


Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens
 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms Used in the Research
2.1.1. Yeasts from the Metschnikowia Clade
2.1.2. Potato Phytopathogens
2.2. Agar-Well Diffusion Method for In Vitro Antimicrobial Activity Determination
2.3. Assessment of Pulcherrimin Production Profile by Selected Strains of Yeast Metschnikowia
2.4. Determination of the Enzymatic Activity of Selected Strains of Yeast Metschnikowia by API ZYM Test
2.5. Fermentation Byproducts of Selected Metschnikowia Yeast Strains by High-Performance Liquid Chromatography (HPLC)
2.6. Optimisation of the Acid-Whey-Based Culture Media for Cultivation of Metschnikowia Pulcherrima TK1
2.7. Acid-Whey-Based Media for Yeast Cultivation
2.8. Characteristics of the Chemical Composition of Selected Acid-Whey-Based Media
2.9. In Situ Assessment of Antimicrobial Activity on Seed Potatoes
2.10. Statistical Analysis
3. Results and Discussion
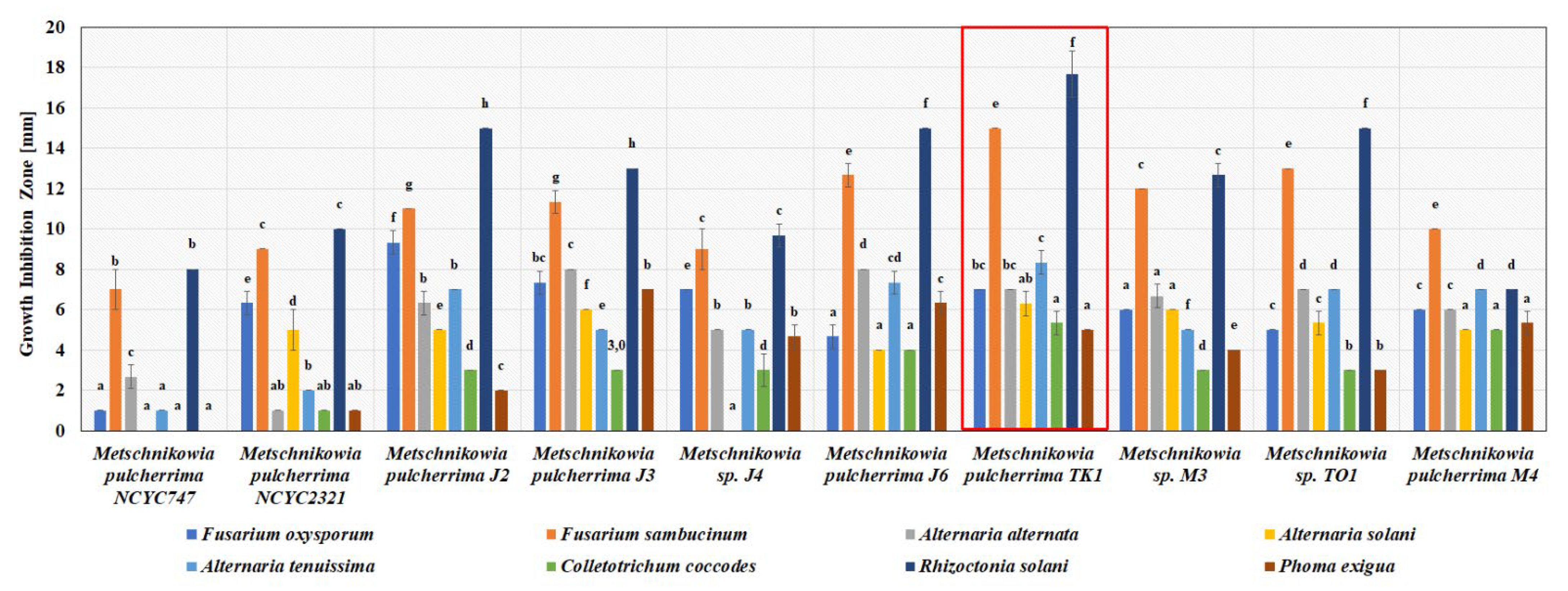
3.1. Antimicrobial Activity of Metschnikowia Yeasts In Vitro
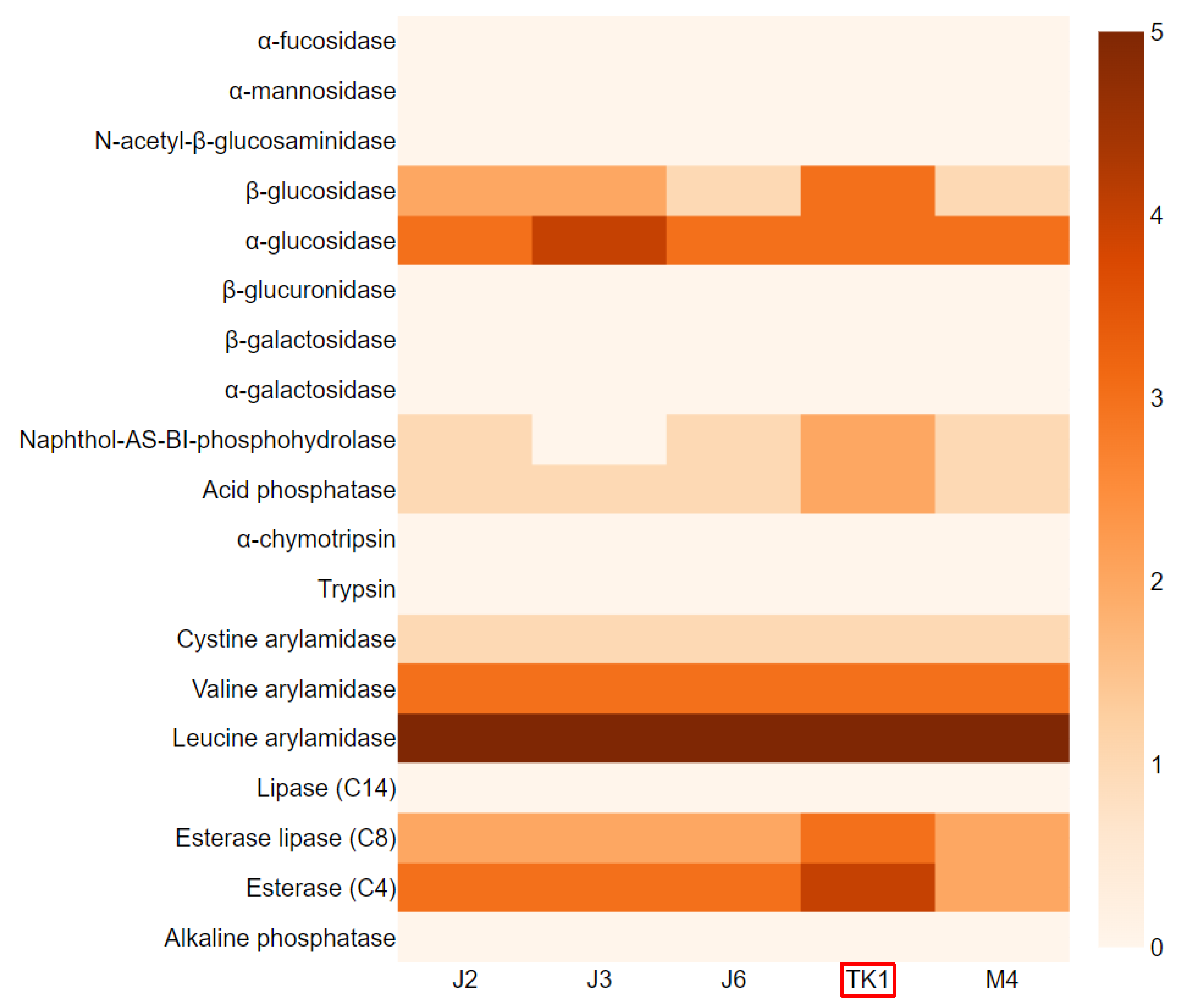
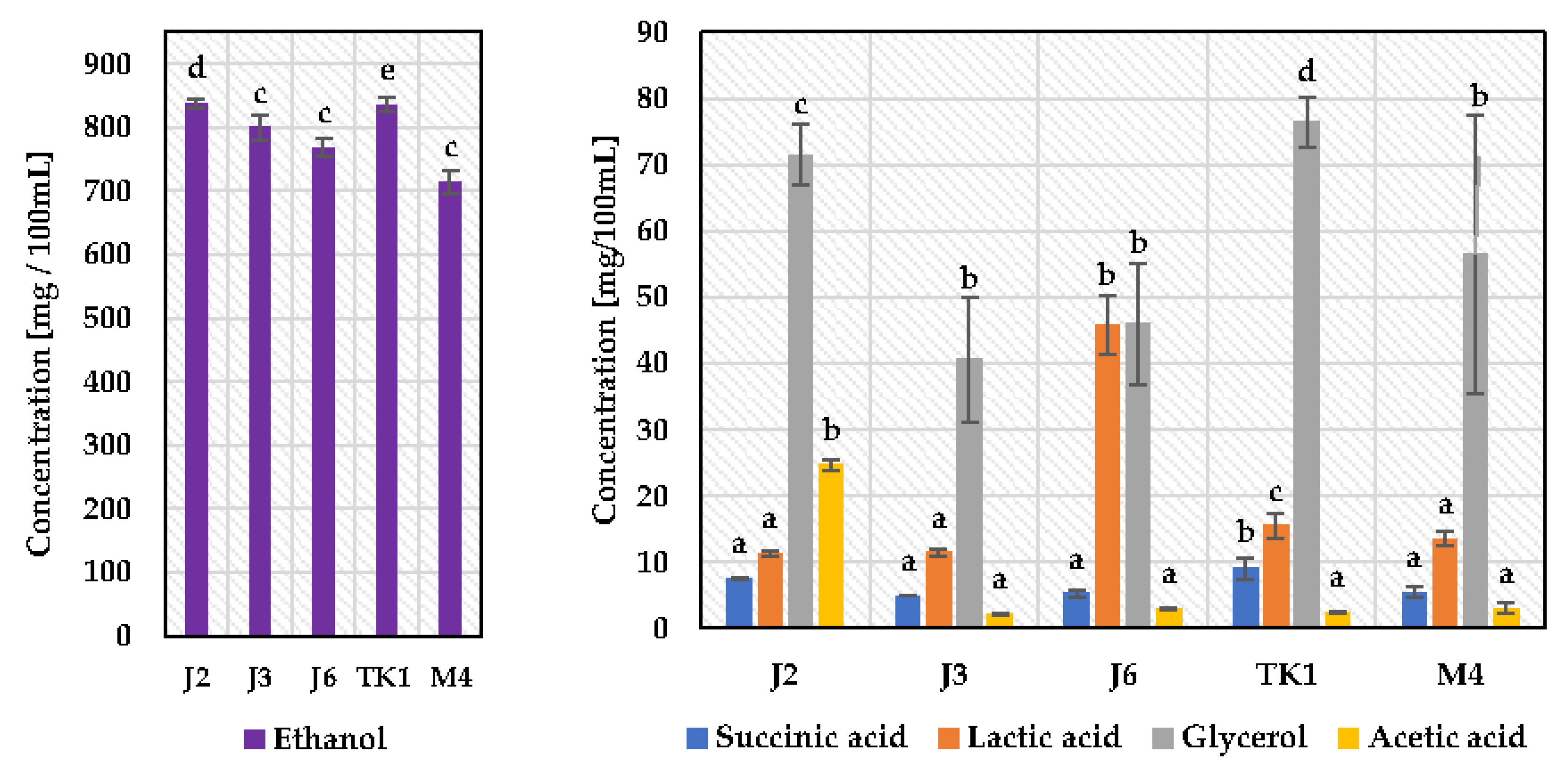
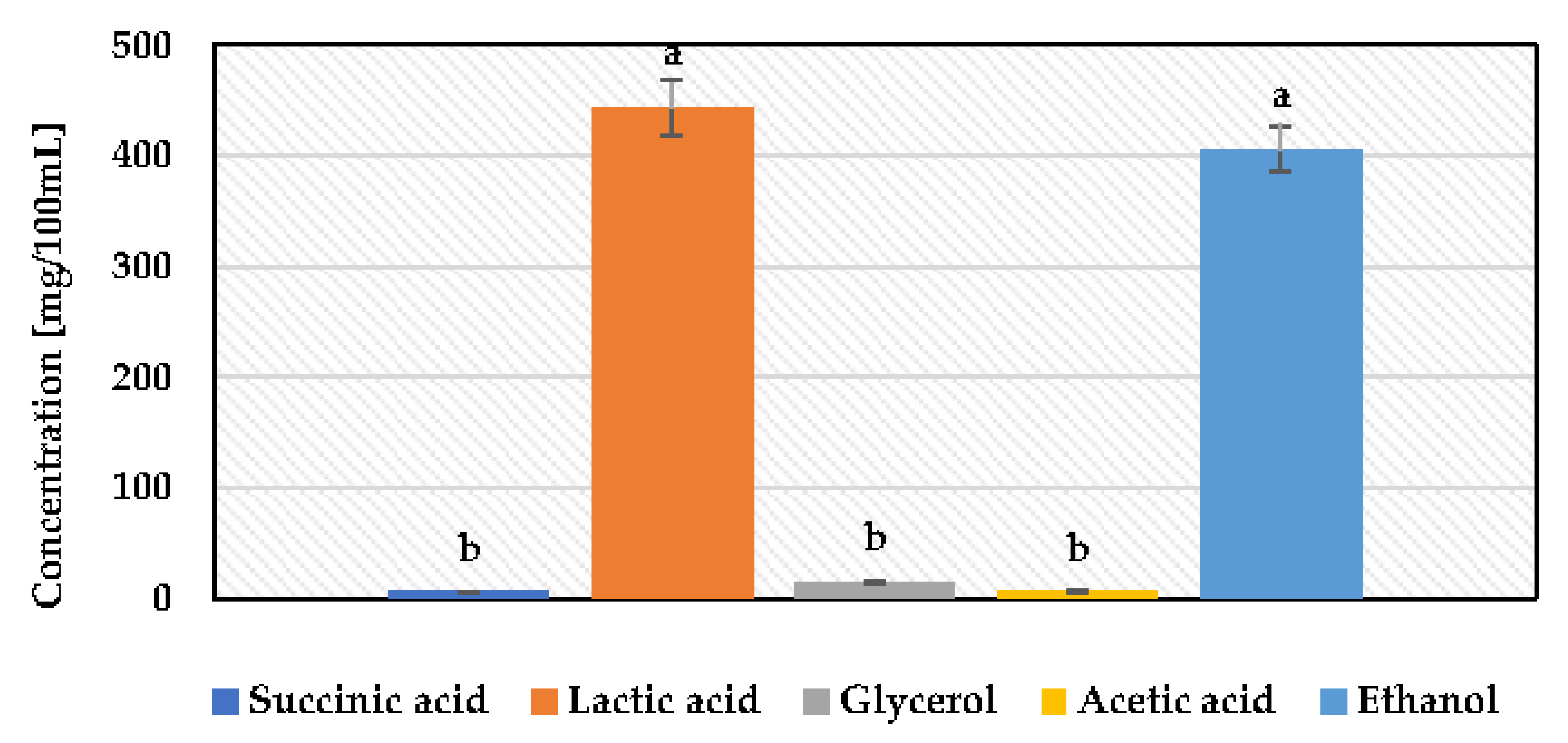
3.2. Determination of Pulcherrimin Formation, Enzymatic Activity, and Fermentation Byproducts of Metschnikowia Pulcherrima
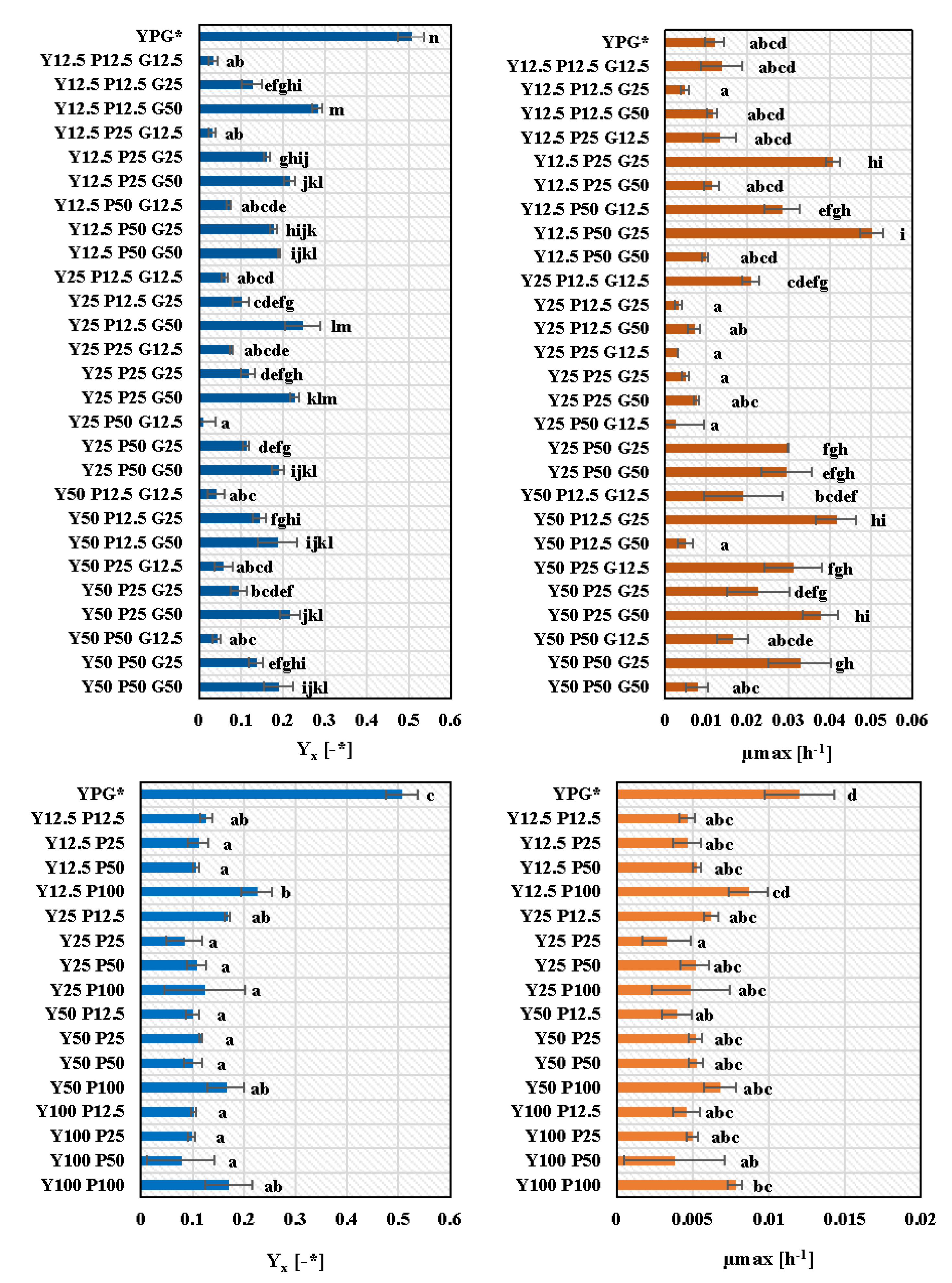
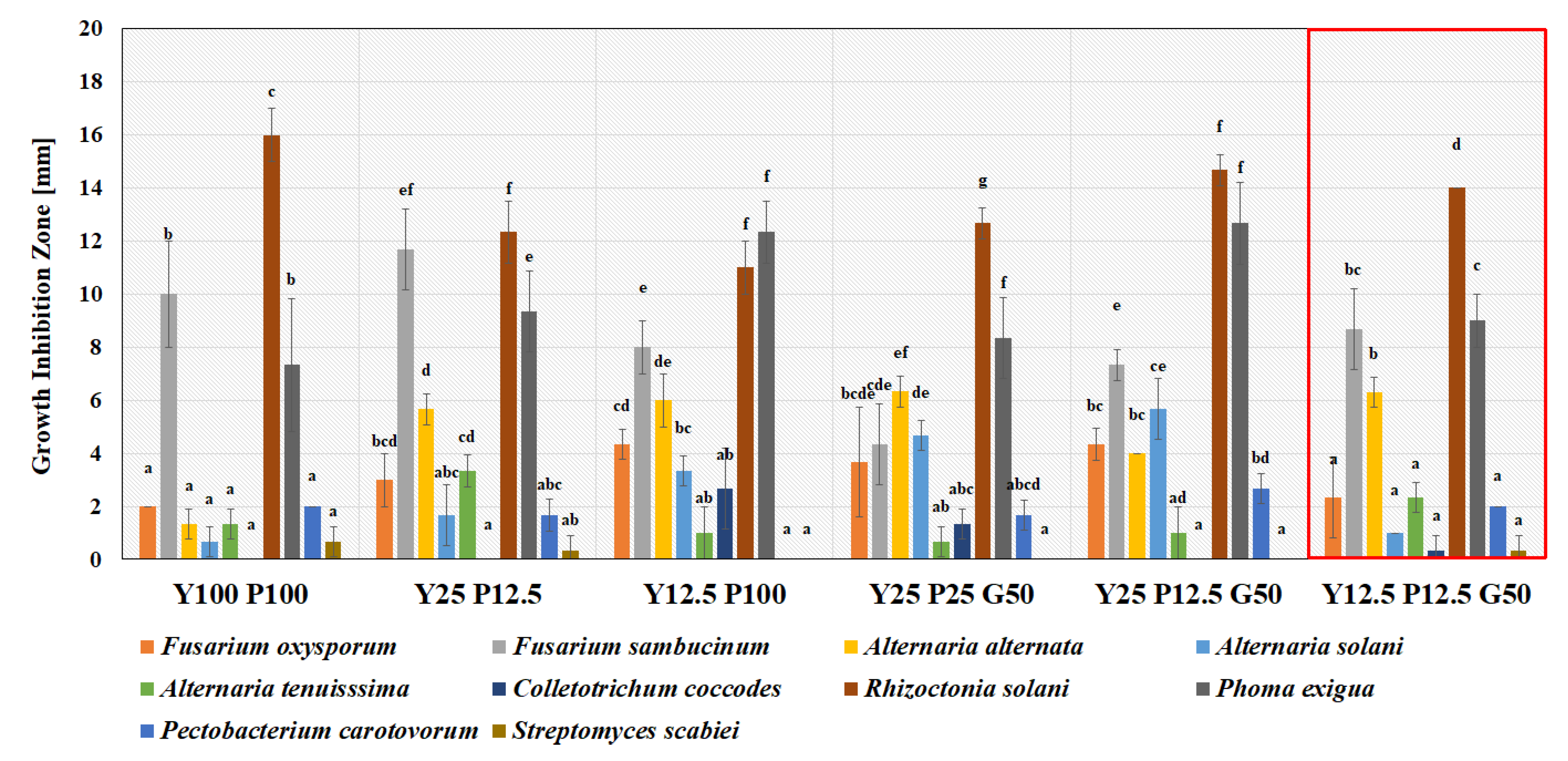
3.3. Growth Medium Selection for M. pulcherrima TK1 Cultivation
3.4. In Situ Test of Biocontrol Potential of M. pulcherrima TK1 against Phytopathogens on Potatoes
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, H.P. Policies and Strategies Conducive to Potato Development in Asia and the Pacific Region; RAP Publication (FAO): Rome, Italy, 2008; pp. 18–29. [Google Scholar]
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and Human Health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef] [PubMed]
- FAO. Top Countries in Potatoes Production. Potatoes Production—Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2019. [Google Scholar]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. Causing Potato Dry Rot in China and Susceptibility Evaluation of Chinese Potato Germplasm to the Pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Adolf, B.; Andrade-Piedra, J.; Bittara Molina, F.; Przetakiewicz, J.; Hausladen, H.; Kromann, P.; Lees, A.; Lindqvist-Kreuze, H.; Perez, W.; Secor, G.A. Fungal, Oomycete, and Plasmodiophorid Diseases of Potato. In The Potato Crop; Springer International Publishing: Cham, Switzerland, 2020; pp. 307–350. [Google Scholar]
- Tomilova, O.G.; Shaldyaeva, E.M.; Kryukova, N.A.; Pilipova, Y.V.; Schmidt, N.S.; Danilov, V.P.; Kryukov, V.Y.; Glupov, V.V. Entomopathogenic fungi decrease Rhizoctonia disease in potato in field conditions. PeerJ 2020, 8, e9895. [Google Scholar] [CrossRef] [PubMed]
- Sukul, P.; Spiteller, M. Metalaxyl: Persistence, degradation, metabolism, and analytical methods. Rev. Environ. Contam. Toxicol. 2000, 164, 1–26. [Google Scholar]
- Huang, Z.; Wang, P.; Pu, Z.; Lu, L.; Chen, G.; Hu, X.; Fayyaz, A.; Gai, Y. Effects of mancozeb on citrus rhizosphere bacterial community. Microb. Pathog. 2021, 154, 104845. [Google Scholar] [CrossRef] [PubMed]
- OuYang, M.-N.; Liu, X.; Guo, H.-M.; Lu, Z.-H.; Zhou, D.-D.; Yang, Z.-H. The different toxic effects of metalaxyl and metalaxyl-M on Tubifex tubifex. Ecotoxicol. Environ. Saf. 2021, 208, 111587. [Google Scholar] [CrossRef] [PubMed]
- Saquib, Q.; Siddiqui, M.A.; Ansari, S.M.; Alwathnani, H.A.; Musarrat, J.; Al-Khedhairy, A.A. Cytotoxicity and genotoxicity of methomyl, carbaryl, metalaxyl, and pendimethalin in human umbilical vein endothelial cells. J. Appl. Toxicol. 2021, 41, 832–846. [Google Scholar] [CrossRef]
- Eurostat. The EU Potato Sector—Statistics on Production, Prices and Trade. 2021. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=The_EU_potato_sector_-_statistics_on_production,_prices_and_trade (accessed on 28 June 2022).
- Eurostat, 2013: Glossary: Organic Farming. Statistics Explained. Available online: https://ec.europa.eu/eurostat/statisticsexplained/index.php?title=Glossary:Organic_farming&oldid=122448 (accessed on 28 June 2022).
- Thurman, J.H.; Crowder, D.W.; Northfield, T.D. Biological control agents in the Anthropocene: Current risks and future options. Curr. Opin. Insect Sci. 2017, 23, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Mendonca-Hagler, L.C.; Hagler, A.N.; Kurtzman, C.P. Phylogeny of Metschnikowia Species Estimated from Partial rRNA Sequences. Int. J. Syst. Bacteriol. 1993, 43, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef] [PubMed]
- Banani, H.; Spadaro, D.; Zhang, D.; Matic, S.; Garibaldi, A.; Gullino, M.L. Postharvest application of a novel chitinase cloned from Metschnikowia fructicola and overexpressed in Pichia pastoris to control brown rot of peaches. Int. J. Food Microbiol. 2015, 199, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol capability of local Metschnikowia sp. isolates. Antonie van Leeuwenhoek 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipiczki, M. Taxonomic Revision of the pulcherrima Clade of Metschnikowia (Fungi): Merger of Species. Taxonomy 2022, 2, 107–123. [Google Scholar] [CrossRef]
- Leverentz, B.; Conway, W.S.; Janisiewicz, W.; Abadias, M.; Kurtzman, C.P.; Camp, M.J. Biocontrol of the food-borne pathogens Listeria monocytogenes and Salmonella enterica serovar poona on fresh-cut apples with naturally occurring bacterial and yeast antagonists. Appl. Environ. Microbiol. 2006, 72, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Türkel, S.; Korukluoğlu, M.; Yavuz, M. Biocontrol Activity of the Local Strain of Metschnikowia pulcherrima on Different Postharvest Pathogens. Biotechnol. Res. Int. 2014, 2014, 397167. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Czyżowska, A.; Kucharska, A.Z.; Nowak, A.; Sokół-Łętowska, A.; Motyl, I.; Piórecki, N. Suitability of the probiotic lactic acid bacteria strains as the starter cultures in unripe cornelian cherry (Cornus mas L.) fermentation. J. Food Sci. Technol. 2017, 54, 2936–2946. [Google Scholar] [CrossRef]
- EN ISO 20483:2013; Cereals and Pulses—Determination of the Nitrogen Content and Calculation of the Crude Protein Content—Kjeldahl Method. ISO: Geneva, Switzerland, 2013.
- Standard methods for the examination of water and wastewater. In APHA; Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. (Eds.) American Public Health Association (APHA); American Water Works Association (AWWA); Water Environment Federation (WEF): Washington, DC, USA, 2012. [Google Scholar]
- Steglińska, A.; Bekhter, A.; Wawrzyniak, P.; Kunicka-Styczyńska, A.; Jastrząbek, K.; Fidler, M.; Śmigielski, K.; Gutarowska, B. Antimicrobial Activities of Plant Extracts against Solanum tuberosum L. Phytopathogens. Molecules 2022, 27, 1579. [Google Scholar] [CrossRef] [PubMed]
- Steglińska, A.; Kołtuniak, A.; Motyl, I.; Berłowska, J.; Czyżowska, A.; Cieciura-Włoch, W.; Okrasa, M.; Kręgiel, D.; Gutarowska, B. Lactic Acid Bacteria as Biocontrol Agents against Potato (Solanum tuberosum L.). Pathogens. Appl. Sci. 2022, 12, 7763. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol potential of Metchnikowia pulcherrima strains against blue mold of apple. Phytopathology 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadaro, D.; Ciavorella, A.; Dianpeng, Z.; Garibaldi, A.; Gullino, M. Effect of culture media and pH on the biomass production and biocontrol efficacy of a Metschnikowia pulcherrima strain to be used as a biofungicide for postharvest disease control. Can. J. Microbiol. 2010, 56, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Lorè, A.; Garibaldi, A.; Gullino, M.L. A new strain of Metschnikowia fructicola for postharvest control of Penicillium expansum and patulin accumulation on four cultivars of apple. Postharvest Biol. Technol. 2013, 75, 1–8. [Google Scholar] [CrossRef]
- Settier-Ramírez, L.; López-Carballo, G.; Hernández-Muñoz, P.; Fontana, A.; Strub, C.; Schorr-Galindo, S. New isolated metschnikowia pulcherrima strains from apples for postharvest biocontrol of penicillium expansum and patulin accumulation. Toxins 2021, 13, 397. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Smilanick, J.L.; Gabler, F.M.; Mansour, M.; Droby, S. Near-Harvest Applications of Metschnikowia fructicola, Ethanol, and Sodium Bicarbonate to Control Postharvest Diseases of Grape in Central California. Plant Dis. 2003, 87, 1384–1389. [Google Scholar] [CrossRef] [Green Version]
- Agarbati, A.; Canonico, L.; Pecci, T.; Romanazzi, G.; Ciani, M.; Comitini, F. Biocontrol of Non-Saccharomyces Yeasts in Vineyard against the Gray Mold Disease Agent Botrytis cinerea. Microorganisms 2022, 10, 200. [Google Scholar] [CrossRef]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Guo, D.; Zhu, L.; Hou, X. Combination of UV-C Treatment and Metschnikowia pulcherrimas for Controlling Alternaria Rot in Postharvest Winter Jujube Fruit. J. Food Sci. 2015, 80, M137–M141. [Google Scholar] [CrossRef] [PubMed]
- Kregiel, D.; Nowacka, M.; Rygala, A.; Vadkertiová, R. Biological Activity of Pulcherrimin from the Meschnikowia pulcherrima Clade. Molecules 2022, 27, 1855. [Google Scholar] [CrossRef] [PubMed]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Schläfli, M.; Kirner, P.; Santos Kron, A.; Rueda-Mejia, M.P.; et al. Snf2 controls pulcherriminic acid biosynthesis and antifungal activity of the biocontrol yeast Metschnikowia pulcherrima. Mol. Microbiol. 2019, 112, 317–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadaro, D.; Vola, R.; Piano, S.; Gullino, M.L. Mechanisms of action and efficacy of four isolates of the yeast Metschnikowia pulcherrima active against postharvest pathogens on apples. Postharvest Biol. Technol. 2002, 24, 123–134. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Kregiel, D.; Pawlikowska, E. Kwas pulcherriminowy i jego znaczenie w inhibicji mikroflory zanieczyszczającej żywność oraz środowiska fermentacyjne. In Wybrane Substancje o Znaczeniu Biologicznym: Spojrzenie Młodych Naukowców, 1st ed.; Kwolek-Mirek, M., Mołoń, M., Eds.; Creativetime: Kraków, Poland, 2017; pp. 143–149. [Google Scholar]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef]
- Pawlikowska, E.; Kregiel, D. Enzymatic profiles and antimicrobial activity of the yeast Metschnikowia. Acta Innov. 2017, 23, 17–24. [Google Scholar]
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [Green Version]
- Le Lay, C.; Coton, E.; Le Blay, G.; Chobert, J.-M.; Haertlé, T.; Choiset, Y.; Van Long, N.N.; Meslet-Cladière, L.; Mounier, J. Identification and quantification of antifungal compounds produced by lactic acid bacteria and propionibacteria. Int. J. Food Microbiol. 2016, 239, 79–85. [Google Scholar] [CrossRef]
- Coban, H.B. Organic acids as antimicrobial food agents: Applications and microbial productions. Bioprocess Biosyst. Eng. 2020, 43, 569–591. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Biocontrol of postharvest brown rot of sweet cherries by Saccharomyces cerevisiae Disva 599, Metschnikowia pulcherrima Disva 267 and Wickerhamomyces anomalus Disva 2 strains. Postharvest Biol. Technol. 2014, 96, 64–68. [Google Scholar] [CrossRef]
- Tian, Y.-q.; Li, W.; Jiang, Z.-t.; Jing, M.-m.; Shao, Y.-Z. The preservation effect of Metschnikowia pulcherrima yeast on anthracnose of postharvest mango fruits and the possible mechanism. Food Sci. Biotechnol. 2018, 27, 95–105. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin |
|---|---|
| Metschnikowia pulcherrima NCYC 747 | National Collection of Yeast Cultures (Norwich, UK) |
| Metschnikowia pulcherrima NCYC 2321 | |
| Metschnikowia pulcherrima J2* | Isolate from apple fruit |
| Metschnikowia pulcherrima J3* | Isolate from apple fruit |
| Metschnikowia sp. J4 | Isolate from apple fruit |
| Metschnikowia pulcherrima J6* | Isolate from apple fruit |
| Metschnikowia pulcherrima TK1* | Isolate from strawberry flower |
| Metschnikowia sp. M3 | Isolate from raspberry fruit |
| Metschnikowia sp. TO1 | Isolate from strawberry fruit |
| Metschnikowia pulcherrima M4* | Isolate from raspberry fruit |
| Medium Variants | Yeast Extract (%) | Peptone (%) | Glucose (%) | Medium Variants | Yeast Extract (%) | Peptone (%) |
|---|---|---|---|---|---|---|
| Y50 P50 G50 | 0.5 | 1.0 | 1.0 | Y100 P100 | 1.0 | 2.0 |
| Y50 P50 G25 | 0.5 | 1.0 | 0.5 | Y100 P50 | 1.0 | 1.0 |
| Y50 P50 G12.5 | 0.5 | 1.0 | 0.25 | Y100 P25 | 1.0 | 0.5 |
| Y50 P25 G50 | 0.5 | 0.5 | 1.0 | Y100 P12.5 | 1.0 | 0.25 |
| Y50 P25 G25 | 0.5 | 0.5 | 0.5 | Y50 P100 | 0.5 | 2.0 |
| Y50 P25 G12.5 | 0.5 | 0.5 | 0.25 | Y50 P50 | 0.5 | 1.0 |
| Y50 P12.5 G50 | 0.5 | 0.25 | 1.0 | Y50 P25 | 0.5 | 0.5 |
| Y50 P12.5 G25 | 0.5 | 0.25 | 0.5 | Y50 P12.5 | 0.5 | 0.25 |
| Y50 P12.5 G12.5 | 0.5 | 0.25 | 0.25 | Y25 P100 | 0.25 | 2.0 |
| Y25 P50 G50 | 0.25 | 1.0 | 1.0 | Y25 P50 | 0.25 | 1.0 |
| Y25 P50 G25 | 0.25 | 1.0 | 0.5 | Y25 P25 | 0.25 | 0.5 |
| Y25 P50 G12.5 | 0.25 | 1.0 | 0.25 | Y25 P12.5 | 0.25 | 0.25 |
| Y25 P25 G50 | 0.25 | 0.5 | 1.0 | Y12.5 P100 | 0.13 | 2.0 |
| Y25 P25 G25 | 0.25 | 0.5 | 0.5 | Y12.5 P50 | 0.13 | 1.0 |
| Y25 P25 G12.5 | 0.25 | 0.5 | 0.25 | Y12.5 P25 | 0.13 | 0.5 |
| Y25 P12.5 G50 | 0.25 | 0.25 | 1.0 | Y12.5 P12.5 | 0.13 | 0.25 |
| Y25 P12.5 G25 | 0.25 | 0.25 | 0.5 | |||
| Y25 P12.5 G12.5 | 0.25 | 0.25 | 0.25 | |||
| Y12.5 P50 G50 | 0.13 | 1.0 | 1.0 | |||
| Y12.5 P50 G25 | 0.13 | 1.0 | 0.5 | |||
| Y12.5 P50 G12.5 | 0.13 | 1.0 | 0.25 | |||
| Y12.5 P25 G50 | 0.13 | 0.5 | 1.0 | |||
| Y12.5 P25 G25 | 0.13 | 0.5 | 0.5 | |||
| Y12.5 P25 G12.5 | 0.13 | 0.5 | 0.25 | |||
| Y12.5 P12.5 G50 | 0.13 | 0.25 | 1.0 | |||
| Y12.5 P12.5 G25 | 0.13 | 0.25 | 0.5 | |||
| Y12.5 P12.5 G12.5 | 0.13 | 0.25 | 0.25 | |||
| YPG | 1 | 2 | 2 |
| FeCl3 Supplementation (mg/mL) | Growth Features and Pulcherrimin Formation | Strain | ||||
|---|---|---|---|---|---|---|
| J2 | J3 | J6 | TK1 | M4 | ||
| Without (control) | Colony colour top/bottom | white/creamy | white/creamy | white/creamy | white/creamy | white/creamy |
| Coloured halo (mm) | 0.5 | 0.5 | 0.5 | 4.0 | 0.0 | |
| 0.005 | Colony colour top/bottom | white/creamy | white/creamy | white/creamy | white/creamy | white/creamy |
| Coloured halo (mm) | 1.0 | 1.0 | 2.0 | 6.0 | 1.0 | |
| 0.01 | Colony colour top/bottom | white/red | white/red | white/red | white/red | white/red |
| Coloured halo (mm) | 0.5 | 0.0 | 0.0 | 1.5 | 0.0 | |
| 0.2 | Colony colour top/bottom | red/red | red/red | red/red | red/dark red | red/red |
| Coloured halo (mm) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Analytical Analysis | Medium | ||
|---|---|---|---|
| Acid Whey Broth Supplemented Y12.5 P12.5 G50 | Acid Whey Broth without Supplementation | YPG Broth | |
| Phosphorus (mg/L) | 175.56 | 181.90 | 182.08 |
| Nitrogen (mg/L) | 75.76 | 38.20 | 62.59 |
| Iron (mg/L) | 28.00 | 0.50 | 5.00 |
| Potassium (mg/L) | 104.5 | 75.00 | 65.70 |
| Calcium (mg/L) | 289.00 | 324.55 | 9.20 |
| Chloride (mg/L) | 32.80 | 57.10 | 0.18 |
| Sodium (mg/L) | 38.90 | 62.85 | 0.26 |
| Lactose (g/L) | 9.00 | 14.55 | 0.20 |
| Protein (g/kg) | 11.38 | 8.50 | 32.64 |
| Lactic acid (g/L) | 16.30 | 15.85 | 1.22 |
| Total solids (g/kg) | 78.71 | 58.00 | 58.18 |
| Volatile solids (g/kg) | 69.37 | 48.05 | 55.04 |
| Water content (g/kg) | 921.29 | 941.10 | 941.82 |
| Phytopathogens | Inhibition of Potato Infestation (%) | |
|---|---|---|
| Acid Whey Broth Supplemented Y12.5 P12.5 G50 | YPD Broth | |
| Fusarium oxysporum | 30 ± 10 | 30 ± 5 |
| Fusarium sambucinum | 100 ± 0 * | 80 ± 10 * |
| Alternaria alternata | 100 ± 0 * | 80 ± 5 * |
| Alternaria solani | 100 ± 0 * | 90 ± 5 * |
| Alternaria tenuissima | 100 ± 0 | 95 ± 5 |
| Colletotrichum coccodes | 100 ± 0 | 100 ± 0 |
| Rhizoctonia solani | 50 ± 5 | 60 ± 5 |
| Phoma exigua | 100 ± 0 * | 80 ± 5 * |
| Pectobacterium carotovorum | - | - |
| Streptomyces scabiei | 40 ± 10 * | 20 ± 5 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steglińska, A.; Kołtuniak, A.; Berłowska, J.; Czyżowska, A.; Szulc, J.; Cieciura-Włoch, W.; Okrasa, M.; Kręgiel, D.; Gutarowska, B. Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens. Agronomy 2022, 12, 2546. https://doi.org/10.3390/agronomy12102546
Steglińska A, Kołtuniak A, Berłowska J, Czyżowska A, Szulc J, Cieciura-Włoch W, Okrasa M, Kręgiel D, Gutarowska B. Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens. Agronomy. 2022; 12(10):2546. https://doi.org/10.3390/agronomy12102546
Chicago/Turabian StyleSteglińska, Aleksandra, Artur Kołtuniak, Joanna Berłowska, Agata Czyżowska, Justyna Szulc, Weronika Cieciura-Włoch, Małgorzata Okrasa, Dorota Kręgiel, and Beata Gutarowska. 2022. "Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens" Agronomy 12, no. 10: 2546. https://doi.org/10.3390/agronomy12102546
APA StyleSteglińska, A., Kołtuniak, A., Berłowska, J., Czyżowska, A., Szulc, J., Cieciura-Włoch, W., Okrasa, M., Kręgiel, D., & Gutarowska, B. (2022). Metschnikowia pulcherrima as a Biocontrol Agent against Potato (Solanum tuberosum) Pathogens. Agronomy, 12(10), 2546. https://doi.org/10.3390/agronomy12102546