
LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, In Vitro Establishment, Rooting and Acclimatization
2.2. Culture Conditions and Light Quality
2.3. Data Collection
2.4. Statistical Analysis
3. Results
3.1. Morphological and Physiological Traits
3.2. Photosynthetic Pigments
3.3. Protein and Malondialdehyde (MDA) Content
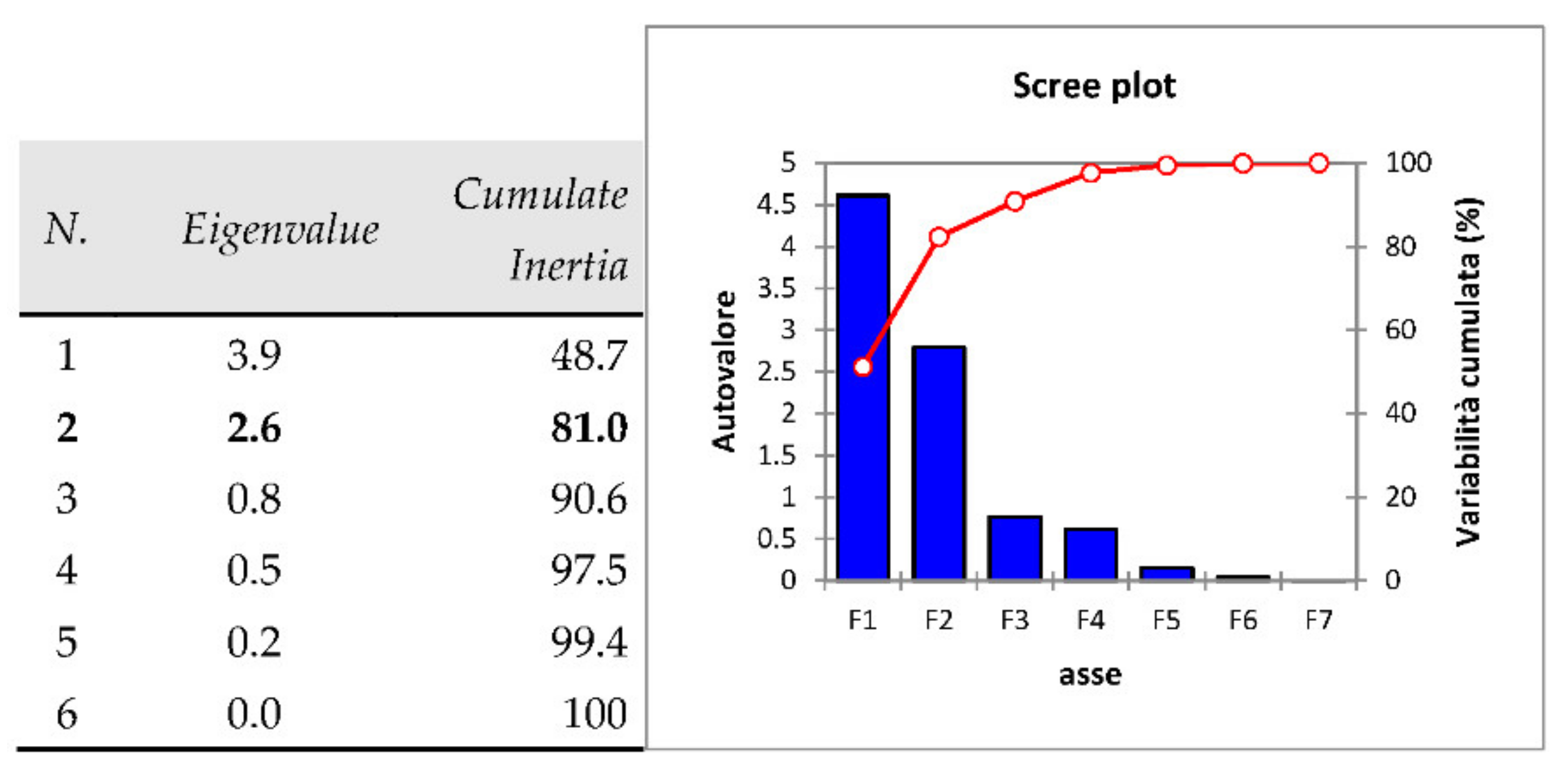
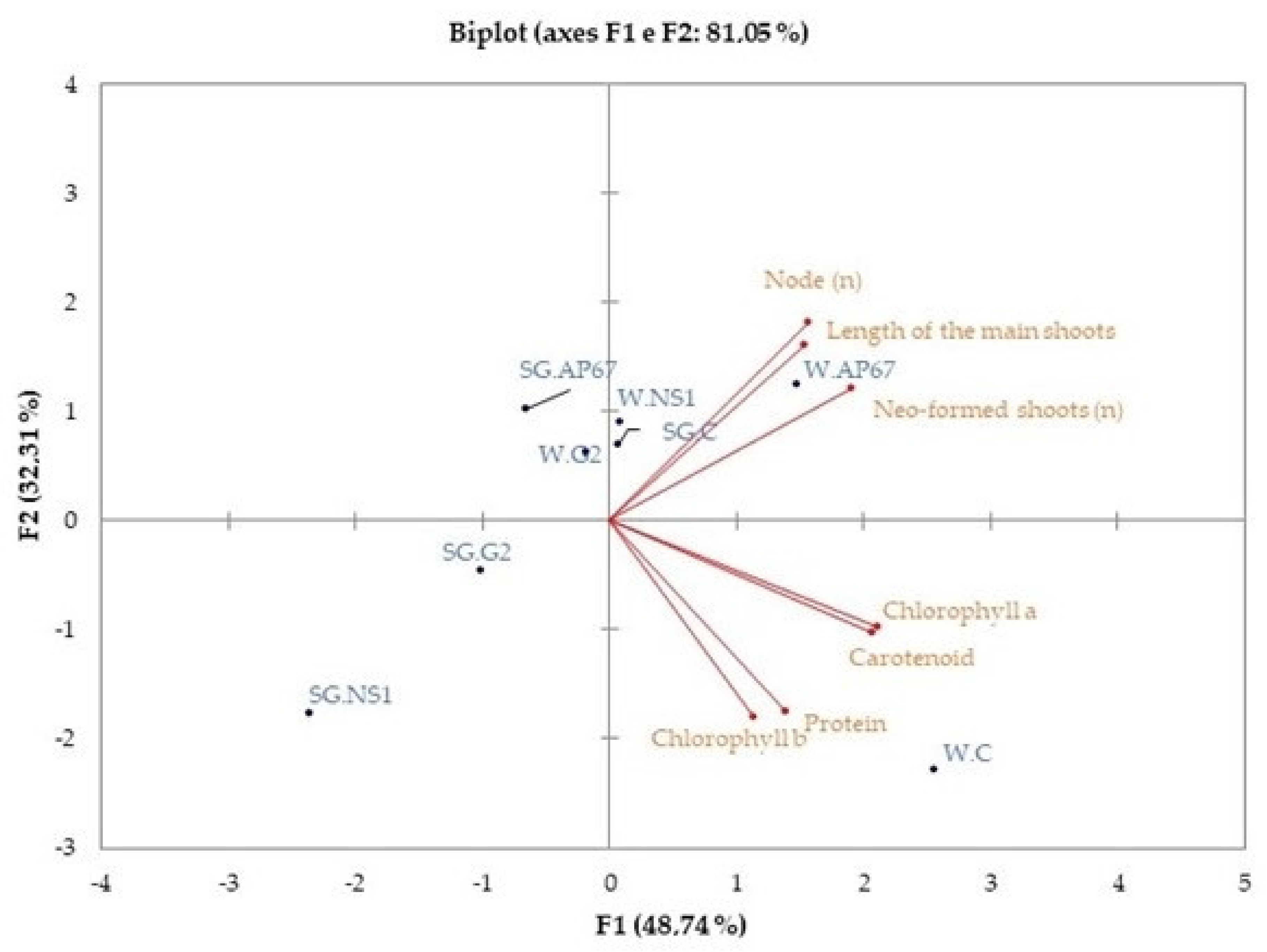
3.4. Principal Component Analysis (PCA)
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lizárraga, A.; Fraga, M.; Ascasíbar, J.; González, M. In Vitro Propagation and Recovery of Eight Apple and Two Pear Cultivars Held in a Germplasm Bank. Am. J. Plant Sci. 2017, 8, 2238–2254. [Google Scholar] [CrossRef] [Green Version]
- Luziatelli, F.; Gatti, L.; Ficca, A.G.; Medori, G.; Silvestri, C.; Melini, F.; Muleo, R.; Ruzzi, M. Metabolites secreted by a plant-growth-promoting Pantoea agglomerans strain improved rooting of Pyrus communis L. cv dar gazi cuttings. Front. Microbiol. 2020, 11, 539359. [Google Scholar] [CrossRef] [PubMed]
- Aygun, A.; Dumanoglu, H. In vitro shoot proliferation and in vitro and ex vitro root formation of Pyruselae agrifolia Pallas. Front. Plant Sci. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.M.; Wada, S.; DeNoma, J.; Niedz, R.P. Mineral nutrition influences physiological responses of pear in vitro. In Vitro Cell Dev. Biol. Plant 2013, 49, 699–709. [Google Scholar] [CrossRef]
- Bell, R.L.; Scorza, R.; Lomberk, D. Adventitious shoot regeneration of pear (Pyrus spp.) genotypes. Plant Cell Tissue Organ Cult. 2012, 108, 229–236. [Google Scholar] [CrossRef]
- USDA ARS. GRIN-Global. Available online: https://www.ars-grin.gov/ (accessed on 15 February 2021).
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Ahlman, L.; Bånkestad, D.; Wik, T. Using chlorophyll a fluorescence gains to optimize LED light spectrum for short term photosynthesis. Comput. Electron. Agric. 2017, 142, 224–234. [Google Scholar] [CrossRef]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the role of red:blue LED lights on resource use efficiency and nutritional properties of indoor grown sweet basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef]
- Schulze, P.S.C.; Barreira, L.A.; Pereira, H.G.C.; Perales, J.A.; Varela, J.C.S. Light emitting diodes (LEDs) applied to microalgal production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Gourrierec, J.G.-L.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Silvestri, C.; Caceres, M.E.; Ceccarelli, M.; Pica, A.L.; Rugini, E.; Cristofori, V. Influence of Continuous Spectrum Light on Morphological Traits and Leaf Anatomy of Hazelnut Plantlets. Front. Plant Sci. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Liu-Gitz, L.; Britz, S.J.; Wergin, W.P. Blue light inhibits stomatal development in soybean isolines containing kaempferol- 3-O-2G-glycosylgentiobioside (K9), a unique flavonoid glycoside. Plant Cell Environ. 2000, 23, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, J.; Qu, L.; Chen, Z.; Zhao, H.; Deng, X.W. Light control of Arabidopsis development entails coordinated regulation of genome expression and cellular pathways. Plant Cell 2001, 13, 2589–2607. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Pack, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Savvides, A.; Fanourakis, D.; van Leperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wei, L.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light and blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Smirnakou, S.; Ouzounis, T.; Radoglou, M. Continuous spectrum LEDs promote seedling quality traits and performance of Quercus ithaburensis var. macrolepis. Front. Plant Sci. 2017, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Van Labeke, M.C. Long-term effects of red- and blue light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Galvao, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Huché-Thélier, L.; Crespel, L.; Gourrierec, J.G.-L.; Morel, P.; Sakr, S.; LeDuc, N. Light signaling and plant responses to blue and UV radiations—Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Ballaré, C.E.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Kotb, O.M.; Abd EL-Latif, F.M.; Atawia, A.R.; Saleh, S.S.; EL-Gioushy, S.F. In vitro Propagation and Callus Induction of Pear (Pyrus communis) Cv. Le-Conte. Asian J. Biotechnol. Genet. Eng. 2020, 3, 1–10. [Google Scholar]
- Xu, J.; Wang, Y.; Zhang, Y.; Chai, T.Y. Rapid in Vitro Multiplication and ex Vitro Rooting of Malus zumi (Matsumura) Rehd. Acta Physiol. Plant 2008, 30, 129–132. [Google Scholar] [CrossRef]
- Erig, A.C.; Fortes, G.R. In Vitro Establishment of Pear (Pyrus spp.) Starting from Meristems and Buds. Cienc. Rural 2002, 32, 577–582. [Google Scholar] [CrossRef]
- Thakur, A.; Kanwar, J.S. Mircropropagation of “Wild Pear” Pyrus pyrifolia (Burm F) Nakai I Explant Establishment and Shoot Multiplication. Not. Bot. Horti Agrobot. Cluj-Napoca 2008, 36, 103–108. [Google Scholar]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Agati, G.; Cerovic, Z.G.; Pinelli, P.; Tattini, M. Light-induced accumulation of ortho-dihydroxylated flavonoids as nondestructively monitored by chlorophyll fluorescence excitation techniques. Environ. Exp. Bot. 2011, 73, 3–9. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Gupta, S.D.; Jatothu, B. Fundamentals and applications of light emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Cavallaro, V.; Pellegrino, A.; Muleo, R.; Forgione, I. Light and Plant Growth Regulators on In Vitro Proliferation. Plants 2022, 11, 844. [Google Scholar] [CrossRef] [PubMed]
- Gnasekaran, P.; Rahman, Z.A.; Chew, B.L.; Appalasamy, S.; Mariappan, V.; Subramaniam, S. Development of micropropagation system of Zingiber officinale var. rubrum Theilade using different spectrum light-emitting diode (LED) irradiation. Ind. Crops Prod. 2021, 170, 113748. [Google Scholar] [CrossRef]
- Miranda, N.A.; Xavier, A.; Otoni, W.C.; Gallo, R.; Gatti, K.C.; De Moura, L.C.; Souza, D.M.S.C.; Maggioni, J.H.; de Santos, S.S.O. Quality, and Intensity of Light in the In Vitro Development of Microstumps of Eucalyptus urophylla in a Photoautotrophic System. For. Sci. 2020, 66, 754–760. [Google Scholar] [CrossRef]
- Oliveira, T.D.R.D.; Aragão, V.P.M.; Moharana, K.C.; Fedosejevs, E.; Amaral, F.P.D.; de Sousa, K.R.; Thelen, J.J.; Venâncio, T.M.; Silveira, V.; Santa-Catarina, C. Light spectra affect the in vitro shoot development of Cedrela fissilis Vell. (Meliaceae) by changing the protein profile and polyamine contents. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140529. [Google Scholar] [CrossRef]
- Smirnakou, S.; Ouzounis, T.; Radoglou, K. Effects of continuous spectrum LEDs used in indoor cultivation of two coniferous species Pinus sylvestris L. and Abies borisii-regis Mattf. Scand. J. For. Res. 2017, 32, 115–122. [Google Scholar] [CrossRef]
- Díaz-Rueda, P.; Cantos-Barragán, M.; Colmenero-Flores, J.M. Growth Quality and Development of Olive Plants Cultured In-Vitro under Different Illumination Regimes. Plants 2021, 10, 2214. [Google Scholar] [CrossRef]
- Silvestri, C.; Rugini, E.; Cristofori, V. The effect of CuSO4 for establishing in vitro culture, and the role nitrogen and iron sources in in vitro multiplication of Corylus avellana L. cv. tonda gentile romana. Plant Biosyst. 2020, 154, 17–23. [Google Scholar] [CrossRef]
- Quoirin, M.; Lepoivre, P.H. Improved media for in vitro culture of Prunus sp. In Symposium on Tissue Culture for Horticultural Purposes; ISHS: Gent, Belgium, 1977; Volume 78, pp. 437–442. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Astolfi, S.; Zuchi, S.; Passera, C. Effect of cadmium on H+ ATPase activity of plasma membrane vesicles isolated from roots of different S-supplied maize (Zea mays L.) plants. Plant Sci. 2005, 169, 361–368. [Google Scholar] [CrossRef]
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E.; Tatham, R.L. Multivariate Data Analysis; Pearson University Press: Upper Saddle River, NJ, USA, 2006; Volume 6, p. 289. [Google Scholar]
- Smith, H. Physiological and ecological function within the phytochrome family. Ann. Rev. Plant Biol. 1995, 46, 289–315. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Murakami, A.; Murakami, K.; Tanaka, M. Sugar-free micropropagation of Eucalyptus citriodora using light-emitting diode (LEDs) and film-rockwool culture system. Environ. Control Biol. 2002, 40, 147–155. [Google Scholar]
- Lian, M.L.; Murthy, H.N.; Paek, K.Y. Effects of light emitting diodes (LED) on the in vitro induction and growth of bulblets of Lilium oriental hybrid ‘Pesaro’. Sci. Hortic. 2002, 94, 365–370. [Google Scholar] [CrossRef]
- Duong, T.N.; Hong, L.T.A.; Watanabe, H.; Goi, M.; Tanaka, M. Efficiency of a novel culture system by using light-emitting diode (LED) on in vitro and subsequent growth of micro propagated banana plantlets. Acta Hortic. 2003, 616, 121–127. [Google Scholar]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green light supplementation for enhanced lettuce growth under red and blue light emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Stomatal conductance of lettuce grown under or exposed to different light qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of Chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Poudel, R.P.; Kataoka, I.; Mochioka, R. Effect of red-and blue-light emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Shin, S.K.; Murthy, N.H.; Heo, W.J.; Hahn, J.E.; Paek, Y.K. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Lee, S.W.; Seo, J.M.; Lee, M.K.; Chun, J.H.; Antonisamy, P.; Valan Arasu, M.; Suzuki, T.; Al-Dhabi, N.A.; Kim, S.J. Influence of different LED lamps on the production of phenolic compounds in common and Tartary buckwheat sprouts. Ind. Crops Prod. 2014, 54, 320–326. [Google Scholar] [CrossRef]
- Ouzounis, T.; Razi Parijkolaei, B.; Fretté, X.; Rosenqvist, E.; Ottosen, C.O. Predawn and high intensity application of supplemental blue light decreases the quantum yield of PSII and enhances the amount of phenolic acids, flavonoids, and pigments in Lactuca sativa. Front. Plant Sci. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for nondestructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Muleo, R.; Morini, S. Light quality regulates shoot cluster growth and development of MM106 apple genotype in in vitro culture. Sci. Hortic. 2006, 108, 364–370. [Google Scholar] [CrossRef]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol. 2008, 50, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol. 2008, 26, 631–638. [Google Scholar] [CrossRef]
- Chory, J. Light signal transduction: An infinite spectrum of possibilities. Plant J. 2010, 61, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Bohne, F.; Linden, H. Regulation of carotenoid biosynthesis genes in response to light in Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2002, 1579, 26–34. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z. Primary, secondary metabolites, H2O2, malondialdehyde and photosynthetic responses of Orthosiphon stimaneus Benth. To different irradiance levels. Molecules 2012, 17, 1159–1176. [Google Scholar] [CrossRef] [Green Version]
- Werbrouck, S.; Buyle, H.; Geelen, D.; Van Labeke, M.C. Effect or red-, far-red- and blue-light emitting diodes on in vitro growth of Ficus benjamina. Acta Hortic. 2012, 961, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Bello-Bello, J.J.; Martinez-Estrada, E.; Caamal-Velazquez, J.H.; Morales-Ramos, V. Effect of LED light quality on in vitro shoot proliferation and growth of vanilla (Vanilla planifolia Andrews). Afr. J. Biotechnol. 2016, 15, 272–277. [Google Scholar]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef]
- Saleem, M.H.; Rehman, M.; Zahid, M.; Imran, M.; Xiang, W.; Liu, L. Morphological changes and antioxidative capacity of jute (Corchorus capsularis, Malvaceae) under different color light-emitting diodes. Braz. J. Bot. 2019, 42, 581–590. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Light | Spectrum Composition (%) | ||||
|---|---|---|---|---|---|
| 400–500 nm | 500–600 nm | 600–700 nm | 700–800 nm | R:FR Ratio | |
| Fluorescent Lamp (C) | 34.8 | 24.1 | 36.7 | 4.4 | 5.7 |
| AP67 | 12 | 16 | 57 | 16 | 3.0 |
| G2 | 8 | 2 | 65 | 25 | 3.1 |
| NS1 | 20 | 39 | 35 | 5 | 10.4 |
| Variety | Light Quality | Number of Neo-Formed Shoots | Node Number | Length of the Main Shoots (mm) |
|---|---|---|---|---|
| William | Control | 2.25 ± 0.48 ab | 9.92 ± 1.7 b | 2.43 ± 0.36 b |
| AP67 | 2.92 ± 0.39 a | 13.50 ± 1.8 a | 2.94 ± 0.25 ab | |
| NS1 | 2.15 ± 0.23 b | 12.51 ± 1.4 a | 2.96 ± 0.16 ab | |
| G2 | 2.08 ± 0.50 b | 9.67 ± 1.5 b | 3.09 ± 0.46 a | |
| William (avg) | 2.35 ± 0.6 A | 11.31 ± 1.7 A | 2.85 ± 0.25 A | |
| San Giovanni | Control | 1.82 ± 0.13 a | 11.25 ± 3.2 a | 2.54 ± 0.49 a |
| AP67 | 1.92 ± 0.39 a | 10.51 ± 1.8 ab | 2.15 ± 0.33 ab | |
| NS1 | 1.09 ± 0.20 b | 4.75 ± 2.4 c | 0.86 ± 0.36 c | |
| G2 | 1.68 ± 0.29 a | 8.25 ± 3.5 b | 1.95 ± 0.71 b | |
| San Giovanni (avg) | 1.63 ± 0.5 B | 8.94 ± 3.0 B | 1.88 ± 0.26 B | |
| Variety | Light Quality | Total Chlorophyll | Chlorophyll a | Chlorophyll b | Chl a/Chl b | Carotenoid | MDA (nmol g−1 f.w.) |
|---|---|---|---|---|---|---|---|
| William | Control | 1.91 ± 0.34 a | 1.23 ± 0.23 a | 0.68 ± 0.11 a | 1.81 ± 0.06 b | 0.33 ± 0.03 a | 3.72 ± 0.22 ab |
| AP67 | 1.05 ± 0.16 b | 0.82 ± 0.11ab | 0.23 ± 0.08 b | 3.53 ± 0.11 a | 0.25 ± 0.02 b | 3.98 ± 0.17 a | |
| NS1 | 1.04 ± 0.21 b | 0.70 ± 0.11 b | 0.34 ± 0.09 b | 2.19 ± 0.08 b | 0.16 ± 0.04 c | 3.11 ± 0.16 c | |
| G2 | 0.89 ± 0.24 b | 0.57 ± 0.12 c | 0.32 ± 0.05 b | 1.93 ± 0.09 b | 0.16 ± 0.01 c | 3.52 ± 0.30 bc | |
| William (average) | 1.22 ± 0.46 A | 0.83 ± 0.29 A | 0.39 ± 0.19 A | 2.37 ± 0.09 A | 0.23 ± 0.08 A | 3.63± 0.29 A | |
| San Giovanni | Control | 1.17 ± 0.22 a | 0.71 ± 0.09 a | 0.46 ± 0.07 a | 1.80 ± 0.04 b | 0.16 ± 0.04 a | 4.88 ± 0.34 a |
| AP67 | 0.60 ± 0.18 b | 0.47 ± 0.08 b | 0.13 ± 0.07 b | 3.61 ± 0.12 a | 0.15 ± 0.02 a | 4.65 ± 0.18 a | |
| NS1 | 0.89 ± 0.17 ab | 0.47 ± 0.10 b | 0.36 ± 0.09 ab | 1.41 ± 0.11 b | 0.14 ± 0.03 a | 2.78 ± 0.24 c | |
| G2 | 0.84 ± 0.21 ab | 0.56 ± 0.11 ab | 0.32 ± 0.08 ab | 1.77 ± 0.14 b | 0.16 ± 0.03 a | 3.17 ± 0.27 b | |
| San Giovanni (average) | 0.88 ± 0.24 B | 0.55 ± 0.11 B | 0.32 ± 0.11 A | 2.15 ± 0.18 A | 0.15 ± 0.01 A | 3.81 ± 0.35 A | |
| Kaiser-Meyer Olkin Measure of Sampling Adequacy | 0.6 | |
|---|---|---|
| Bartlett’s Test of Sphericity | Approx. Chi-Square | 5099.85 |
| df | 36 | |
| Sign. | 0.05 | |
| Variables | Chlorophyll a | Chlorophyll b | Carotenoid | Protein | MDA | Number of Neo-Formed Shoots | Node Number | Length of the Main Shoots |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll a | 1 | |||||||
| Chlorophyll b | 0.8 | 1 | ||||||
| Carotenoid | 0.9 | 0.6 | 1 | |||||
| Protein | 0.7 | 0.6 | 0.8 | 1 | ||||
| MDA | 0.1 | −0.1 | 0.1 | 0.0 | 1 | |||
| number of neo-formed shoots | 0.6 | −0.1 | 0.6 | 0.2 | 0.3 | 1 | ||
| node number | 0.4 | −0.1 | 0.3 | −0.1 | 0.5 | 0.9 | 1 | |
| length of the main shoots | 0.4 | 0.0 | 0.3 | −0.1 | 0.4 | 0.8 | 0.9 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupo, M.; Bashir, M.A.; Silvestri, C.; Brunori, E.; Pica, A.L.; Cristofori, V. LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro. Agronomy 2022, 12, 2531. https://doi.org/10.3390/agronomy12102531
Lupo M, Bashir MA, Silvestri C, Brunori E, Pica AL, Cristofori V. LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro. Agronomy. 2022; 12(10):2531. https://doi.org/10.3390/agronomy12102531
Chicago/Turabian StyleLupo, Michela, Muhammad Ajmal Bashir, Cristian Silvestri, Elena Brunori, Aniello Luca Pica, and Valerio Cristofori. 2022. "LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro" Agronomy 12, no. 10: 2531. https://doi.org/10.3390/agronomy12102531
APA StyleLupo, M., Bashir, M. A., Silvestri, C., Brunori, E., Pica, A. L., & Cristofori, V. (2022). LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro. Agronomy, 12(10), 2531. https://doi.org/10.3390/agronomy12102531