
Weeds in Cereal Crop Rotations May Host Fusarium Species That Cause Fusarium Head Blight and Grain Weight Losses in Wheat

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolate Collection
2.2. Pathogenicity Test
2.3. Spring Wheat Head Inoculation under Field Condition
2.4. Determination of 1000 Grains Weight
2.5. Grain Homogenization
2.6. DNA Extraction, Fungal DNA Content Determination
2.7. Molecular Identification and RT-PCR
2.8. Fusarium spp. DNA Content Quantification
2.9. Statistical Analysis
3. Results
3.1. F. graminearum and F. avenaceum DNA Content Identification in Grains
3.2. Pathogenicity of Fusarium Strains from Alternative (Weed) and Main Host (Wheat) to Spring Wheat
3.3. Effect of Fusarium Strains Isolated from Alternative (Weeds) and Main (Wheat) Host Plants on Grain Weight
3.4. Correlation Determination between the DNA Content of Fusarium Strains, Fusarium Head Blight Intensity, and Spring Wheat 1000-Grain Weight
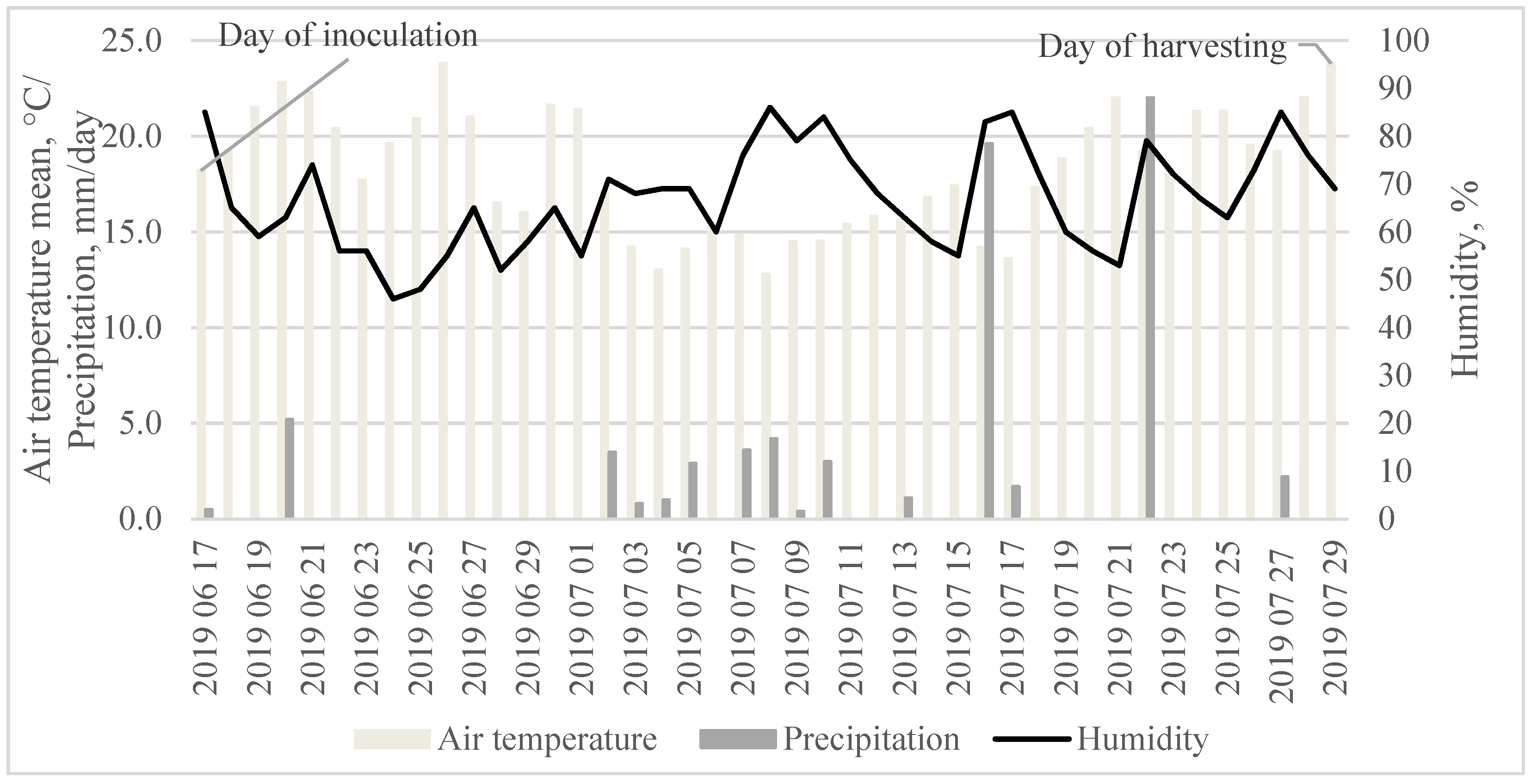
3.5. Weather Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaughan, M.; Backhouse, D.; Del Ponte, E.M. Climate change impacts on the ecology of Fusarium graminearum species complex and susceptibility of wheat to Fusarium head blight: A review. World Mycotoxin J. 2016, 9, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Lu, F.; Zhou, Z.; Zhao, H.; Zhang, J.; Mao, Y.; Li, M.; Wang, J.; Zhou, M. Quinone outside inhibitors affect DON biosynthesis, mitochondrial structure and toxisome formation in Fusarium graminearum. J. Hazard. Mater. 2020, 398, 122908. [Google Scholar] [CrossRef] [PubMed]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Cor, O.; Barbier, G.; Le Floch, G. Development of qPCR assays to monitor the ability of Gliocladium catenulatum J1446 to reduce the cereal pathogen Fusarium graminearum inoculum in soils. Eur. J. Plant Pathol. 2018, 152, 285–295. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekiunaite, L.; Petrović, D.M.; Westereng, B.; Vaaje-Kolstad, G.; Hachem, M.A.; Várnai, A.; Eijsink, V.G. FgLPMO 9A from Fusarium graminearum cleaves xyloglucan independently of the backbone substitution pattern. FEBS Lett. 2016, 590, 3346–3356. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Xu, J.; Zhang, X.; Wang, S.; Xing, Y.; Mokoena, M.P.; Olaniran, A.O.; Shi, J. Gramineous weeds near paddy fields are alternative hosts for the Fusarium graminearum species complex that causes fusarium head blight in rice. Plant Pathol. 2020, 69, 433–441. [Google Scholar] [CrossRef]
- Brown, N.A.; Bass, C.; Baldwin, T.K.; Chen, H.; Massot, F.; Carion, P.W.C.; Urban, M.; van de Meene, A.M.L.; Hammond-Kosack, K.E. Characterisation of the Fusarium graminearum-wheat floral interaction. J. Pathog. 2011, 2011, 626345. [Google Scholar] [CrossRef] [Green Version]
- Wegulo, S.N. Factors influencing deoxynivalenol accumulation in small grain cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Hofte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2013, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Janavičienė, S.; Mankevičienė, A.; Kochiieru, Y. The fate of deoxynivalenol and its derivatives in spring wheat whole-grain flour during storage. Zemdirb. Agric. 2020, 107, 123–130. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef] [PubMed]
- Becher, R.; Miedaner, T.; Wirsel, S.G. 8 Biology, diversity, and management of FHB-causing Fusarium species in small-grain cereals. In Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2012; pp. 199–241. [Google Scholar]
- Brown, N.A.; Urban, M.; van de Meene, A.M.L.; Hammond-Kosack, K.E. The infection biology of Fusarium graminearum: Defining the pathways of spikelet to spikelet colonisation in wheat ears. Fungal Biol. 2010, 114, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.T.; McCormick, S.P.; Vaughan, M.M. Effects of Atmospheric CO2 and Temperature on Wheat and Corn Susceptibility to Fusarium graminearum and Deoxynivalenol Contamination. Plants 2021, 10, 2582. [Google Scholar] [CrossRef] [PubMed]
- Postic, J.; Cosic, J.; Vrandecic, K.; Jurkovic, D.; Saleh, A.A.; Leslie, J.F. Diversity of Fusarium Species Isolated from Weeds and Plant Debris in Croatia. J. Phytopathol. 2012, 160, 76–81. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Mwangi, M.; Aigbe, S.O.; Leslie, J.F. Fusarium species from the cassava root rot complex in West Africa. Phytopathology 2006, 96, 673–676. [Google Scholar] [CrossRef] [Green Version]
- Suproniene, S.; Kadziene, G.; Irzykowski, W.; Sneideris, D.; Ivanauskas, A.; Sakalauskas, S.; Serbiak, P.; Svegzda, P.; Kelpsiene, J.; Pranaitiene, S.; et al. Asymptomatic weeds are frequently colonized by pathogenic species of Fusarium in cereal-based crop rotations. Weed Res. 2019, 59, 312–323. [Google Scholar] [CrossRef]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Mourelos, C.A.; Malbrán, I.; Balatti, P.A.; Ghiringhelli, P.D.; Lori, G.A. Gramineous and non-gramineous weed species as alternative hosts of Fusarium graminearum, causal agent of Fusarium head blight of wheat, in Argentina. Crop Prot. 2014, 65, 100–104. [Google Scholar] [CrossRef]
- Altinok, H.H. Fusarium species isolated from common weeds in eggplant fields and symptomless hosts of Fusarium oxysporum f. sp. melongenae in Turkey. J. Phytopathol. 2013, 161, 335–340. [Google Scholar] [CrossRef]
- Ilić, J.; Ćosić, J.; Jurković, D.; Vrandečić, K. Pathogenicity of Fusarium spp. isolated from weeds and plant debris in eastern Croatia to wheat and maize. Poljoprivreda 2012, 18, 7–11. Available online: https://hrcak.srce.hr/94622 (accessed on 3 September 2022).
- Fulcher, M.R.; Winans, J.B.; Quan, M.; Oladipo, E.D.; Bergstrom, G.C. Population genetics of Fusarium graminearum at the interface of wheat and wild grass communities in New York. Phytopathology 2019, 109, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Inch, S.; Gilbert, J. The incidence of Fusarium species recovered from inflorescences of wild grasses in southern Manitoba. Plant Pathol. 2013, 25, 379–383. [Google Scholar] [CrossRef]
- Turkington, T.K.; Clear, R.M.; Demeke, T.; Lange, R.; Xi, K.; Kumar, K. Isolation of Fusarium graminearum from cereal, grass and corn residues from Alberta, 2001–2003. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2011, 33, 179–186. [Google Scholar] [CrossRef]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Toth, B.; Varga, J.; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.H.; Schofbeck, D.; McCormick, S.; Broz, K.; Stuckler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef]
- Lofgren, L.A.; LeBlanc, N.R.; Certano, A.K.; Nachtigall, J.; LaBine, K.M.; Riddle, J.; Broz, K.; Dong, Y.; Bethan, B.; Kafer, C.W.; et al. Fusarium graminearum: Pathogen or endophyte of North American grasses? New Phytol. 2018, 217, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Cuperlovic-Culf, M.; Vaughan, M.M.; Vermillion, K.; Surendra, A.; Teresi, J.; McCormick, S.P. Effects of atmospheric CO2 level on the metabolic response of resistant and susceptible wheat to Fusarium graminearum infection. Mol. Plant-Microbe Interact. 2019, 32, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Sakalauskas, S.; Stumbriene, K.; Suproniene, S.; Svegzda, P. Changes in Fusarium Link species composition from Lithuanian wheat grain in years 2005–2007 to 2011–2013. Proc. Latv. Univ. Agric. 2014, 32, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Suproniene, S.; Sakalauskas, S.; Stumbriene, K.; Zvirdauskiene, R.; Svegzda, P. Variances in trichothecene chemotype distribution in Lithuanian wheat grain and within pure culture Fusarium graminearum isolated from the same grain samples. Eur. J. Plant Pathol. 2015, 144, 371–381. [Google Scholar] [CrossRef]
- Góral, T.; Wiśniewska, H.; Ochodzki, P.; Nielsen, L.K.; Walentyn-Góral, D.; Stępień, Ł. Relationship between Fusarium Head Blight, Kernel Damage, Concentration of Fusarium Biomass, and Fusarium Toxins in Grain of Winter Wheat Inoculated with Fusarium culmorum. Toxins 2019, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Góral, T.; Ochodzki, P.; Nielsen, L.K.; Walentyn-Góral, D. Fusarium species and Fusarium mycotoxins in grain of wheat in Poland in 2009 and 2010. Preprints 2019, 2019090108. [Google Scholar] [CrossRef] [Green Version]
- Suproniene, S.; Kadziene, G.; Irzykowski, W.; Sneideris, D.; Ivanauskas, A.; Sakalauskas, S.; Serbiak, P.; Svegzda, P.; Auskalniene, O.; Jedryczka, M. Weed species within cereal crop rotations can serve as alternative hosts for Fusarium graminearum causing Fusarium head blight of wheat. Fungal Ecol. 2019, 37, 30–37. [Google Scholar] [CrossRef]
- Engle, J.S.; Lipps, P.E.; Mills, D. Fusarium Head Blight Severity Scale for Winter Wheat; Bulletin AC-48-03, Extension Factsheet; Ohio State University: Columbus, OH, USA, 2003. [Google Scholar]
- Nicolaisen, M.; Supronienė, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Arata, A.F.; Fernández, M.D.; Stenglein, S.A.; Dinolfo, M.I. Fusarium species richness in mono-and dicotyledonous weeds and their ability to infect barley and wheat. Mycol. Prog. 2021, 20, 1203–1216. [Google Scholar] [CrossRef]
- Sieber, T.; Riesen, T.K.; Müller, E.; Fried, P.M. Endophytic fungi in four winter wheat cultivars (Triticum aestivum L.) differing in resistance against Stagonospora nodorum (Berk.) Cast. & Germ. = Septoria nodorum (Berk.) Berk. J. Phytopathol. 1988, 122, 289–306. [Google Scholar] [CrossRef]
- Jenkinson, P.; Parry, D.W. Isolation of Fusarium species from common broad-leaved weeds and their pathogenicity to winter wheat. Mycol. Res. 1994, 98, 776–780. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. In Epidemiology of Mycotoxin Producing Fungi; Springer: Dordrecht, The Netherlands, 2003; pp. 755–768. [Google Scholar] [CrossRef]
- Janaviciene, S.; Suproniene, S.; Kadziene, G.; Pavlenko, R.; Berzina, Z.; Bartkevics, V. Toxigenicity of F. graminearum residing on host plants alternative to wheat as influenced by environmental conditions. Toxins 2022, 14, 541. [Google Scholar] [CrossRef]
- Fulcher, M.R.; Oladipo, E.D.; Winans, J.B.; Quan, M.; Bergstrom, G.C. Structure and diversity of Fusarium communities inhabiting non-cultivated grass inflorescences in New York State. Can. J. Plant Pathol. 2021, 43, 48–55. [Google Scholar]
- Parikka, P.; Hakala, K.; Tiilikkala, K. Expected shifts in Fusarium species’ composition on cereal grain in Northern Europe due to climatic change. Food Addit. Contam. 2012, 29 Pt A, 1543–1555. [Google Scholar] [CrossRef]
- Backhouse, D. Global distribution of Fusarium graminearum, F. asiaticum and F. boothii from wheat in relation to climate. Eur. J. Plant Pathol. 2014, 139, 161–173. [Google Scholar] [CrossRef]
- Schöneberg, T.; Musa, T.; Forrer, H.R.; Mascher, F.; Bucheli, T.D.; Bertossa, M.; Keller, B.; Vogelgsang, S. Infection conditions of Fusarium graminearum in barley are variety specific and different from those in wheat. Eur. J. Plant Pathol. 2018, 151, 975–989. [Google Scholar] [CrossRef]
- Twamley, T.; Gaffney, M.; Feechan, A.A. Microbial Fermentation Mixture Reduces Fusarium Head Blight and Promotes Grain Weight but does not impact Septoria tritici blotch. Eur. J. Plant Pathol. 2022, 162, 203–219. [Google Scholar] [CrossRef]
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and deoxynivalenol during the 2012/13 wheat Fusarium head blight outbreak in Argentina. Cereal Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Xue, A.; Ho, K.; Butler, G.; Vigier, B.; Babcock, C. Pathogenicity of Fusarium species causing head blight in barley. Phytoprotection 2006, 87, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Vogelgsang, S.; Sulyok, M.; Hecker, A.; Jenny, E.; Krska, R.; Schuhmacher, R.; Forrer, H.R. Toxigenicity and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. Eur. J. Plant Pathol. 2008, 122, 265–276. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Bartók, T.; Kászonyi, G.; Varga, M.; Tóth, B.; Varga, J. Common resistance to different Fusarium spp. causing Fusarium head blight in wheat. Eur. J. Plant Pathol. 2005, 112, 267–281. [Google Scholar] [CrossRef]
- Abdellatif, L.; Fernandez, M.R.; Bouzid, S.; Vujanovic, V. Characterization of virulence and PCR-DGGE profiles of Fusarium avenaceum from western Canadian Prairie Ecozone of Saskatchewan. Can. J. Plant Pathol. 2010, 32, 468–480. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment No. | Host Plant | F. avenaceum Strain Code | F. graminearum Strain Code |
|---|---|---|---|
| 1 | Spring wheat (Triticum aestivum) | SW-G1 | SW-6K5V1 |
| 2 | SW-TG5 | SW-6K4V1 | |
| 3 | Wild buckwheat (Fallopia convolvulus) | WB-1180l | WB-544r |
| 4 | WB-1178fl | WB-144r | |
| 5 | Meadow grass (Poa annua) | MG-1128f | MG-161r |
| 6 | MG-1126s | MG-90c | |
| 7 | Shepherd’s purse (Capsella bursa-pastoris) | SP-1149c | SP-1400l |
| 8 | SP-1101fl | SP-1151f | |
| 9 | Field pansy (Viola arvensis) | FP-1110f | FP-541s |
| 10 | FP-1109s | FP-153l | |
| 11 | Scentless false mayweed (Tripleurospermum inodorum) | SFM-1143s | SFM-1265f |
| 12 | SFM-1118c | SFM-1120c | |
| 13 | Control | Sterile distilled water | |
| Target | Primer Name | Sequence (5′–3′) | Literature Source |
|---|---|---|---|
| F. graminearum | FgramB379 fwd | CCATTCCCTGGGCGCT | [35] |
| FgramB411 rev | CCTATTGACAGGTGGTTAGTGACTGG | ||
| F. avenaceum | Fave574 fwd | TATGTTGTCACTGTCTCACACCACC | |
| Fave627 rev | AGAGGGATGTTAGCATGATGAAG | ||
| Plant EF1α | Hor1fwd | TCTCTGGGTTTGAGGGTGAC | |
| Hor2rev | GGCCCTTGTACCAGTCAAGGT |
| Variables (n = 25) | FHBs (%) | 1000-Grain Weight (g) |
|---|---|---|
| F. avenaceum DNA content (pg/ng) | −0.682 ** | 0.686 ** |
| F. graminearum DNA content (pg/ng) | 0.791 ** | −0.808 ** |
| FHBs (%) | −0.910 ** |
| Host Plant | No. | Strain | DNA Content of F. avenaceum | DNA Content of F. graminearum | FHBs (%) | 1000-Grain Weight (g) | |
|---|---|---|---|---|---|---|---|
| F. graminearum | Spring wheat (Triticum aestivum) | 1 | SW-6K5V1 | 0.007 a | 1277,374 e | 64.8 def | 17.3 ab |
| 2 | SW-6K4V1 | 0.010 a | 2068.344 f | 83.4 hi | 15.9 a | ||
| Wild buckwheat (Fallopia convolvulus) | 3 | WB-544r | 0.022 a | 1126.426 e | 66.0 defg | 18.6 abcd | |
| 4 | WB-144r | 0.004 a | 743.660 bcde | 53.6 d | 21.4 de | ||
| Meadow grass (Poa annua) | 5 | MG-161r | 0.002 a | 406.922 abcd | 76.3 fghi | 17.6 abcd | |
| 6 | MG-90c | 0.004 a | 194.898 ab | 69.0 efg | 20.0 bcde | ||
| Shepherd’s purse (Capsella bursa-pastoris) | 7 | SP-1400l | 0.010 a | 909.203 de | 78.5 ghi | 20.0 bcde | |
| 8 | SP-1151f | 0.049 a | 410.992 abcd | 57.9 de | 20.6 cde | ||
| Field pansy (Viola arvensis) | 9 | FP-541s | 0.015 a | 770.917 cde | 67.0 efg | 21.4 de | |
| 10 | FP-153l | 0.002 a | 839.418 cde | 84.1 hi | 17.8 abc | ||
| Scentless false mayweed (Tripleurospermum inodorum) | 11 | SFM-1265f | 0.050 a | 815.719 cde | 72.1 fghi | 23.0 ef | |
| 12 | SFM-1120c | 0.423 a | 348.702 abc | 71.3 fghi | 22.8 ef | ||
| F. avenaceum | Spring wheat (Triticum aestivum) | 13 | SW-G1-II | 11.846 bcd | 0.566 a | 7.5 ab | 34.0 k |
| 14 | SW-TG5-IV | 6.201 ab | 0.414 a | 10.8 ab | 30.2 i | ||
| Wild buckwheat (Fallopia convolvulus) | 15 | WB-1180l | 52.153 f | 5.860 a | 14.3 b | 31.2 ijk | |
| 16 | WB-1178fl | 12.374 bcd | 0.253 a | 27.2 c | 25.0 fg | ||
| Meadow grass (Poa annua) | 17 | MG-1128f | 7.311 abc | 0.291 a | 3.5 ab | 26.6 gh | |
| 18 | MG-1126s | 19.860 de | 2.995 a | 4.9 ab | 28.4 hi | ||
| Shepherd’s purse (Capsella bursa-pastoris) | 19 | SP-1149c | 19.029 de | 0.334 a | 1.2 a | 31.3 ijk | |
| 20 | SP-1101fl | 20.374 de | 2.092 a | 1.7 a | 28.5 hi | ||
| Field pansy (Viola arvensis) | 21 | FP-1110f | 17.313 cde | 17.233 a | 1.8 a | 30.2 i | |
| 22 | FP-1109s | 23.574 e | 1.682 a | 2.4 ab | 29.3 hi | ||
| Scentless false mayweed (Tripleurospermum inodorum) | 23 | SFM-1143s | 19.536 de | 6.724 a | 1.4 a | 26.1 fgh | |
| 24 | SFM-1118c | 26.618 e | 0.381 a | 1.1 a | 28.4 hi | ||
| Control | H2O | 25 | - | 0.014 a | 0.593 a | 0.4 a | 29.4 hi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matelionienė, N.; Supronienė, S.; Shamshitov, A.; Zavtrikovienė, E.; Janavičienė, S.; Kadžienė, G. Weeds in Cereal Crop Rotations May Host Fusarium Species That Cause Fusarium Head Blight and Grain Weight Losses in Wheat. Agronomy 2022, 12, 2741. https://doi.org/10.3390/agronomy12112741
Matelionienė N, Supronienė S, Shamshitov A, Zavtrikovienė E, Janavičienė S, Kadžienė G. Weeds in Cereal Crop Rotations May Host Fusarium Species That Cause Fusarium Head Blight and Grain Weight Losses in Wheat. Agronomy. 2022; 12(11):2741. https://doi.org/10.3390/agronomy12112741
Chicago/Turabian StyleMatelionienė, Neringa, Skaidrė Supronienė, Arman Shamshitov, Evelina Zavtrikovienė, Sigita Janavičienė, and Gražina Kadžienė. 2022. "Weeds in Cereal Crop Rotations May Host Fusarium Species That Cause Fusarium Head Blight and Grain Weight Losses in Wheat" Agronomy 12, no. 11: 2741. https://doi.org/10.3390/agronomy12112741
APA StyleMatelionienė, N., Supronienė, S., Shamshitov, A., Zavtrikovienė, E., Janavičienė, S., & Kadžienė, G. (2022). Weeds in Cereal Crop Rotations May Host Fusarium Species That Cause Fusarium Head Blight and Grain Weight Losses in Wheat. Agronomy, 12(11), 2741. https://doi.org/10.3390/agronomy12112741