
Role of Glycine Betaine in the Thermotolerance of Plants




Abstract
:1. Introduction
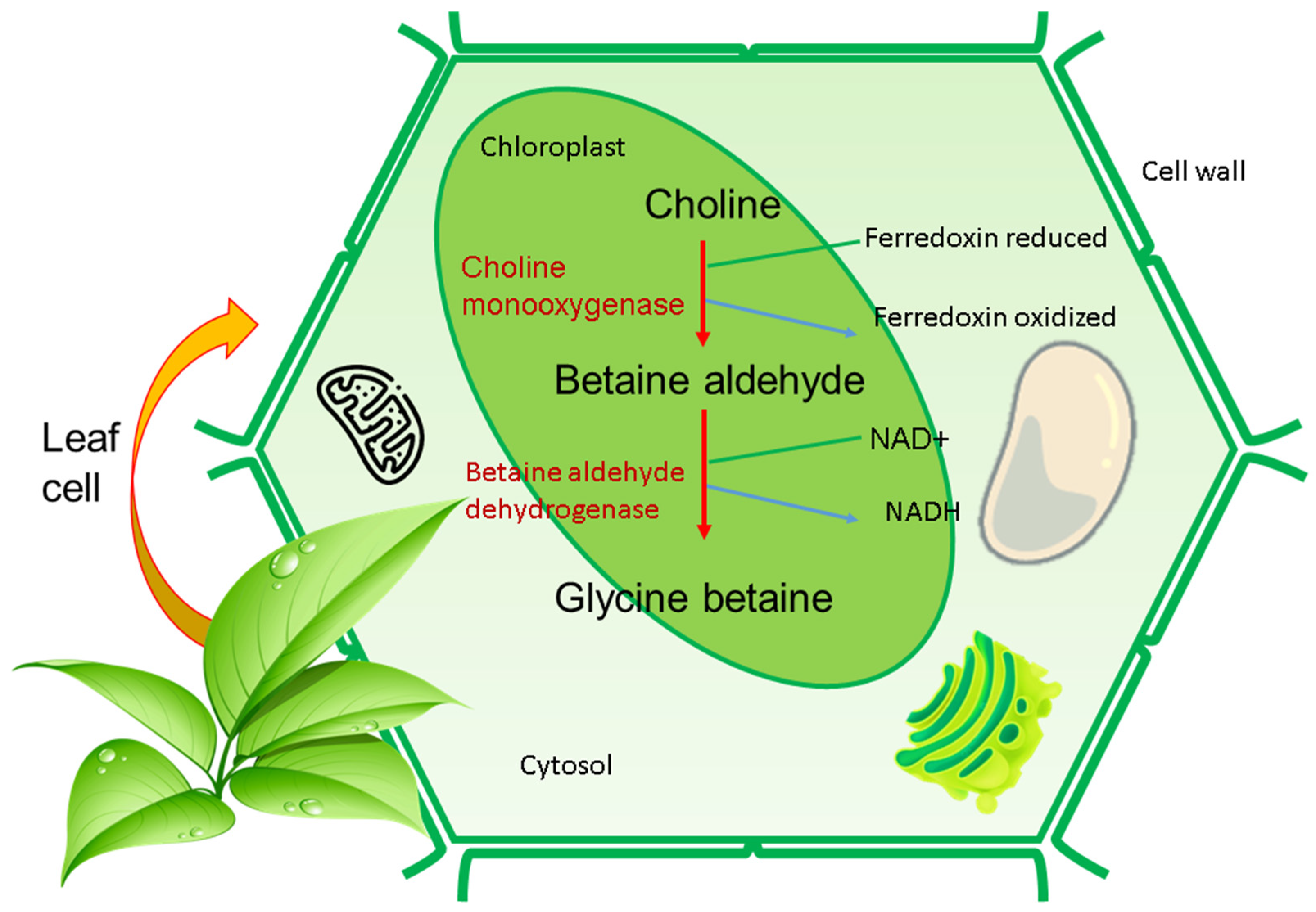
2. GB Structure and Biosynthesis in Plants
3. Glycine Betaine-Accumulating and -Non-Accumulating Plants
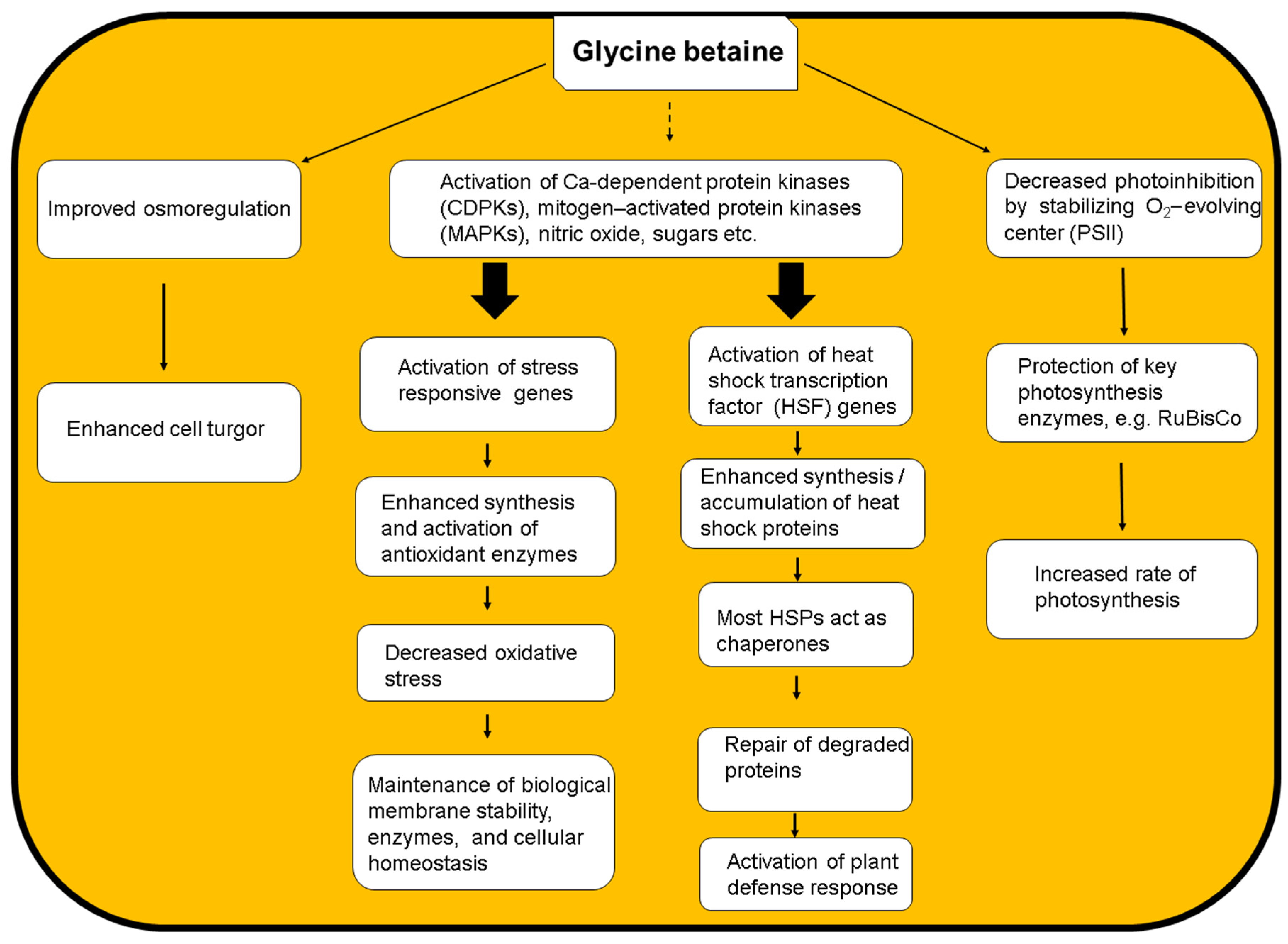
4. Mechanisms of GB-Mediated Thermotolerance
5. Improvement in Heat Tolerance through Exogenous Application of GB
6. Genetic Engineering for Enhanced Thermotolerance
7. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geange, S.R.; Arnold, P.A.; Catling, A.A.; Coast, O.; Cook, A.M.; Gowland, K.M.; Leigh, A.; Notarnicola, R.F.; Posch, B.C.; Venn, S.E.; et al. The thermal tolerance of photosynthetic tissues: A global systematic review and agenda for future research. New Phytol. 2020, 229, 2497–2513. [Google Scholar] [CrossRef]
- Gil, K.E.; Park, C.M. Thermal adaptation and plasticity of the plant circadian clock. New Phytol. 2018, 221, 1215–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhao, C.; Müller, C.; Wang, C.; Ciais, P.; Janssens, I.; Peñuelas, J.; Asseng, S.; Li, T.; Elliott, J.; et al. Emergent constraint on crop yield response to warmer temperature from field experiments. Nat. Sustain. 2020, 3, 908–916. [Google Scholar] [CrossRef]
- Sadok, W.; Lopez, J.R.; Smith, K.P. Transpiration increases under high-temperature stress: Potential mechanisms, trade-offs and prospects for crop resilience in a warming world. Plant Cell Environ. 2021, 44, 2102–2116. [Google Scholar] [CrossRef]
- IPCC. Climate change 2014: Impacts, adaptation and vulnerability. In Working Group II Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Ferguson, J.N.; Tidy, A.C.; Murchie, E.H.; Wilson, Z.A. The potential of resilient carbon dynamics for stabilizing crop reproductive development and productivity during heat stress. Plant Cell Environ. 2021, 44, 2066–2089. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Carter, T.R.; Ebi, K.; Harrison, P.A. Achievements and needs for the climate change scenario framework. Nat. Clim. Chang. 2020, 10, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Hertel, T.W.; de Lima, C.Z. Climate impacts on agriculture: Searching for keys under the streetlight. Food Policy 2020, 95, 101954. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.L.; Yang, Y.J.; Liu, T.; Zhang, S.B.; Huang, W. Responses of photosystem I compared with photosystem II to combination of heat stress and fluctuating light in tobacco leaves. Plant Sci. 2020, 292, 110371. [Google Scholar] [CrossRef]
- Parrotta, L.; Aloisi, I.; Faleri, C.; Romi, M.; Del Duca, S.; Cai, G. Chronic heat stress affects the photosynthetic apparatus of Solanum lycopersicum L. cv Micro-Tom. Plant Physiol. Biochem. 2020, 154, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Katano, K. Coordination between ROS regulatory systems and other pathways under heat stress and pathogen attack. Front Plant Sci. 2018, 9, 490. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Yang, W.J.; Lo, H.F.; Yeh, D.M. Physiology, anatomy, and cell membrane thermostability selection of leafy radish (Raphanus sativus var. oleiformis Pers.) with different tolerance under heat stress. Sci. Hortic. 2014, 179, 367–375. [Google Scholar] [CrossRef]
- Akter, N.; Islam, M.R. Heat stress effects and management in wheat. A review. Agron. Sustain. Dev. 2017, 37, 37. [Google Scholar] [CrossRef]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins–evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, R.; Ahmad, N.; Wyn Jones, R.G. Taxonomic and ecological aspects of the distribution of glycinebetaine and related compounds in plants. Oecologia 1977, 27, 319–332. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Los, D.A.; Mohanty, P.; Nishiyama, Y.; Murata, N. Glycinebetaine alleviates the inhibitory effect of moderate heat stress on the repair of photosystem II during photoinhibition. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, S.; Zhang, H.; Zhao, Y.; Zhao, H.; Liu, H. Glycine betaine application in grain filling wheat plants alleviates heat and high light-induced photoinhibition by enhancing the psbA transcription and stomatal conductance. Acta Physiol. Plant 2014, 36, 2195–2202. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Alhaithloul, H.A.; Soliman, M.H.; Ameta, K.L.; El-Esawi, M.A.; Elkelish, A. Changes in ecophysiology, osmolytes, and secondary metabolites of the medicinal plants of Mentha piperita and Catharanthus roseus subjected to drought and heat stress. Biomolecules 2020, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Sorwong, A.; Sakhonwasee, S. Foliar application of glycine betaine mitigates the effect of heat stress in three marigold (Tagetes erecta) cultivars. Hort. J. 2015, 48, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef]
- Hanson, A.D.; Rhodes, D. 14C tracer evidence for synthesis of choline and betaine via phosphoryl base intermediates in salinized sugarbeet leaves. Plant Physiol. 1983, 71, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Weretilnyk, E.A.; Bednarek, S.; McCue, K.F.; Rhodes, D.; Hanson, A.D. Comparative biochemical and immunological studies of the glycine betaine synthesis pathway in diverse families of dicotyledons. Planta 1989, 178, 342–352. [Google Scholar] [CrossRef]
- Chen, T.H.; Murata, N. Glycinebetaine: An effective protectant against abiotic stress in plants. Trends Plant Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Brouquisse, R.; Weigel, P.; Rhodes, D.; Yocum, C.F.; Hanson, A.D. Evidence for a ferredoxin-dependent choline monooxygenase from spinach chloroplast stroma. Plant Physiol. 1989, 90, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibino, T.; Waditee, R.; Araki, E.; Ishikawa, H.; Aoki, K.; Tanaka, Y.; Takabe, T. Functional characterization of choline monooxygenase, an enzyme for betaine synthesis in plants. J. Biol. Chem. 2002, 277, 41352–41360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathinasabapathi, B.; Burnet, M.; Russell, B.L. Choline monooxygenase, an unusual iron-sulfur enzyme catalyzing the first step of glycine betaine synthesis in plants: Prosthetic group characterization and cDNA Cloning. Proc. Natl. Acad. Sci. USA 1997, 94, 3454–3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Chen, T.H.; Murata, N. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol. 2002, 5, 250–257. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Glycinebetaine in saline conditions: An assessment of the current state of knowledge. Acta Physiol. Plant. 2017, 39, 56. [Google Scholar] [CrossRef]
- Chen, T.H.; Murata, N. Glycinebetaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell Environ. 2011, 34, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hirji, R.; Adam, L.; Rozwadowski, K.L.; Hammerlindl, J.K.; Keller, W.A.; Selvaraj, G. Genetic engineering of glycinebetaine production toward enhancing stress tolerance in plants: Metabolic limitations. Plant Physiol. 2000, 122, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, D.; Hanson, A. Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 357–384. [Google Scholar] [CrossRef]
- Wyn Jones, R.G.; Storey, R. Betaines. In The Physiology and Biochemistry of Drought Resistance in Plants; Paleg, L.G., Aspinall, D., Eds.; Academic Press: Cambridge, MA, USA, 1981; pp. 171–204. [Google Scholar]
- Ladyman, J.A.R. The accumulation of glycinebetaine in barley in relation to water stress. Diss. Abs. Bull. 1982, 43, 1320. [Google Scholar]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible solute engineering in plants for abiotic stress tolerance role of glycine betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Wang, W.; Liang, C.; Wang, X.; Wang, G.; Wang, W. Overaccumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop J. 2017, 5, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Weimberg, R.; Lerner, H.R.; Poljakoff-Mayber, A. Changes in growth and water soluble solute concentrations in Sorghum bicolor stressed with sodium and potassium. Physiol. Plant. 1984, 62, 472–480. [Google Scholar] [CrossRef]
- Yang, W.-J.; Rich, P.J.; Axtell, J.D.; Wood, K.V.; Bonham, C.C.; Ejeta, G.; Mickelbart, M.V.; Rhodes, D. Genotypic variation for glycine betaine in sorghum. Crop Sci. 2003, 43, 162–169. [Google Scholar] [CrossRef]
- McCue, R.F.; Hanson, A.D. Drought and salt tolerance: Towards understanding and application. Tibtech 1990, 8, 358–362. [Google Scholar] [CrossRef]
- Kishitani, S.; Watanabe, K.; Yasuda, S.; Arakawa, K.; Takabe, T.J.P.C. Accumulation of glycinebetaine during cold acclimation and freezing tolerance in leaves of winter and spring barley plants. Plant Cell Environ. 1994, 17, 89–95. [Google Scholar] [CrossRef]
- Quan, R.; Shang, M.; Zhang, H.; Zhao, Y.; Zhang, J. Engineering of enhanced glycine betaine synthesis improves drought tolerance in maize. Plant Biotechnol. J. 2004, 2, 477–486. [Google Scholar] [CrossRef] [PubMed]
- McNeil, S.D.; Nuccio, M.L.; Hanson, A.D. Betaines and related osmoprotectants. Targets for metabolic engineering of stress resistance. Plant Physiol. 1999, 120, 945–949. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Takabe, T.; Takabe, T.; Kishitani, S. Accumulation of glycinebetaine in rice plants that overexpress choline monooxygenase from spinach and evaluation of their tolerance to abiotic stress. Ann. Bot. 2006, 98, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkelä, P.; Munns, R.; Colmer, T.D.; Condon, A.G.; Peltonen-Sainio, P. Effect of foliar applications of glycinebetaine on stomatal conductance, abscisic acid and solute concentrations in leaves of salt-or drought-stressed tomato. Funct. Plant Biol. 1998, 25, 655–663. [Google Scholar] [CrossRef]
- Al-Huqail, A.; El-Dakak, R.M.; Sanad, M.N.; Badr, R.H.; Ibrahim, M.M.; Soliman, D.; Khan, F. Effects of climate temperature and water Stress on plant growth and accumulation of antioxidant compounds in sweet basil (Ocimum basilicum L.) leafy vegetable. Scientifica 2020, 2020, 3808909. [Google Scholar] [CrossRef] [Green Version]
- Hemantaranjan, A.; Bhanu, A.N.; Singh, M.N.; Yadav, D.K.; Patel, P.K.; Singh, R.; Katiyar, D. Heat stress responses and thermotolerance. Adv. Plants Agric. Res. 2014, 1, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Divya, K.; Bhatnagar-Mathur, P.; Sharma, K.K.; Reddy, P.S. Heat shock proteins (Hsps) mediated signalling pathways during abiotic stress conditions. In Plant Signaling Molecules; Woodhead Publishing: Shaston, UK, 2019; pp. 499–516. [Google Scholar]
- Hashemi-Petroudi, S.H.; Nematzadeh, G.; Mohammadi, S.; Kuhlmann, M. Expression pattern analysis of heat shock transcription factors (HSFs) gene family in Aeluropus littoralis under salinity stress. Environ. Stresses Crop Sci. 2020, 13, 571–581. [Google Scholar]
- Zulfiqar, F.; Ashraf, M. Bioregulators: Unlocking their potential role in regulation of the plant oxidative defense system. Plant Mol. Biol. 2021, 105, 11–41. [Google Scholar] [CrossRef]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Andrási, N.; Pettkó-Szandtner, A.; Szabados, L. Diversity of plant heat shock factors: Regulation, interactions, and functions. J. Exp. Bot. 2020, 72, 1558–1575. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Reddy, P.S.; Chakradhar, T.; Reddy, R.A.; Nitnavare, R.B.; Mahanty, S.; Reddy, M.K. Role of heat shock proteins in improving heat stress tolerance in crop plants. In Heat Shock Proteins and Plants; Springer: Cham, Switzerland, 2016; pp. 283–307. [Google Scholar]
- Yang, G.; Rhodes, D.; Joly, R.J. Effects of high temperature on membrane stability and chlorophyll fluorescence in glycinebetaine-deficient and glycinebetaine-containing maize lines. Funct. Plant Biol. 1996, 23, 437–443. [Google Scholar] [CrossRef]
- Jagadish, S.K.; Way, D.A.; Sharkey, T.D. Plant heat stress: Concepts directing future research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, F.; Wang, J.; Zhang, W.; Meng, Q.; Chen, T.H.H.; Murata, N.; Yang, X. Glycinebetaine enhances the tolerance of tomato plants to high temperature during germination of seeds and growth of seedlings. Plant Cell Environ. 2011, 34, 1931–1943. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A.; El Madidi, S.; Strasser, R.J. Exogenous glycine betaine and proline play a protective role in heat-stressed barley leaves (Hordeum vulgare L.): A chlorophyll a fluorescence study. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2012, 146, 1037–1043. [Google Scholar]
- Wahid, A.; Shabbir, A. Induction of heat stress tolerance in barley seedlings by pre-sowing seed treatment with glycinebetaine. Plant Growth Regul. 2005, 46, 133–141. [Google Scholar] [CrossRef]
- Rasheed, R.; Wahid, A.; Farooq, M.; Hussain, I.; Basra, S.M. Role of proline and glycinebetaine pretreatments in improving heat tolerance of sprouting sugarcane (Saccharum sp.) buds. Plant Growth Regul. 2011, 65, 35–45. [Google Scholar] [CrossRef]
- Makela, P.; Jokinen, K.; Kontturi, M.; Peltonen-Sainio, P.; Pehu, E.; Somersalo, S. Foliar application of glycine betaine—A novel product from sugar beet as an approach to increase tomato yield. Ind. Crops Prod. 1998, 7, 139–148. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.; Xue, X.; Lu, C.; Chen, R.; Wang, L.; Han, X. Foliar spraying of glycine betaine lowers photosynthesis inhibition of Malus hupehensis leaves under drought and heat stress. Int. J. Agric. Biol. 2020, 23, 1121–1128. [Google Scholar]
- Chowdhury, A.R.; Ghosh, M.; Lal, M.; Pal, A.; Hazra, K.K.; Acharya, S.; Chaurasiya, A.; Pathak, S.K. Foliar spray of synthetic osmolytes alleviates terminal heat stress in late-sown wheat. Int. J. Plant Prod. 2020, 14, 321–333. [Google Scholar] [CrossRef]
- Li, M.; Li, Z.; Li, S.; Guo, S.; Meng, Q.; Li, G.; Yang, X. Genetic Engineering of glycine betaine biosynthesis reduces heat-enhanced photoinhibition by enhancing antioxidative defense and alleviating lipid peroxidation in tomato. Plant Mol. Biol. Rep. 2014, 32, 42–51. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Z.; Li, D.; Li, C.; Wei, D.; Li, S.; Liu, Y.; Chen, T.H.H.; Yang, X. Comparative effects of glycinebetaine on the thermotolerance in codA-and BADH-transgenic tomato plants under high temperature stress. Plant Cell Rep. 2020, 39, 1525–1538. [Google Scholar] [CrossRef]
- Yang, X.H.; Liang, Z.; Lu, C.M. Genetic engineering of the biosynthesis of glycinebetaine enhances photosynthesis against high temperature stress in transgenic tobacco plants. Plant Physiol. 2005, 138, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wen, X.; Gong, H.; Lu, Q.; Yang, Z.; Tang, Y.; Liang, Z.; Lu, C. Genetic engineering of the biosynthesis of glycinebetaine enhances thermotolerance of photosystem II in tobacco plants. Planta 2007, 225, 719–733. [Google Scholar] [CrossRef]
- Yu, H.Q.; Wang, Y.G.; Yong, T.M.; She, Y.H.; Fu, F.L.; Li, W.C. Heterologous expression of betaine aldehyde dehydrogenase gene from Ammopiptanthus nanus confers high salt and heat tolerance to Escherichia coli. Gene 2014, 549, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.P.; Li, F.; Zhang, J.; Zhao, M.R.; Hui, Z.; Wang, W. Overaccumulation of glycine betaine enhances tolerance of the photosynthetic apparatus to drought and heat stress in wheat. Photosynthetica 2010, 48, 30–41. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| GB Accumulators | Accumulating Condition | References |
|---|---|---|
| Plant families with known naturally high accumulation of GB: Asteraceae, Chenopodiaceae, Poaceae, and Solanaceae | Different types of stresses | [37,38,39] |
| Spinach (Spinacia oleracea) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditions | [31,32,36] |
| Sugar beet (Beta vulgaris) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditions | [31,32,36] |
| Barley (Hordeum vulgare) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditions | [31,36,40] |
| Wheat (Triticum aestivum) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditions | [31,36,40] |
| Sorghum (Sorghum bicolor) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditions | [41,42] |
| Maize (Zea mays) | Naturally accumulates under non-stress conditions; GB levels increase under stress conditons | [36,43,44,45] |
| GB-non-accumulators | ||
| Rice (Oryza sativa) | Non-stress and stress conditions | [31,36,46,47] |
| Mustard (Brassica spp.) | Non-stress and stress conditions | [46] |
| Arabidopsis (Arabidopsis thaliana) | Non-stress and stress conditions | [33,36,46] |
| Tobacco (Nicotiana tabacum) | Non-stress and stress conditions | [33,46] |
| Tomato (Solanum lycopersicum) | Non-stress and stress conditions | [33,36,46,48] |
| Potato (Solanum tuberosum) | Non-stress and stress conditions | [36,46] |
| Crop | Heat Stress Range | GB Concentration Applied | Exogenously Applied GB-Induced Regulation of Different Attributes in Heat-Stressed Plants | Reference |
|---|---|---|---|---|
| Tomato (Lycopersicon esculentum) | 34 °C | 0, 0.1, 1, and 5 mM GB |
| [60] |
| Barley (Hordeum vulgare) | 45 °C | 10 mM |
| [61] |
| Wheat (Triticum aestivum) | 25/20 °C day/night | 100 mM |
| [20] |
| Wheat (T. aestivum) | 30–38 °C | 100 and 50 mM |
| [66] |
| Barley (H. vulgare) | 40/32 °C day/night | 10, 20, 30, 40, and 50 mM |
| [62] |
| Sugarcane (Saccharum spp.) | 42 °C | 20 mM |
| [63] |
| Marigold (Tagetes erecta) | 39/29 °C day/night | 0.5 and 1 mM |
| [23] |
| Gene Transformed | Donor/Source | Gene Action | Transgenic Plant Species | Stress Condition | Transgenic Plant Response | References |
|---|---|---|---|---|---|---|
| BADH and codA | Spinach (Spinacia oleracea L.) as a donor of the BADH gene; binary vector pCG/codA for chloroplast-targeted expression of the codA gene | Genes related to key enzymes involved in GB synthesis | Tomato (Solanum lycopersicum) | Two months after transplanting, plants were exposed to 42 °C for 0–8 h in a growth chamber |
| [68] |
| BADH | Spinach (S. oleracea L.) | Gene for betaine aldehyde dehydrogenase | Tobacco (Nicotiana tabacum) | Two-month-old seedlings subjected to various temperatures (25–50 °C) for 4 h in a growth chamber |
| [70] |
| BADH | Spinach (S. oleracea L.) | Gene for betaine aldehyde dehydrogenase | Tomato (S. lycopersicum) | Six-week-old seedlings placed in a growth chamber at 42 °C for 0, 2, 4, or 6 h |
| [67] |
| BADH | Spinach (S. oleracea L.) | ---do--- | Tomato (Solanum lycopersicum) | Two-month-old plants exposed to varying temperature regimes (25–45 °C) for 2 h |
| [69] |
| BADH | Garden orache (Atriplex hortensis L.) | --do-- | Wheat (Triticum aestivum) | Individual and combined heat stress (40 °C) and drought stress as PEG-6000 (osmotic potential about −1.88 MPa) for 3 h in an artificial chamber |
| [72] |
| codA | Spinach (S. oleracea L.) | Gene encodescholine oxidase (COD), a key enzyme for GB synthesis | Rice (Oryza sativa) | Plants grown at 28/13 °C for 5 weeks |
| [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulfiqar, F.; Ashraf, M.; Siddique, K.H.M. Role of Glycine Betaine in the Thermotolerance of Plants. Agronomy 2022, 12, 276. https://doi.org/10.3390/agronomy12020276
Zulfiqar F, Ashraf M, Siddique KHM. Role of Glycine Betaine in the Thermotolerance of Plants. Agronomy. 2022; 12(2):276. https://doi.org/10.3390/agronomy12020276
Chicago/Turabian StyleZulfiqar, Faisal, Muhammad Ashraf, and Kadambot H. M. Siddique. 2022. "Role of Glycine Betaine in the Thermotolerance of Plants" Agronomy 12, no. 2: 276. https://doi.org/10.3390/agronomy12020276
APA StyleZulfiqar, F., Ashraf, M., & Siddique, K. H. M. (2022). Role of Glycine Betaine in the Thermotolerance of Plants. Agronomy, 12(2), 276. https://doi.org/10.3390/agronomy12020276