
Evaluation of the Composition and Accumulation Pattern of Fatty Acids in Tartary Buckwheat Seed at the Germplasm Level

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Chemicals and Reagents
2.3. Determination of Total Flavonoid Content
2.4. Lipid Extraction and Quantification
2.5. Identification and Expression Analysis of Fatty Acid Biosynthesis Genes in Tartary Buckwheat
2.6. Statistical Analysis
3. Results and Discussion
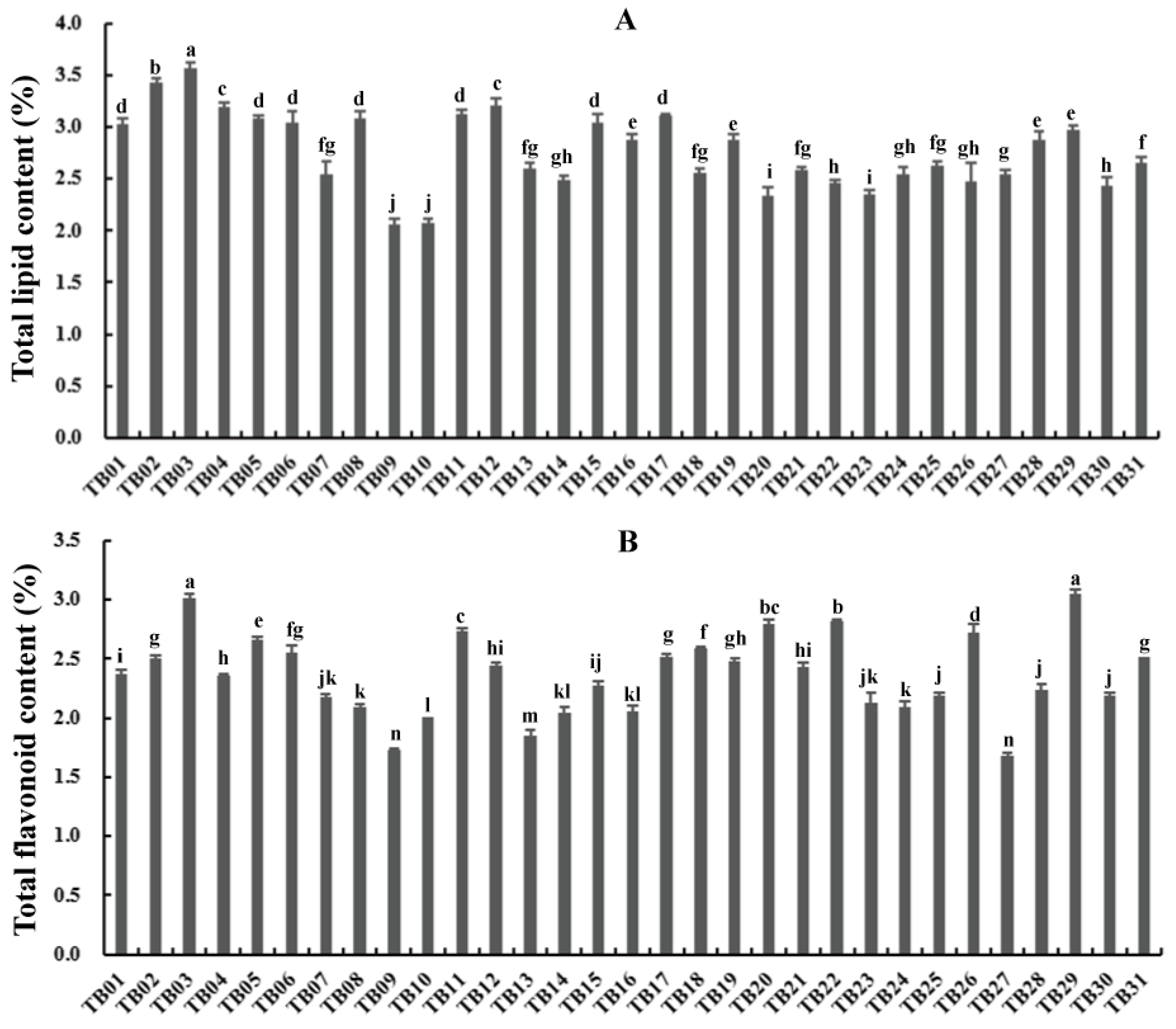
3.1. Total Lipid and Flavonoid Content among Different Tartary Buckwheat Accessions
3.2. Composition and Content of Fatty Acids among Different Tartary Buckwheat Accessions
3.3. Identification of Tartary Buckwheat Germplasms with High USFAs and High Flavonoid Content
3.4. Accumulation Patterns of Fatty Acids in the Developing Seeds of Tartary Buckwheat
3.5. Relative Expression of Fatty Acid Biosynthesis Genes in Tartary Buckwheat
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sánchez-Salcedo, E.M.; Sendra, E.; Carbonell-Barrachina, Á.A.; Martínez, J.J.; Hernández, F. Fatty acids composition of Spanish black (Morus nigra L.) and white (Morus alba L.) mulberries. Food Chem. 2016, 190, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Piskernik, S.; Vidrih, R.; Demšar, L.; Koron, D.; Rogelj, M.; Žontar, T.P. Fatty acid profiles of seeds from different Ribes species. LWT Food Sci. Technol. 2018, 98, 424–427. [Google Scholar] [CrossRef]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. Jpen-Parenter. Enter. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Capouchová, I.; Koŭrimskŭ, L.; Pazderů, L.; Škvorová, P.; Božik, M.; Konvalina, P.; Petr Dvořaká, P.; Dvořáček, V. Fatty acid profile of new oat cultivars grown via organic and conventional farming. J. Cereal Sci. 2021, 98, 103180. [Google Scholar] [CrossRef]
- Montaner, C.; Zufiaurre, R.; Movila, M.; Mallor, C. Evaluation of borage (Borago officinalis L.) genotypes for nutraceutical value based on leaves fatty acids composition. Foods 2022, 11, 16. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Ma, B.; Gao, Q.; Du, H.; Han, Y.; Li, Y.; Cao, Y.; Qi, M.; Zhu, Y.; et al. The Tartary buckwheat genome provides insights into rutin biosynthesis and abiotic stress tolerance. Mol. Plant 2017, 10, 1224–1237. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Lv, Q.; Liu, A.; Wang, J.; Sun, X.; Deng, J.; Chen, Q.; Wu, Q. Comparative metabolomics study of Tartary (Fagopyrum tataricum (L.) Gaertn) and common (Fagopyrum esculentum Moench) buckwheat seeds. Food Chem. 2022, 371, 131125. [Google Scholar] [CrossRef]
- Li, H.Y.; Wu, C.X.; Lv, Q.Y.; Shi, T.X.; Chen, Q.J.; Chen, Q.F. Comparative cellular, physiological and transcriptome analyses reveal the potential easy dehulling mechanism of rice-tartary buckwheat (Fagopyrum Tararicum). BMC Plant Biol. 2020, 20, 505. [Google Scholar] [CrossRef]
- Huda, M.N.; Lu, S.; Jahan, T.; Ding, M.; Jha, R.; Zhang, K.; Zhang, W.; Georgiev, M.I.; Park, S.U.; Zhou, M. Treasure from garden: Bioactive compounds of buckwheat. Food Chem. 2021, 335, 127653. [Google Scholar] [CrossRef]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef]
- Qin, P.Y.; Wang, Q.A.; Shan, F.; Hou, Z.H.; Ren, G.X. Nutritional composition and flavonoids content of flour from different buckwheat cultivars. Int. J. Food Sci. Tech. 2010, 45, 951–958. [Google Scholar]
- Pirzadah, T.B.; Malik, B.; Tahir, I.; Ul Rehman, R. Buckwheat journey to functional food sector. Curr. Nutr. Food Sci. 2019, 15, 1–8. [Google Scholar]
- Tsuzuki, W.; Ogata, Y.; Akasaka, K.; Shibata, S.; Suzuki, T. Fatty acid composition of selected buckwheat species by fluorometric high-performance liquid chromatography. Cereal Chem. 1991, 68, 365–369. [Google Scholar]
- Zhou, Y.; Hong, W.; Cui, L.; Zhou, X.; Wen, T.; Song, X. Evolution of nutrient ingredients in tartary buckwheat seeds during germination. Food Chem. 2015, 186, 244–248. [Google Scholar]
- Peng, L.X.; Zou, L.; Tan, M.L.; Deng, Y.Y.; Zhao, G. Free amino acids, fatty acids, and phenolic compounds in tartary buckwheat of different hull colour. Czech J. Food Sci. 2017, 35, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Dziedzic, K.; Kurek, S.; Mildner-Szkudlarz, S.; Ivan Kreft, I.; Walkowiak, J. Fatty acids profile, sterols, tocopherol and squalene content in Fagopyrum tataricum seed milling fractions. J. Cereal Sci. 2020, 96, 103118. [Google Scholar] [CrossRef]
- Sinkovič, L.; Kokalj Sinkovič, D.; Meglič, V. Milling fractions composition of common (Fagopyrum esculentum Moench) and Tartary (Fagopyrum tataricum (L.) Gaertn.) buckwheat. Food Chem. 2021, 365, 130459. [Google Scholar]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Huang, Y.; Sun, Z.; Huang, J.; Wang, Z. Transcriptome analysis of genes involved in lipid biosynthesis in the developing embryo of pecan (Carya illinoinensis). J. Agric. Food Chem. 2017, 65, 4223–4236. [Google Scholar] [CrossRef]
- Hu, X.D.; Pan, B.Z.; Fu, Q.; Niu, L.; Chen, M.S.; Xu, Z.F. De novo transcriptome assembly of the eight major organs of Sacha Inchi (Plukenetia volubilis) and the identification of genes involved in α-linolenic acid metabolism. BMC Genom. 2018, 19, 380. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, A.; Shanklin, J.; Whittle, E.; Lapchyk, L.; Hildebrand, D.; Kachroo, P. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 2007, 63, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zhou, X.R.; Wood, C.C.; Green, A.G.; Singh, S.P.; Liu, L.; Liu, Q. A large and functionally diverse family of Fad2 genes in safflower (Carthamus tinctorius L.). BMC Plant Biol. 2013, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslam, R.P.; Sayanova, O.; Kim, H.J.; Cahoon, E.B.; Napier, J.A. Synthetic redesign of plant lipid metabolism. Plant J. 2016, 87, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Miao, L.; He, J.; Zhang, K.; Li, Y.; Gai, J. Dynamic transcriptome changes related to oil accumulation in developing soybean seeds. Int. J. Mol. Sci. 2019, 20, 2202. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Xiang, D.B.; Yan, L.; Song, Y.; Zhao, G.; Wang, Y.H.; Zhang, B.L. Changes in seed growth, levels and distribution of flavonoids during tartary buckwheat seed development. Plant Prod. Sci. 2016, 19, 1–10. [Google Scholar] [CrossRef]
- Saoussem, H.; Sadok, B.; Habib, K.; Mayer, P.M. Fatty acid accumulation in the different fractions of the developing corn kernel. Food Chem. 2009, 117, 432–437. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wu, B.; Cao, X.; Liu, H.; Zhu, C.; Klee, H.; Zhang, B.; Chen, K. UDP-glucosyltransferase PpUGT85A2 controls volatile glycosylation in peach. J. Exp. Bot. 2019, 70, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yin, X.; Xiao, Y.; Zhang, Z.; Li, S.; Liu, X.; Zhang, B.; Yang, X.; Grierson, D.; Jiang, G.; et al. An ethylene response Factor-MYB transcription complex regulates furaneol biosynthesis by activating quinone oxidoreductase expression in strawberry. Plant Physiol. 2018, 178, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Narayan, B.; Miyashita, K.; Hosakawa, M. Physiological effects of eicosapentaenoic acid (epa) and docosahexaenoic acid (dha)-a review. Food Rev. Int. 2006, 22, 291–307. [Google Scholar] [CrossRef]
- Wang, R.; Shu, P.; Zhang, C.; Zhang, J.; Chen, Y.; Zhang, Y.; Du, K.; Xie, Y.; Li, M.; Ma, T.; et al. Integrative analyses of metabolome and genome-wide transcriptome reveal the regulatory network governing flavor formation in kiwifruit (Actinidia chinensis). New Phytol. 2022, 233, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shen, S.; Zhou, S.; Li, Y.; Mao, Y.; Zhou, J.; Shi, Y.; An, L.; Zhou, Q.; Peng, W.; et al. Rice metabolic regulatory network spanning the entire life cycle. Mol. Plant 2022, 15, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.; Zhou, L.; You, S.; Deng, H.; Chen, Y.; Alseekh, S.; Yuan, Y.; Fu, R.; Zhang, Z.; et al. MicroTom metabolic network: Rewiring tomato metabolic regulatory network throughout the growth cycle. Mol. Plant 2020, 13, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, X.D.; Rose, R.J. Oil body biogenesis and biotechnology in legume seeds. Plant Cell Rep. 2017, 36, 1519–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of wild olive and the evolution of oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 | C20:0 | C20:4 | C20:5 | C22:6 |
|---|---|---|---|---|---|---|---|---|---|---|
| TB01 | 3150.31 ± 30.39 def | 1814.18 ± 3.83 e | 864.94 ± 11.33 c | 7642.43 ± 129.45 f | 4562.48 ± 74.67 g | 152.38 ± 2.64 hij | 10.57 ± 0.44 fgh | 40.15 ± 1.52 de | 29.68 ± 2.19 f | 32.29 ± 0.15 a |
| TB02 | 3728.28 ± 32.96 b | 2300.62 ± 27.83 a | 899.34 ± 34.54 bc | 9542.63 ± 88.21 a | 4936.79 ± 129.04 f | 182.64 ± 2.10 b | 9.83 ± 0.59 fghij | 55.85 ± 0.72 b | 21.20 ± 0.58 mn | 14.26 ± 0.32 e |
| TB03 | 4365.18 ± 119.05 a | 1857.70 ± 14.04 d | 768.09 ± 10.57 efgh | 8060.30 ± 121.74 d | 7470.18 ± 122.89 a | 152.55 ± 1.88 ghij | 13.50 ± 0.27 a | 40.49 ± 1.79 de | 35.74 ± 0.62 bc | 10.84 ± 0.81 o |
| TB04 | 2408.99 ± 125.68 i | 1750.46 ± 8.43 f | 801.15 ± 15.90 de | 7800.89 ± 207.61 e | 6274.17 ± 90.66 c | 154.63 ± 2.28 ghi | 10.65 ± 0.70 efgh | 41.76 ± 2.61 d | 32.11 ± 1.01 e | 10.52 ± 0.15 hi |
| TB05 | 2927.97 ± 38.58 fgh | 1652.66 ± 15.81 h | 1140.16 ± 19.94 a | 7085.88 ± 79.96 k | 5280.88 ± 27.97 d | 150.58 ± 0.75 ij | 10.96 ± 0.94 def | 36.97 ± 2.67 fg | 25.23 ± 0.70 hij | 10.11 ± 0.16 hijk |
| TB06 | 3397.59 ± 130.13 bcde | 1789.61 ± 25.50 e | 766.17 ± 42.25 efgh | 7599.66 ± 115.58 fg | 4155.35 ± 50.59 j | 156.96 ± 3.66 fgh | 8.29 ± 0.87 klmno | 34.61 ± 1.26 gh | 19.28 ± 0.18 o | 17.48 ± 1.03 b |
| TB07 | 2886.19 ± 66.96 fgh | 1458.59 ± 12.34 k | 514.83 ± 12.03 op | 6100.08 ± 52.82 p | 3143.83 ± 88.63 l | 125.25 ± 1.46 no | 9.98 ± 0.93 fghi | 29.47 ± 1.76 ijk | 20.64 ± 1.44 no | 9.90 ± 0.55 jk |
| TB08 | 2689.99 ± 63.59 ghi | 1891.48 ± 30.40 cd | 790.74 ± 23.31 def | 8205.58 ± 98.56 c | 4288.01 ± 28.87 hij | 157.14 ± 0.82 fg | 10.88 ± 0.74 defg | 46.02 ± 0.38 c | 23.44 ± 0.93 jkl | 11.72 ± 0.58 g |
| TB09 | 1972.57 ± 32.14 j | 978.25 ± 2.78 o | 357.61 ± 7.87 s | 4032.12 ± 63.72 s | 4191.48 ± 25.92 ij | 91.40 ± 1.40 q | 10.26 ± 0.75 fghi | 32.07 ± 1.41 hi | 16.15 ± 0.47 p | 11.54 ± 0.89 g |
| TB10 | 2465.97 ± 91.77 i | 1014.42 ± 11.07 n | 421.17 ± 16.55 r | 4468.57 ± 127.71 r | 2750.06 ± 92.62 m | 87.28 ± 0.58 r | 9.46 ± 0.52 ghijkl | 28.15 ± 0.89 jk | 17.16 ± 0.70 p | 11.54 ± 0.97 g |
| TB11 | 2697.63 ± 58.23 ghi | 1799.32 ± 8.42 e | 730.00 ± 24.74 hij | 7745.30 ± 111.37 e | 5319.85 ± 198.46 d | 154.00 ± 1.52 ghi | 11.99 ± 0.61 bcde | 38.47 ± 0.13 ef | 25.83 ± 0.84 hi | 16.54 ± 1.49 c |
| TB12 | 2963.72 ± 26.80 fgh | 2140.70 ± 25.69 b | 908.80 ± 10.19 b | 8516.75 ± 169.08 b | 5268.77 ± 118.82 de | 194.45 ± 2.66 a | 12.51 ± 0.79 ab | 61.95 ± 2.28 a | 40.84 ± 1.12 a | 15.59 ± 0.73 d |
| TB13 | 3522.20 ± 90.52 bcd | 1797.68 ± 9.87 e | 745.81 ± 36.13 ghi | 7219.24 ± 124.53 j | 6194.15 ± 106.30 c | 159.77 ± 1.72 ef | 8.09 ± 0.14 lmno | 28.88 ± 1.37 ijk | 21.74 ± 0.52 lmn | 11.93 ± 0.32 g |
| TB14 | 2609.82 ± 51.96 hi | 1707.90 ± 46.85 g | 667.59 ± 16.55 kl | 7087.59 ± 172.56 k | 5462.93 ± 213.60 d | 148.40 ± 3.98 jk | 7.33 ± 0.63 no | 29.56 ± 1.41 ijk | 25.20 ± 0.89 hij | 9.57 ± 0.27 k |
| TB15 | 3274.02 ± 111.10 cdef | 1631.89 ± 14.16 h | 695.82 ± 7.37 jk | 7024.55 ± 154.66 k | 5415.26 ± 105.32 d | 136.23 ± 1.45 l | 12.23 ± 0.34 abcd | 27.71 ± 0.42 k | 26.72 ± 0.79 gh | 10.39 ± 0.23 hij |
| TB16 | 3399.51 ± 109.55 bcde | 1598.09 ± 7.51 i | 617.50 ± 21.06 mn | 6575.30 ± 86.40 m | 4185.33 ± 42.16 ij | 144.95 ± 1.93 k | 8.47 ± 0.76 jklm | 27.25 ± 0.22 kl | 32.75 ± 2.88 de | 9.97 ± 0.58 ijk |
| TB17 | 3020.24 ± 35.03 efg | 1883.12 ± 13.36 cd | 720.72 ± 33.16 ij | 7532.33 ± 76.51 gh | 5049.41 ± 43.05 ef | 167.69 ± 1.32 d | 12.89 ± 1.30 ab | 47.74 ± 3.07 c | 36.46 ± 1.32 b | 11.92 ± 0.99 g |
| TB18 | 2890.68 ± 22.84 fgh | 1803.47 ± 19.31 e | 653.46 ± 11.16 lm | 7413.87 ± 136.14 i | 6562.06 ± 221.03 b | 152.76 ± 0.95 ghij | 12.35 ± 0.35 abc | 48.00 ± 0.79 c | 34.14 ± 1.06 cd | 14.41 ± 0.45 e |
| TB19 | 3069.49 ± 61.63 efg | 1691.33 ± 19.25 g | 642.03 ± 8.71 lm | 6914.16 ± 75.14 l | 4242.45 ± 110.19 hij | 159.67 ± 0.76 ef | 8.30 ± 0.21 klmno | 36.67 ± 1.67 fg | 24.01 ± 0.88 ijk | 15.43 ± 0.32 d |
| TB20 | 3018.30 ± 55.70 efg | 1353.03 ± 26.45 m | 899.48 ± 29.31 bc | 5747.21 ± 108.68 q | 4447.26 ± 99.56 gh | 126.66 ± 4.98 no | 7.51 ± 0.43 mno | 17.29 ± 0.43 o | 25.62 ± 0.34 hi | 8.15 ± 0.11 l |
| TB21 | 2571.08 ± 130.10 hi | 1864.28 ± 7.94 d | 780.23 ± 27.79 defg | 7445.83 ± 212.14 hi | 6298.93 ± 67.50 c | 172.74 ± 1.73 c | 12.40 ± 0.59 abc | 37.69 ± 1.54 efg | 26.44 ± 1.02 gh | 9.60 ± 0.26 k |
| TB22 | 3515.69 ± 92.57 bcd | 1515.19 ± 38.35 j | 808.31 ± 13.31 d | 6233.75 ± 110.48 o | 5342.70 ± 89.10 d | 139.94 ± 0.49 l | 9.31 ± 0.16 hijkl | 21.94 ± 1.06 mn | 27.71 ± 1.16 g | 14.12 ± 0.23 ef |
| TB23 | 2036.43 ± 39.04 j | 1342.38 ± 28.36 m | 510.21 ± 10.52 op | 6023.54 ± 75.10 p | 4871 ± 132.75 f | 115.68 ± 2.05 p | 9.36 ± 0.39 hijkl | 16.93 ± 0.62 o | 21.38 ± 0.65 mn | 6.79 ± 0.33 n |
| TB24 | 2882.03 ± 41.69 fgh | 1476.40 ± 15.32 k | 756.90 ± 23.87 fghi | 6403.95 ± 65.11 n | 4454.38 ± 99.22 gh | 122.47 ± 1.42 o | 7.70 ± 0.07 mno | 21.91 ± 1.43 mn | 22.81 ± 0.29 klm | 13.65 ± 0.43 f |
| TB25 | 3209.27 ± 24.31 cdef | 1913.88 ± 49.91 c | 918.90 ± 21.99 b | 7783.30 ± 122.16 e | 3805.38 ± 113.09 k | 169.55 ± 2.14 cd | 8.36 ± 0.57 klmno | 24.60 ± 0.06 lm | 33.17 ± 0.21 de | 9.76 ± 0.45 k |
| TB26 | 2708.32 ± 92.59 ghi | 1386.51 ± 26.28 l | 467.33 ± 12.60 q | 5671.66 ± 120.52 q | 3685.49 ± 78.57 k | 131.53 ± 3.09 m | 7.46 ± 0.75 mno | 20.53 ± 0.92 n | 24.98 ± 0.40 hij | 10.58 ± 0.41 k |
| TB27 | 2668.47 ± 80.37 ghi | 1809.49 ± 11.87 e | 641.56 ± 17.50 lm | 7450.68 ± 32.61 hi | 4223.56 ± 170.88 hij | 155.18 ± 2.82 ghi | 8.84 ± 0.65 ijklm | 23.96 ± 1.11 m | 33.15 ± 1.17 de | 15.19 ± 0.93 d |
| TB28 | 4112.55 ± 10.00 a | 1516.54 ± 21.30 j | 533.74 ± 9.59 o | 6450.25 ± 120.36 n | 4410.52 ± 28.78 ghi | 128.62 ± 0.50 mn | 9.74 ± 0.35 fghijk | 22.36 ± 1.58 mn | 32.49 ± 0.53 de | 6.45 ± 0.11 n |
| TB29 | 3560.48 ± 48.66 bc | 1799.15 ± 14.96 e | 597.32 ± 15.95 n | 7257.85 ± 134.73 j | 3781.57 ± 112.04 k | 162.03 ± 0.95 ef | 6.95 ± 0.09 o | 31.07 ± 0.93 ij | 24.94 ± 0.23 hij | 7.57 ± 0.67 m |
| TB30 | 2919.97 ± 42.04 fgh | 1457.50 ± 52.23 k | 490.08 ± 35.15 pq | 6559.74 ± 211.69 m | 2781.39 ± 72.33 m | 116.70 ± 9.74 p | 12.83 ± 0.07 ab | 28.62 ± 0.77 jk | 28.02 ± 0.39 fg | 13.92 ± 0.56 ef |
| TB31 | 3258.24 ± 85.51 cdef | 1868.62 ± 24.54 d | 786.34 ± 44.47 def | 7761.08 ± 41.28 e | 4384.30 ± 121.07 ghij | 169.87 ± 1.86 cd | 11.06 ± 0.81 cdef | 31.10 ± 1.49 ij | 40.07 ± 1.25 a | 15.77 ± 0.71 d |
| Total Flavonoid (%) | OA (μg g−1) | LA (μg g−1) | ALA (μg g−1) | AA (μg g−1) | ARA (μg g−1) | EPA (μg g−1) | DHA (μg g−1) |
|---|---|---|---|---|---|---|---|
| TB29 (3.04 ± 0.05) | TB02 (9542.63 ± 88.21) | TB03 (7470.18 ± 122.89) | TB12 (194.45 ± 2.66) | TB03 (13.50 ± 0.27) | TB12 (61.95 ± 2.28) | TB12 (40.84 ± 1.12) | TB01 (32.29 ± 0.15) |
| TB03 (3.01 ± 0.04) | TB12 (8516.75 ± 169.08) | TB18 (6562.06 ± 221.03) | TB02 (182.64 ± 2.10) | TB17 (12.89 ± 1.30) | TB02 (55.85 ± 0.72) | TB31 (40.07 ± 1.25) | TB06 (17.48 ± 1.03) |
| TB22 (2.81 ± 0.02) | TB08 (8205.58 ± 98.56) | TB21 (6298.93 ± 67.50) | TB21 (172.74 ± 1.73) | TB30 (12.83 ± 0.07) | TB18 (48.00 ± 0.79) | TB17 (36.46 ± 1.32) | TB11 (16.54 ± 1.49) |
| TB20 (2.79 ± 0.05) | TB03 (8060.30 ± 121.74) | TB04 (6274.17 ± 90.66) | TB31 (169.87 ± 1.86) | TB12 (12.51 ± 0.79) | TB17 (47.74 ± 3.07) | TB03 (35.74 ± 0.62) | TB31 (15.77 ± 0.71) |
| TB11 (2.73 ± 0.03) | TB04 (7800.89 ± 207.61) | TB13 (6194.15 ± 106.30) | TB25 (169.55 ± 2.14) | TB21 (12.40 ± 0.59) | TB08 (46.02 ± 0.38) | TB18 (34.14 ± 1.06) | TB12 (15.59 ± 0.73) |
| TB26 (2.72 ± 0.08) | TB25 (7783.30 ± 122.16) | TB14 (5462.93 ± 213.60) | TB17 (167.69 ± 1.32) | TB18 (12.35 ± 0.35) | TB04 (41.76 ± 2.61) | TB25 (33.17 ± 0.21) | TB19 (15.43 ± 0.32) |
| TB05 (2.66 ± 0.02) | TB31 (7761.08 ± 41.28) | TB15 (5415.26 ± 105.32) | TB29 (162.03 ± 0.95) | TB15 (12.23 ± 0.34) | TB03 (40.49 ± 1.79) | TB27 (33.15 ± 1.17) | TB27 (15.19 ± 0.93) |
| TB18 (2.58 ± 0.04) | TB11 (7745.30 ± 111.37) | TB22 (5342.70 ± 89.10) | TB13 (159.77 ± 1.72) | TB11 (11.99 ± 0.61) | TB01 (40.15 ± 1.52) | TB16 (32.75 ± 2.88) | TB18 (14.41 ± 0.45) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Q.; Wang, J.; Sun, P.; Cai, F.; Ran, B.; Deng, J.; Shi, T.; Chen, Q.; Li, H. Evaluation of the Composition and Accumulation Pattern of Fatty Acids in Tartary Buckwheat Seed at the Germplasm Level. Agronomy 2022, 12, 2447. https://doi.org/10.3390/agronomy12102447
Lv Q, Wang J, Sun P, Cai F, Ran B, Deng J, Shi T, Chen Q, Li H. Evaluation of the Composition and Accumulation Pattern of Fatty Acids in Tartary Buckwheat Seed at the Germplasm Level. Agronomy. 2022; 12(10):2447. https://doi.org/10.3390/agronomy12102447
Chicago/Turabian StyleLv, Qiuyu, Jiarui Wang, Peiyuan Sun, Fang Cai, Bin Ran, Jiao Deng, Taoxiong Shi, Qingfu Chen, and Hongyou Li. 2022. "Evaluation of the Composition and Accumulation Pattern of Fatty Acids in Tartary Buckwheat Seed at the Germplasm Level" Agronomy 12, no. 10: 2447. https://doi.org/10.3390/agronomy12102447
APA StyleLv, Q., Wang, J., Sun, P., Cai, F., Ran, B., Deng, J., Shi, T., Chen, Q., & Li, H. (2022). Evaluation of the Composition and Accumulation Pattern of Fatty Acids in Tartary Buckwheat Seed at the Germplasm Level. Agronomy, 12(10), 2447. https://doi.org/10.3390/agronomy12102447