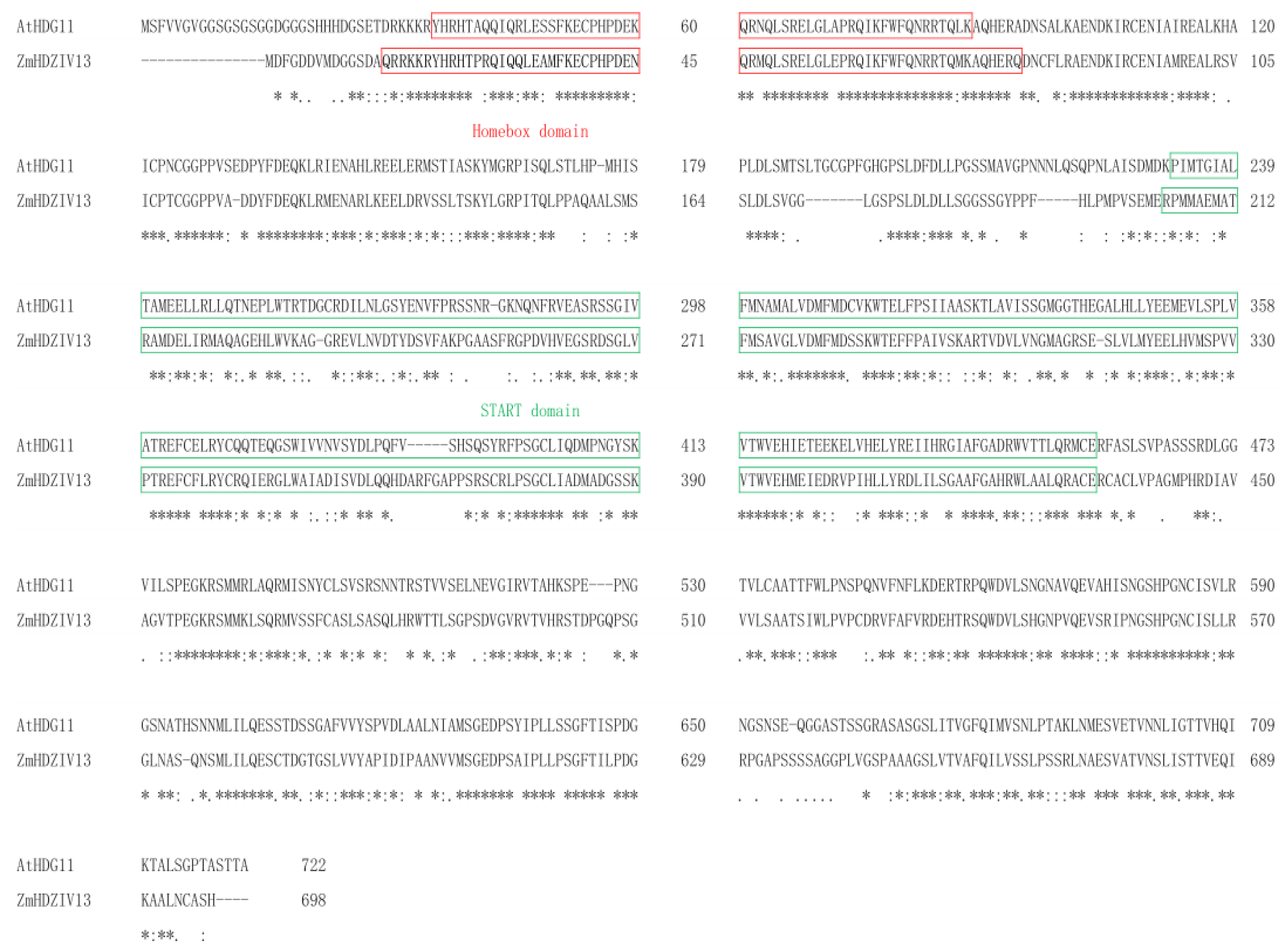
3.1. Informatics Analysis of ZmHDZIV13
3.2. Transformation of ZmHDZIV13 into Arabidopsis
3.3. Morphological and Physiological Characteristics in Response to Drought Stress
3.4. Morphological Characteristics in Response to Different ABA Levels
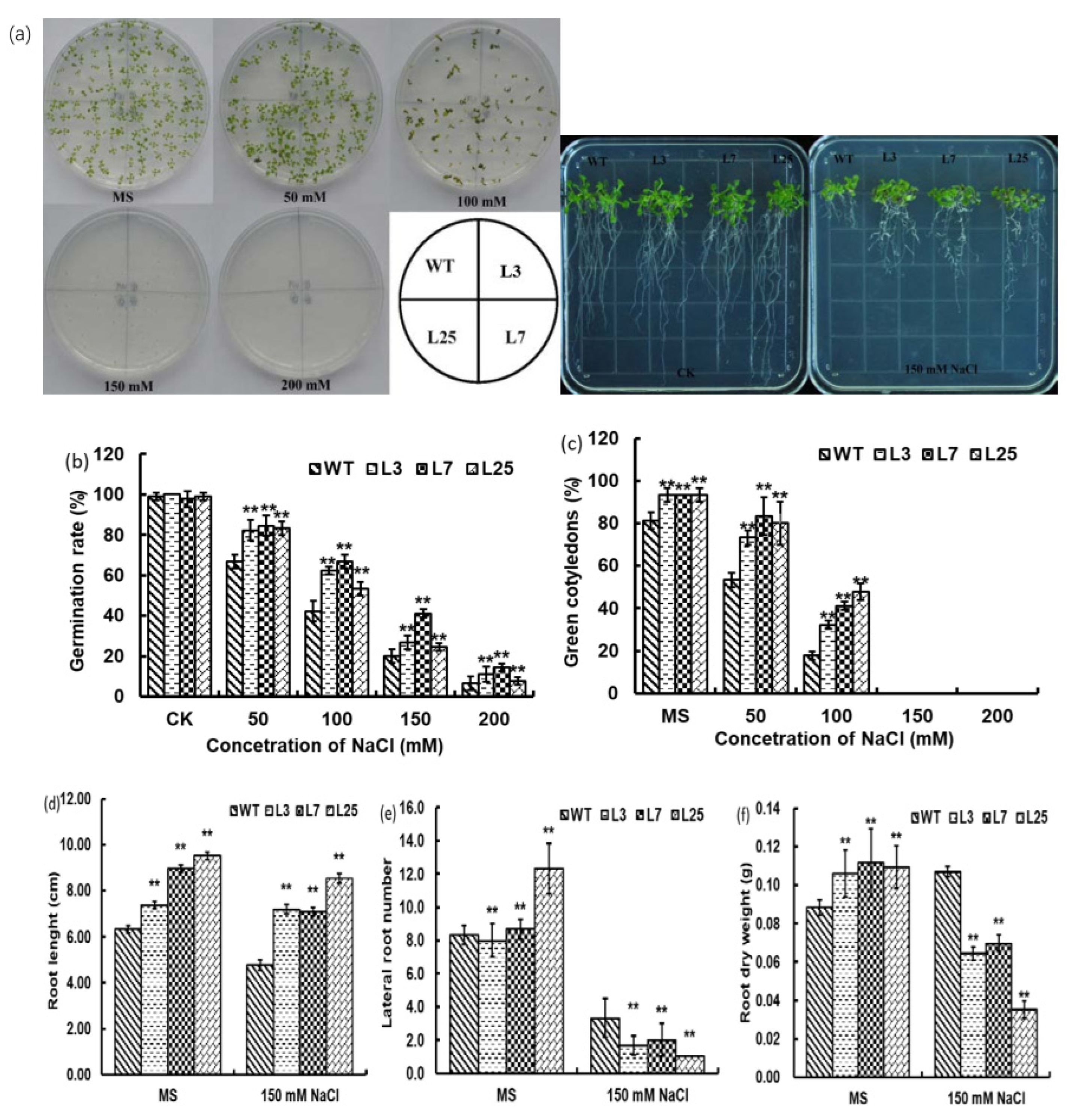
3.5. Morphological Characteristics in Response to NaCl Treatments and Osmotic Stress in Arabidopsis
3.6. Relative Expression Levels of Stress Response Genes under Drought Stress
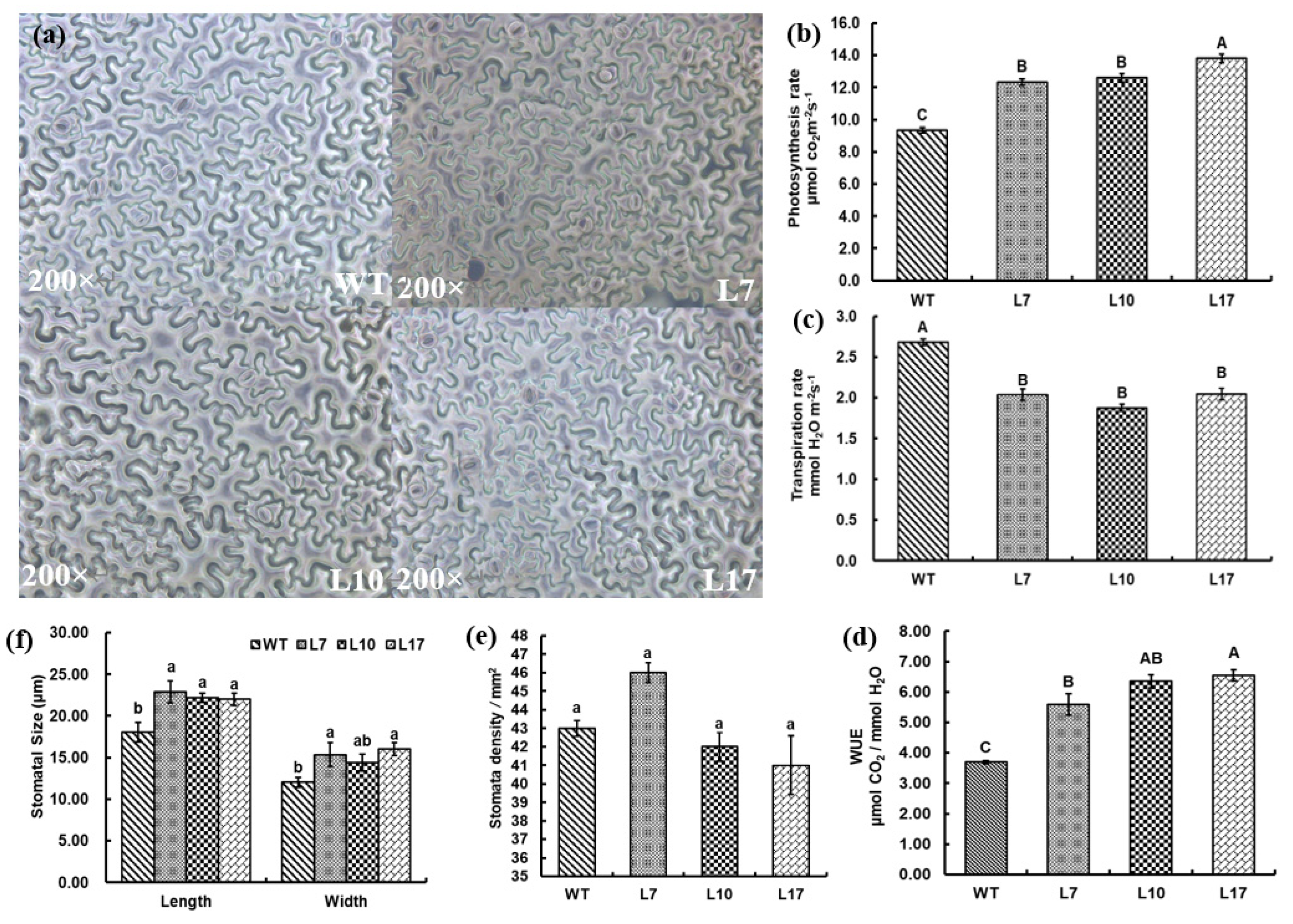
3.7. Leaf Density and Water-Use Efficiency in Transgenic Tobacco
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Verslues, P.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Zhu, J. Salt and drought stress signal transduction in plant. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerances by transgenic technology. Front. Plant Sci. 2016, 7, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Johannesson, H.; Wang, Y.; Hanson, J.; Engstrom, P. The Arabidopsis thaliana homeobox gene ATHB5 is a potential regulator of abscisic acid responsiveness in developing seedlings. Plant Mol. Biol. 2003, 51, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Manavella, P.; Arce, A.; Dezar, C.; Bitton, F.; Renou, J.; Crespi, M.; Chan, R. Cross-talk between ethylene and drought signalling pathways is mediated by the sunflower Hahb-4 transcription factor. Plant J. 2006, 48, 125–137. [Google Scholar] [CrossRef]
- Agalou, A.; Purwantomo, S.; Overnas, E.; Johannesson, H.; Zhu, X.; Estiati, A.; Kam, R.; Engström, P.; Slamet-Loedin, I.; Zhu, Z.; et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Hong, Y.; Yin, M.; Li, C.; Zhang, K.; Grierson, D. A tomato HD-Zip homeobox protein, Le HB-1, plays an important role in floral organogenesis and ripening. Plant J. 2008, 55, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernoud, V.; Laigle, G.; Rozier, F.; Meeley, R.B.; Perez, P.; Rogowsky, P.M. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize. Plant J. 2009, 59, 883–894. [Google Scholar] [CrossRef]
- Ariel, F.; Diet, A.; Verdenaud, M.; Gruber, V.; Frugier, F.; Chan, R.; Crespi, M. Environmental regulation of lateral root emergence in Medicago truncatula requires the HD-Zip I transcription factor HB1. Plant Cell. 2010, 22, 2171–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, H.; Shu, D.; Wang, M.; Xing, G.; Zhan, H.; Du, X.; Song, W.; Nie, X. Genome-wide identification, and expression analysis of the HD-Zip gene family in wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiong, H.; Cuo, D.; Wu, X.; Duan, R. Genome-wide characterization, and expression profiling of the relation of the HD-Zip gene family to abiotic stress in barley (Hordeum vulgare L.). Plant Physiol. Biochem. 2019, 141, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Dong, J.; Cao, M.; Gao, X.; Wang, D.; Liu, B.; Chen, Q. Genome-wide identification, and characterization of HD-ZIP genes in potato. Gene 2019, 697, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Rahat, S. Genome Wide Identification, Characterization and Expression Analysis of HD-ZIP Gene Family in Cucumis sativus L. under Biotic and Abiotic Stresses. Master’s Thesis, Northwest A & F University, Xianyang, China, 2020. [Google Scholar] [CrossRef]
- Han, L.; Guo, Q.; Zhao, K.; Jiang, T.; Zhou, B.; Li, L. Cloning and expression analysis of HD-Zip transcription factor PsnHB63 in Popul. Simonii × P. Nigra. Bull. Bot. Res. 2021, 41, 1006–1014. [Google Scholar] [CrossRef]
- Hu, J.; Li, Y.; Duan, Y.; Ruan, Y.; Gu, X.; Xiao, G. Identification and expression analysis of the HD-Zip I subfamily genes in mulberry. For. Res. 2022, 35, 130–142. [Google Scholar] [CrossRef]
- Ariel, F.; Manavella, P.; Dezar, C.; Chan, R. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Itoh, J.; Hibara, K.; Sato, Y.; Nagato, Y. Developmental role and auxin responsiveness of Class III homeodomain leucine zipper gene family members in rice. Plant Physiol. 2008, 147, 1960–1975. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Chen, X.; Hong, Y.; Wang, Y.; Ping, X.; Ke, S.; Liu, H.; Zhu, J.; Oliver, D.; Xiang, C. Activated expression of an Arabidopsis HD-START protein confers drought tolerance with improved root system and reduced stomatal density. Plant Cell. 2008, 20, 1134–1151. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis At MYC2 (b HLH) and At MYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell. 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chung, M.; Tu, C. OSTF1: A HD-GL2 family homeobox gene is developmentally regulated during early embryogenesis in rice. Plant Cell Physiol. 2002, 43, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Ingram, G.; Boisnard-Lorig, C.; Dumas, C.; Rogowsky, P. Expression patterns of genes encoding HD-Zip IV homeo domain proteins define specific domains in maize embryos and meristems. Plant J. 2010, 22, 401–414. [Google Scholar] [CrossRef]
- Yadava, P.; Tamim, S.; Zhang, H.; Teng, C.; Zhou, X.; Meyers, B.; Walbot, V. Transgenerational conditioned male fertility of HD-ZIP IV transcription factor mutant ocl4: Impact on 21-nt phasiRNA accumulation in pre-meiotic maize anthers. Plant Reprod. 2021, 34, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, N.; Wu, W.; Bazanova, N.; Reid, N.; Singh, R.; Shirley, N.; Eini, O.; Johnson, A.; Langridge, P.; Hrmova, M.; et al. Wheat wounding-responsive HD-Zip IV transcription factor GL7 is predominantly expressed in grain and activates genes encoding defensins. Plant Mol. Biol. 2019, 101, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, S.; Xiao, Y.; Li, Z.; Wu, M.; Xie, X.; Li, H.; Mu, W.; Li, F.; Liu, P.; et al. Functional characterization of a HD-ZIP IV transcription factor NtHDG2 in regulating flavonols biosynthesis in Nicotiana tabacum. Plant Physiol. Biochem. 2020, 146, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, G.; Pan, J.; Chen, Y.; Xiao, T.; Zhang, L.; Zhang, K.; Wen, H.; Xiong, L.; Yu, Y.; et al. The HD-ZIP IV transcription factor Tril regulates fruit spine density through gene dosage effects in cucumber. J. Exp. Bot. 2020, 71, 6297–6310. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wei, Q.; Liao, Y.; Song, H.; Li, X.; Xiang, C.; Kuai, B. Ectopic overexpression of At HDG11 in tall fescue resulted in enhanced tolerance to drought and salt stress. Plant Cell Rep. 2009, 28, 579–588. [Google Scholar] [CrossRef]
- Wei, Q.; Song, H.; Xiang, C. Preliminary study on improving drought tolerance of ryegrass(Lolium perenne) by overexpressing a HD-START transcription factor At HDG11 from Arabidopsis. Plant Physiol. Commun. 2010, 46, 1140–1146. [Google Scholar] [CrossRef]
- Yu, L.; Wu, S.; Peng, Y.; Liu, R.; Xiang, C. Arabidopsis EDT1/HDG11 improves drought and salt tolerance in cotton and poplar and increases cotton yield in the field. Plant Biotechnol. J. 2016, 14, 72–84. [Google Scholar] [CrossRef]
- Li, L.; Zheng, M.; Deng, G.; Liang, J.; Zhang, H.; Pan, Z.; Long, H.; Yu, M. Overexpression of AtHDG11 enhanced drought tolerance in wheat (Triticum aestivum L.). Mol. Breed. 2016, 36, 23. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, B.; Xu, X.; Chen, H.; Zou, L.; Chen, G.; Cao, B.; Chen, C.; Lei, J. Overexpression of AtEDT1/HDG11 in Chinese Kale (Brassica oleracea var. alboglabra) enhances drought and osmotic stress tolerance. Front. Plant Sci. 2016, 7, 1285. [Google Scholar] [CrossRef] [Green Version]
- Banavath, J.; Chakradhar, T.; Pandit, V.; Konduru, S.; Guduru, K.; Akila, C.; Podha, S.; Puli, C. Stress inducible overexpression of AtHDG11 leads to improved drought and salt stress tolerance in peanut (Arachis hypogaea L.). Front. Chem. 2018. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, Y.; Zhao, P.; Xu, P.; Yu, G.; Zhang, L.; Xiong, Y.; Xiang, C. AtEDT1/HDG11 regulates stomatal density and water-use efficiency via ERECTA and E2Fa. New Phytologist. 2019, 223, 1478–1488. [Google Scholar] [CrossRef]
- Xiong, L.; Wang, R.; Mao, G.; Koczan, J. Identification of drought tolerance determinants by genetic analysis of root response to drought stress and abscisic Acid. Plant Physiol. 2006, 142, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Fang, P.; Yan, H.; Chen, F.; Peng, Y. Overexpression of maize ZmHDZIV14 increases abscisic acid sensitivity and mediates drought and salt stress in Arabidopsis and Tobacco. Plant Mol. Biol. Report. 2020, 39, 275–287. [Google Scholar] [CrossRef]
- Clough, S.; Bent, A. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Nauš, J.; Šmecko, S.; Špundová, M. Chloroplast avoidance movement as a sensitive indicator of RWC during leaf desiccation in the dark. Photosynth. Res. 2016, 129, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X. Experimental Guidance of Plant Physiology; South China University of Technology Press: Guangzhou, China, 2002. [Google Scholar]
- Scarpeci, T.E.; Zanor, M.I.; Mueller-Roeber, B.; Valle, E.M. Overexpression of AtWRKY30 enhances abiotic stress tolerance during early growth stages in Arabidopsis thaliana. Plant Mol. Biol. 2013, 83, 265–277. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.; Cho, D.; Park, J.; Kim, S. ABF2, an ABRE-binding b ZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, Os MYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Wu, Y.; Zhang, Y.; Liu, L.; Ning, Y.; Wang, D.; Tong, H.; Chen, S.; Chu, C.; Xie, Q. Os SDIR1 overexpression greatly improves drought tolerance in transgenic rice. Plant Mol. Biol. 2011, 76, 145–156. [Google Scholar] [CrossRef]
- Hu, T.; Ye, J.; Tao, P.; Li, H.; Zhang, J.; Zhang, Y.; Ye, Z. The tomato HD-Zip I transcription factor Sl HZ24 modulates ascorbate accumulation through positive regulation of the D-mannose/L-galactose pathway. Plant J. 2016, 85, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Ding, Y.; Hu, S.; Ding, L.; Chen, Z.; Zhu, C. The role of HD-Zip class I transcription factors in plant response to abiotic stresses. Physiol. Plant. 2019. [Google Scholar] [CrossRef]
- Liu, J.; Shuai, P. Research progress on HD-ZIP transcription factors in response to pathogenic or abiotic stresses. Fujian J. Agric. Sci. 2021, 36, 124–134. [Google Scholar] [CrossRef]
- Nakamura, M.; Katsumata, H.; Abe, M.; Yabe, N.; Kome da, Y.; Yamamoto, K.; Takahashi, T. Characterization of the class IV homeodomain-Leucine Zipper gene family in Arabidopsis. Plant Physiol. 2006, 141, 1363–1375. [Google Scholar] [CrossRef] [Green Version]
- Sahu, B.; Shaw, B. Isolation, identification and expression analysis of salt-induced genes in Suaeda maritima, a natural halophyte, using PCR-based suppression subtractive hybridization. BMC Plant Biol. 2009, 5, 69–94. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xu, B.; Chen, X.; Wang, L. The effects of increased expression of an Arabidopsis HD-ZIP gene on leaf morphogenesis and anther dehiscence. Plant Sci. 2007, 173, 567–576. [Google Scholar] [CrossRef]
- Gagoet, G.; Almoguera, C.; Jordano, J.; Gonzalez, D.; Chan, R. Hahb-4, a Homeobox-Leucine zipper gene potentially involved in abscisic acid-dependent responses to water stress in sunflower plant. Cell Environ. 2002, 25, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ma, Q.; Jin, X.; Peng, X.; Liu, J.; Lin, D.; Yan, H.; Lei, S.; Jiang, H.; Cheng, B. A novel maize homeodomain-leucine zipper (HDZip)I gene, ZmHDZ10, positively regulates drought and salt tolerance in both rice and Arabidopsis. Plant Cell Physiol. 2014, 55, 1142–1156. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhou, W.; Gong, X.; Cheng, B. Expression of ZmHDZ4, a maize homeodomain-leucine zipper I gene, confers tolerance to drought stress in transgenic rice. Plant Mol. Biol. Report. 2016, 34, 845–853. [Google Scholar] [CrossRef]
- Zhang, S.; Haider, I.; Kohlen, W.; Li, J.; Ouwerkerk, P. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Armengaud, P.; Thiery, L.; Buhot, N.; March, G.; Savouré, A. Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol. Plant. 2004, 120, 442–450. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, J.; Cao, G.; Xie, Y.; Liu, X.; Lu, M.; Wang, G. Overexpression of a PLDalpha1 gene from Setaria italica enhances the sensitivity of Arabidopsis to abscisic acid and improves its drought tolerance. Plant Cell Rep. 2010, 29, 793–802. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene expression and signal transduction in water-stress response. Plant Physiol. 1997, 115, 327–334. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Yan, H.; Fang, P.; Ji, X.; Peng, Y. Heterologous Overexpression of ZmHDZIV13 Enhanced Drought and Salt Tolerance in Arabidopsis and Tobacco. Agronomy 2022, 12, 2378. https://doi.org/10.3390/agronomy12102378
Wang F, Yan H, Fang P, Ji X, Peng Y. Heterologous Overexpression of ZmHDZIV13 Enhanced Drought and Salt Tolerance in Arabidopsis and Tobacco. Agronomy. 2022; 12(10):2378. https://doi.org/10.3390/agronomy12102378
Chicago/Turabian StyleWang, Fang, Huiping Yan, Peng Fang, Xiangzhuo Ji, and Yunling Peng. 2022. "Heterologous Overexpression of ZmHDZIV13 Enhanced Drought and Salt Tolerance in Arabidopsis and Tobacco" Agronomy 12, no. 10: 2378. https://doi.org/10.3390/agronomy12102378
APA StyleWang, F., Yan, H., Fang, P., Ji, X., & Peng, Y. (2022). Heterologous Overexpression of ZmHDZIV13 Enhanced Drought and Salt Tolerance in Arabidopsis and Tobacco. Agronomy, 12(10), 2378. https://doi.org/10.3390/agronomy12102378