
Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Plant Cultivation
2.2. Cadmium Treatments
2.3. Estimation of Leaf Gas-Exchange Parameters and Instantaneous WUE
2.4. Estimation of Relative Water Content
2.5. Determination of Biomass, Cd Concentration, and Cd Accumulation
2.6. Measurements of Long-Term WUE
2.7. Measurements for Abscisic Acid Content
2.8. RNA Isolation and Quantitative PCR Analysis
2.9. Statistical Analysis
3. Results
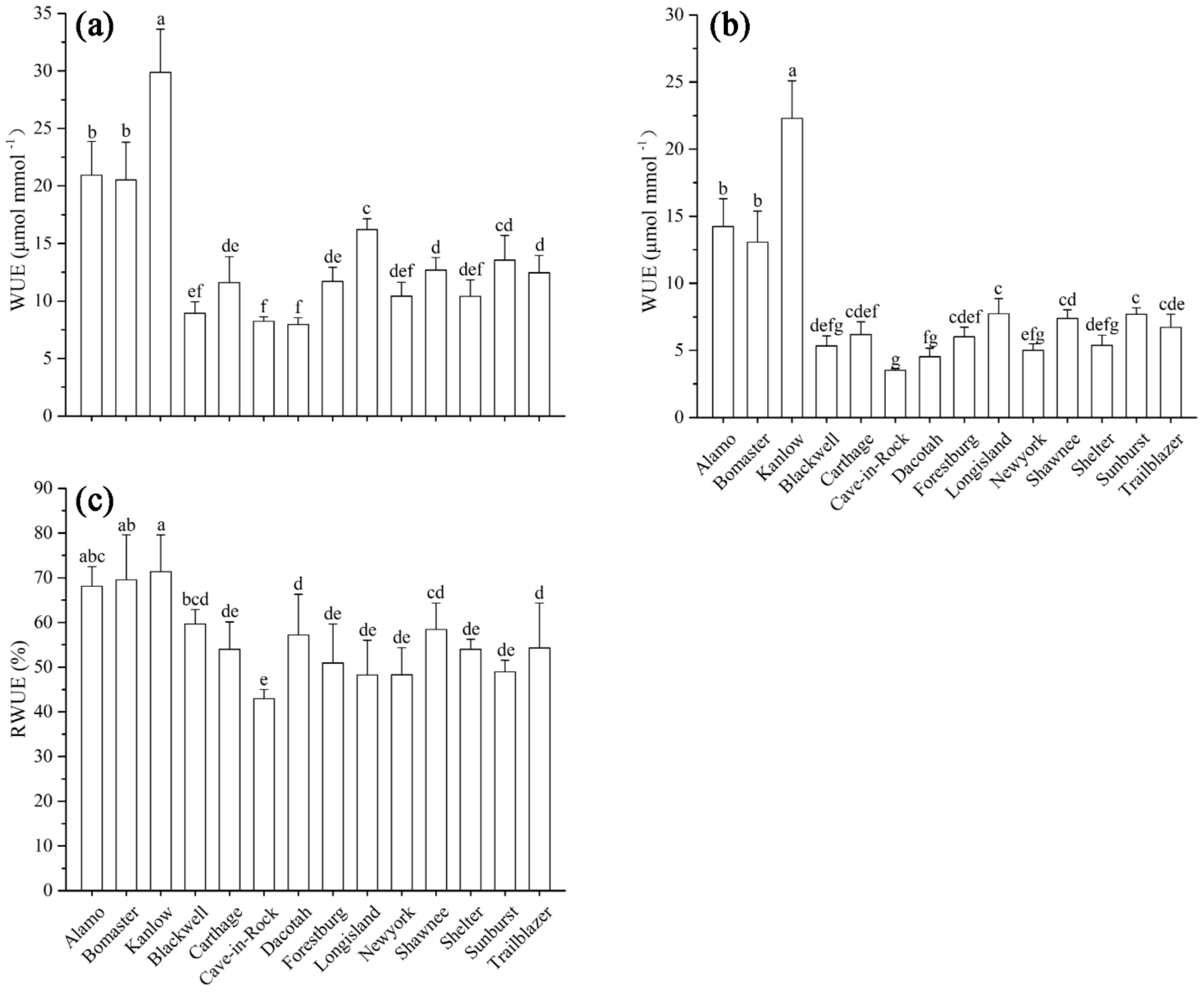
3.1. Effects of Cd on Instantaneous WUE (iWUE) of Switchgrass
3.2. Effects of Cd on the RWC of Switchgrass
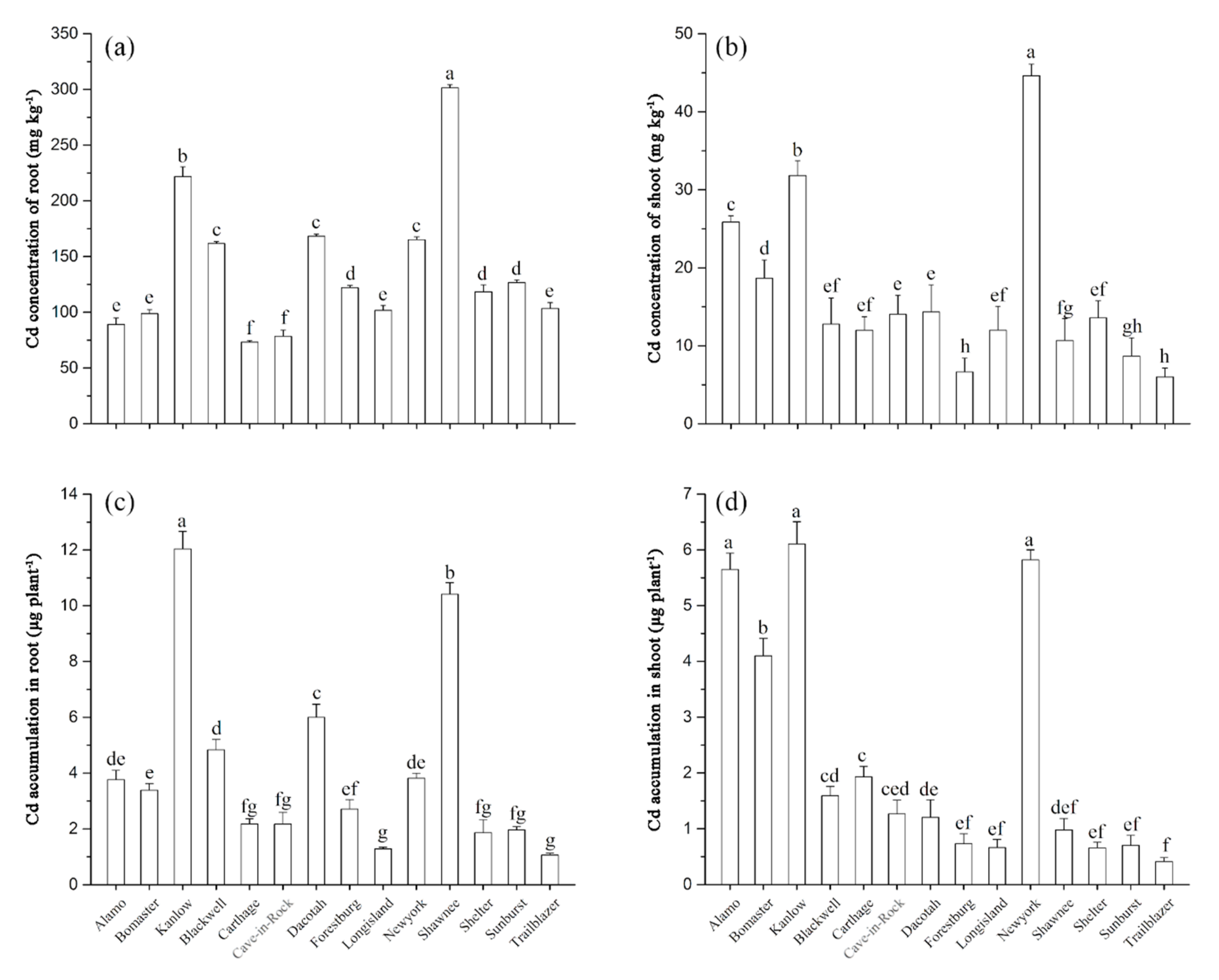
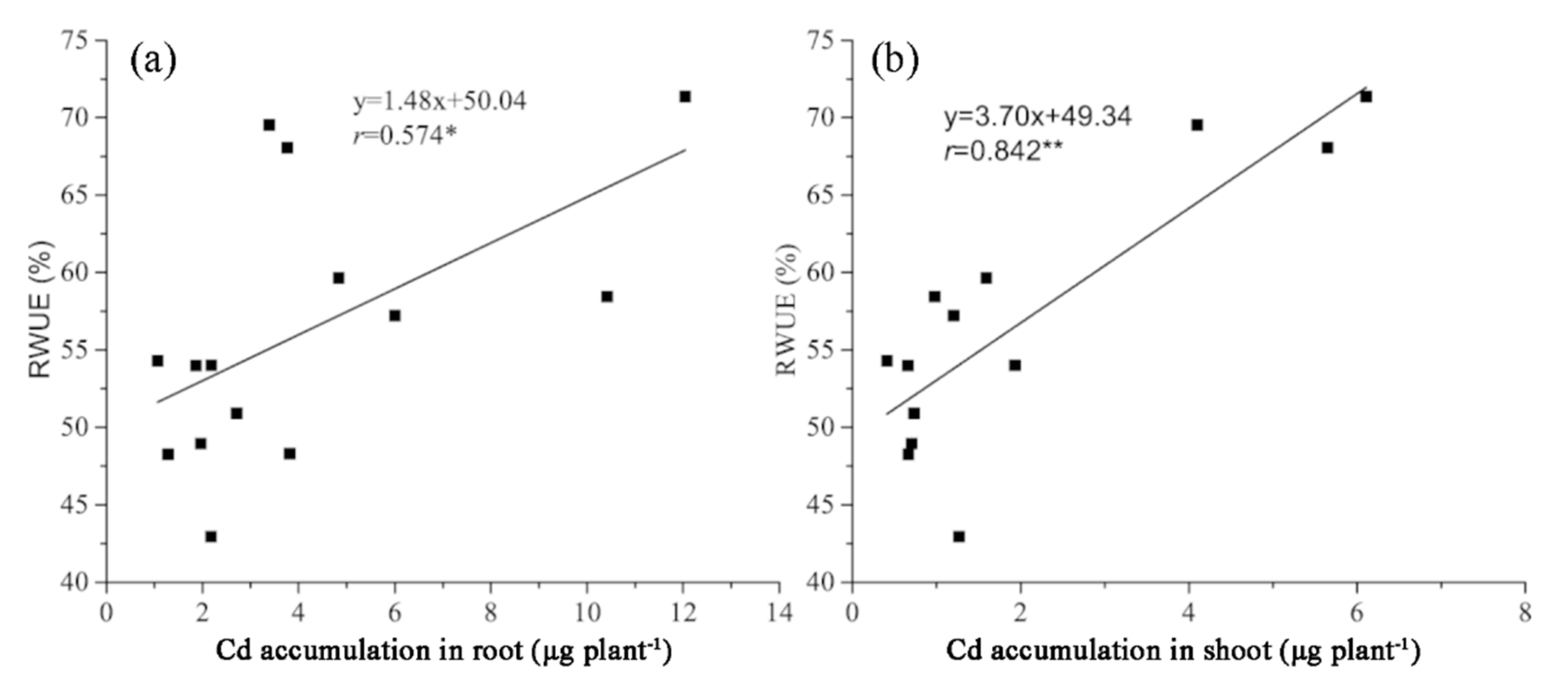
3.3. Cd Concentration, Cd Accumulations, and Their Relationship with RWUE
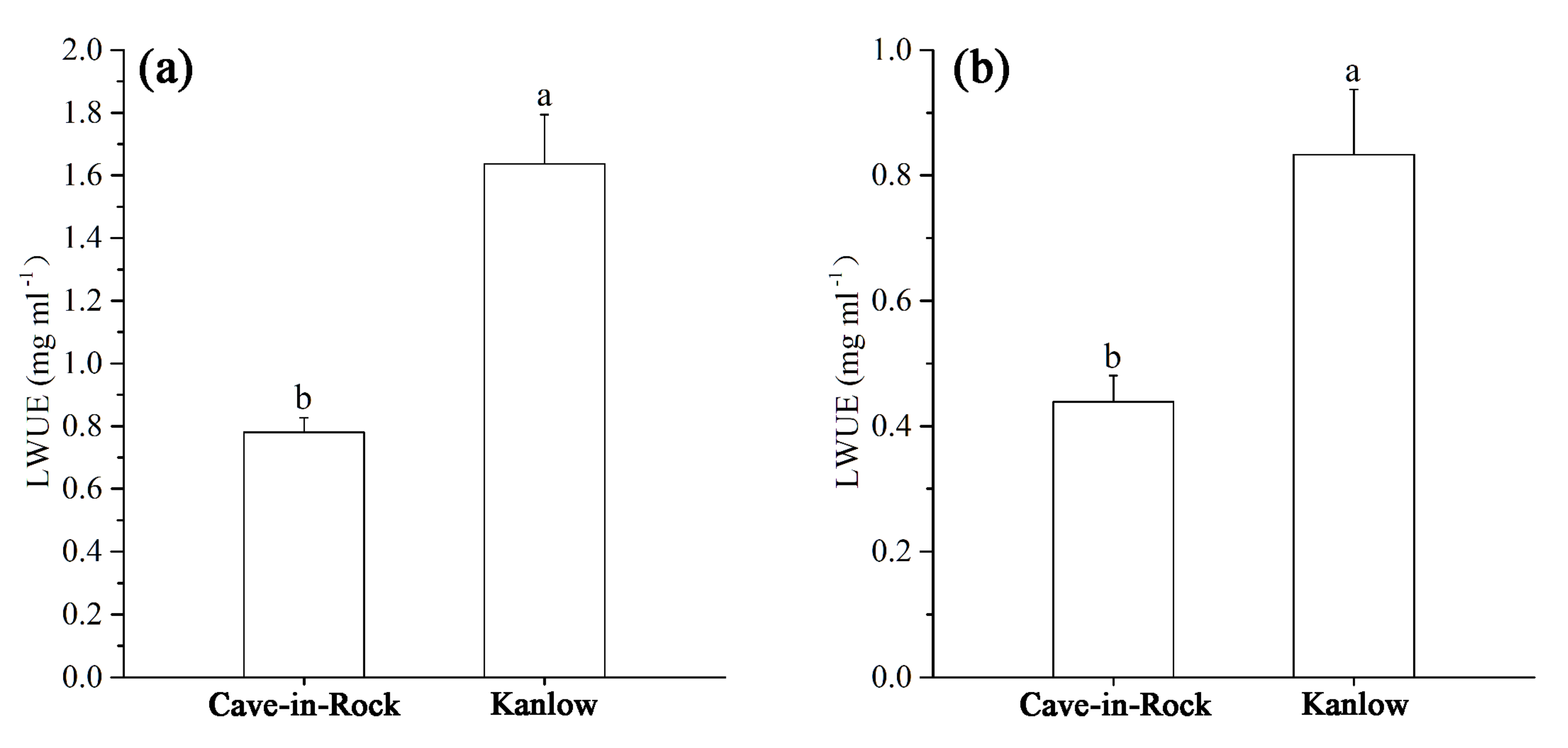
3.4. Effects of Cd on Long-Term WUE of Switchgrass
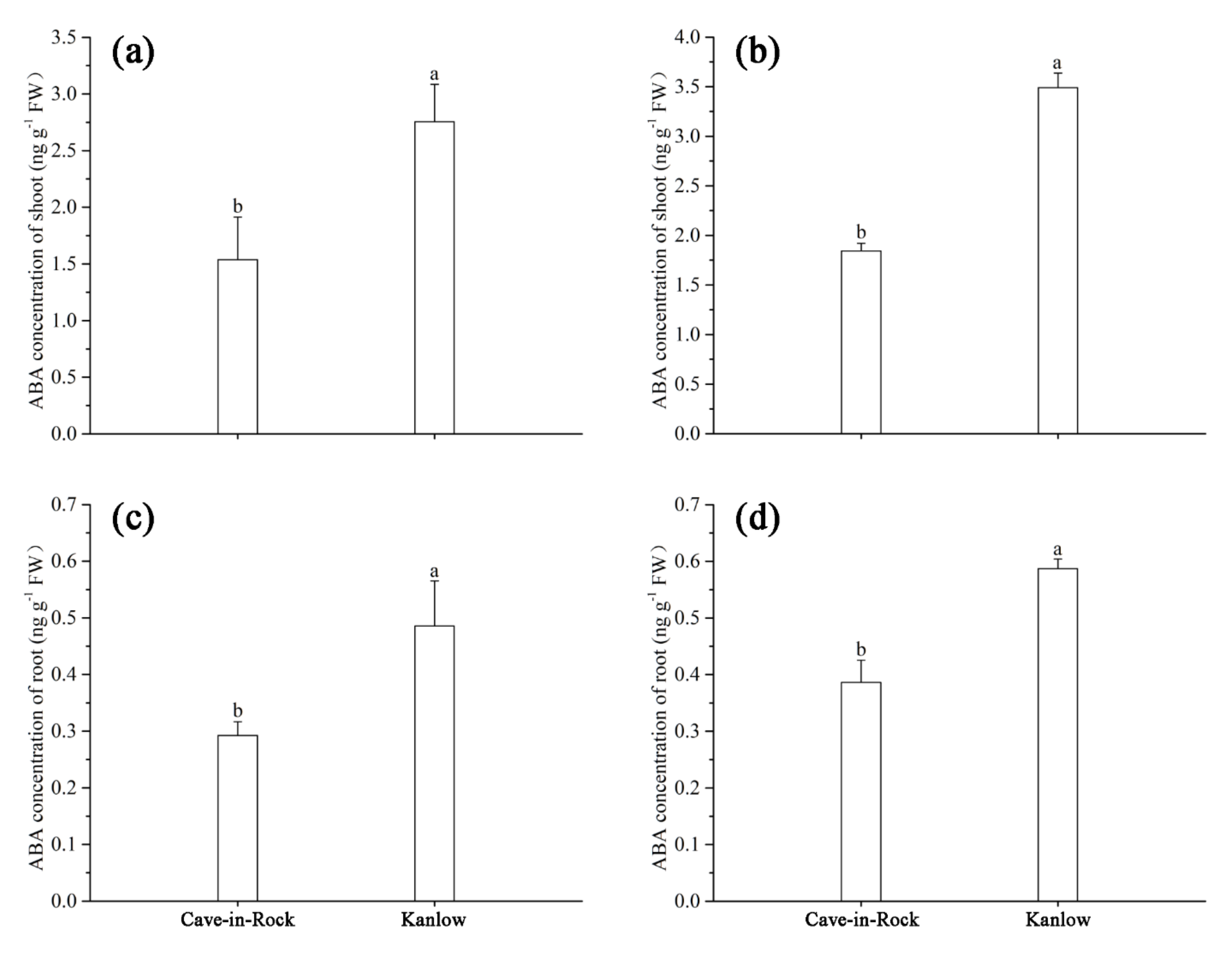
3.5. Effects of Cd on ABA Concentration of Switchgrass
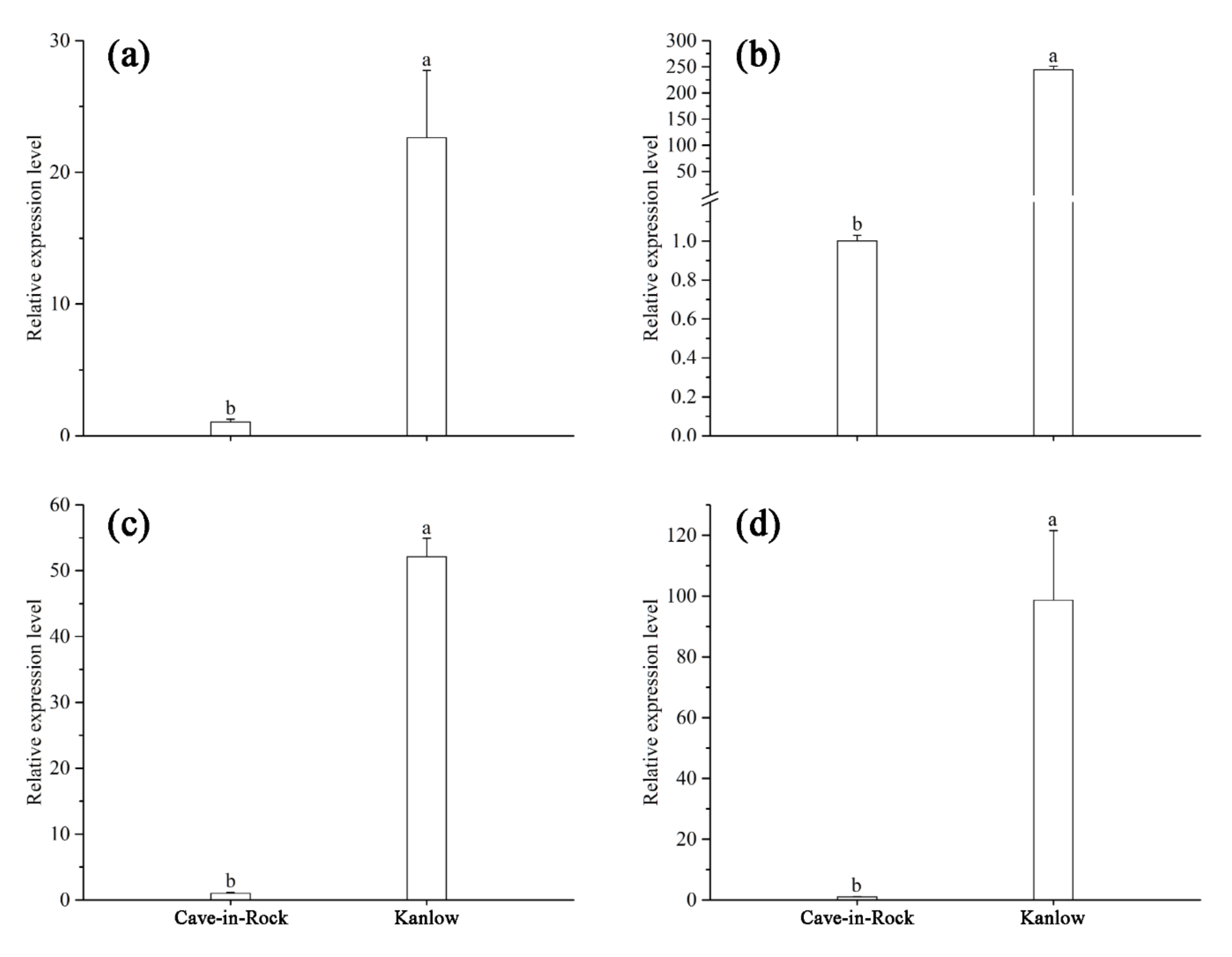
3.6. Relative Expression of PvERECTA and PvEPF1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, S.; Xu, B.; Zhang, J.; Xie, Z.; Cheng, Q.; Yang, Z.; Cai, Q.; Huang, B. Comprehensive analysis of CCCH-type zinc finger family genes facilitates functional gene discovery and reflects recent allopolyploidization event in tetraploid switchgrass. BMC Genom. 2015, 16, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsamo, R.A.; Kelly, W.J.; Satrio, J.A.; Ruiz-Felix, M.N.; Fetterman, M.; Wynn, R.; Hagel, K. Utilization of grasses for potential biofuel production and phytoremediation of heavy metal contaminated soils. Int. J. Phytoremediat. 2015, 17, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Mielenz, J.R.; Xiao, X.; Ge, Y.; Hamilton, C.Y.; Rodriguez, M.; Chen, F.; Foston, M.; Ragauskas, A.; Bouton, J.; et al. Genetic manipulation of lignin reduces recalcitrance and improves ethanol production from switchgrass. Proc. Natl. Acad. Sci. USA 2011, 108, 3803–3808. [Google Scholar] [CrossRef] [Green Version]
- Schmer, M.R.; Vogel, K.P.; Mitchell, R.B.; Perrin, R.K. Net energy of cellulosic ethanol from switchgrass. Proc. Natl. Acad. Sci. USA 2008, 105, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, K.; Sharma, S.; Monti, A. Bio-remediation of Pb and Cd polluted soils by switchgrass: A case study in India. Int. J. Phytoremediation 2015, 18, 704–709. [Google Scholar] [CrossRef]
- Wang, Q.; Gu, M.; Ma, X.; Zhang, H.; Wang, Y.; Cui, J.; Gao, W.; Gui, J. Model optimization of cadmium and accumulation in switchgrass (Panicum virgatum L.): Potential use for ecological phytoremediation in Cd-contaminated soils. Environ. Sci. Pollut. Res. 2015, 22, 16758. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.; Shao, G.; Tang, Z.; Chen, H.; Wang, B.; Tang, Z.; Yang, Y.; Liu, Y.; Zhao, F.J. Genotypic and environmental variations in grain cadmium and arsenic concentrations among a panel of high yielding rice cultivars. Rice 2017, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef]
- The Ministry of Environmental Protection. The Ministry of Land and Resources Report on the National Soil Contamination Survey. Available online: http://www.mee.gov.cn/gkml/sthjbgw/qt/201404/W020140417558995804588.pdf (accessed on 17 April 2014).
- He, S.; Yang, X.; He, Z.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A Review. Pedosphere 2017, 27, 421–438. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-Ur-Rehman, M.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, R.; Zhang, X.; Yu, Y.; Zhao, R.; Li, Z.; Chen, S. Hydrogen sulfide alleviates cadmium toxicity through regulations of cadmium transport across the plasma and vacuolar membranes in Populus euphratica cells. Plant Physiol. Bioch. 2013, 65, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, R.; Bertrand, A.; Reis, R.; Mourato, M.P.; Martins, L.L.; Gonzalez, A. Growth and physiological responses to cadmium stress of two populations of Dittrichia viscosa (L.) Greuter. J. Hazard. Mater. 2013, 244–245, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Kupper, H.; Parameswaran, A.; Leitenmaier, B.; Trtilek, M.; Setlik, I. Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol. 2007, 175, 655–674. [Google Scholar] [CrossRef]
- Benakova, M.; Ahmadi, H.; Ducaiova, Z.; Tylova, E.; Clemens, S.; Tuma, J. Effects of Cd and Zn on physiological and anatomical properties of hydroponically grown Brassica napus plants. Environ. Sci. Pollut. Res. 2017, 24, 20705–20716. [Google Scholar] [CrossRef]
- Ruggiero, A.; Punzo, P.; Landi, S.; Costa, A.; Oosten, M.; Grillo, S. Improving plant water use efficiency through molecular genetics. Horticulturae 2017, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.; Sallam, A.; Munir, M.; Ali-Dinar, H. Effects of deficit irrigation scheduling on water use, gas exchange, yield, and fruit quality of date palm. Agronomy 2021, 11, 2256. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Franks, P.J.; Doheny-Adams, T.; Britton-Harper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef]
- Liu, C.; Lou, L.; Deng, J.; Li, D.; Yuan, S.; Cai, Q. Morph-physiological responses of two switchgrass (Panicum virgatum L.) cultivars to cadmium stress. Grassl. Sci. 2016, 62, 92–101. [Google Scholar] [CrossRef]
- Pandey, V.C.; Bajpai, O.; Singh, N. Energy crops in sustainable phytoremediation. Renew. Sustain. Energy Rev. 2016, 54, 58–73. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Davis, E.B.; Borsuk, M.E.; Gunderson, C.A.; Lynd, L. Biomass production in switchgrass across the United States: Database description and determinants of yield. Agron. J. 2010, 102, 1158–1168. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Wang, Y.; Wang, J.W.; Babla, M.; Zhao, C.; Garcia-Mata, C.; Sani, E.; Differ, C.; Mak, M.; Hills, A.; et al. Nitrate reductase mutation alters potassium nutrition as well as nitric oxide-mediated control of guard cell ion channels in Arabidopsis. New Phytol. 2016, 209, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Wu, L.; Filardo, F.; Yang, X.; Zhao, X.; Fu, D. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant 2017, 39, 125. [Google Scholar] [CrossRef]
- Shi, G.; Xia, S.; Ye, J.; Huang, Y.; Liu, C.; Zhang, Z. PEG-simulated drought stress decreases cadmium acccumulation in castor bean by altering root morphology. Environ. Exp. Bot. 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Yoon, J.; Cao, X.; Zhou, Q.; Ma, L.Q. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef]
- Fang, Z.; Lou, L.; Tai, Z.; Wang, Y.; Yang, L.; Hu, Z.; Cai, Q. Comparative study of Cd uptake and tolerance of two Italian ryegrass (Lolium multiflorum) cultivars. PeerJ 2017, 5, e3621. [Google Scholar] [CrossRef] [Green Version]
- Wituszynska, W.; Karpiński, S. Determination of Water Use Efficiency for Arabidopsis thaliana. Bio-Protocol 2014, 4, e1041. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Song, G.; Yuan, S.; Wen, X.; Xie, Z.; Lou, L.; Hu, B.; Cai, Q.; Xu, B. Transcriptome analysis of Cd-treated switchgrass root revealed novel transcripts and the importance of HSF/HSP network in switchgrass Cd tolerance. Plant Cell Rep. 2018, 37, 1485–1497. [Google Scholar] [CrossRef]
- Cao, X.; Jia, J.B.; Li, H.; Li, M.C.; Luo, J.; Liang, Z.S.; Liu, T.X.; Liu, W.G.; Peng, C.H.; Luo, Z.B. Photosynthesis, water use efficiency and stable carbon isotope composition are associated with anatomical properties of leaf and xylem in six poplar species. Plant Biol. 2012, 14, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Bazihizina, N.; Colzi, I.; Giorni, E.; Mancuso, S.; Gonnelli, C. Photosynthesizing on metal excess: Copper differently induced changes in various photosynthetic parameters in copper tolerant and sensitive Silene paradoxa L. populations. Plant Sci. 2015, 232, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Perez-Romero, J.A.; Redondo-Gomez, S.; Mateos-Naranjo, E. Growth and photosynthetic limitation analysis of the Cd-accumulator Salicornia ramosissima under excessive cadmium concentrations and optimum salinity conditions. Plant Physiol. Biochem. 2016, 109, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Ying, R.R.; Jiang, D.; Zeng, X.W.; Morel, J.L.; Tang, Y.T.; Qiu, R.L. Impaired leaf CO2 diffusion mediates Cd-induced inhibition of photosynthesis in the Zn/Cd hyperaccumulator Picris divaricata. Plant Physiol. Biochem. 2013, 73, 70–76. [Google Scholar] [CrossRef]
- Flexas, J.; Baron, M.; Bota, J.; Ducruet, J.M.; Galle, A.; Galmes, J.; Jimenez, M.; Pou, A.; Ribas-Carbo, M.; Sajnani, C.; et al. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri x V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef] [Green Version]
- Sagardoy, R.; Vazquez, S.; Florez-Sarasa, I.D.; Albacete, A.; Ribas-Carbo, M.; Flexas, J.; Abadia, J.; Morales, F. Stomatal and mesophyll conductances to CO2 are the main limitations to photosynthesis in sugar beet (Beta vulgaris) plants grown with excess zinc. New Phytol. 2010, 187, 145–158. [Google Scholar] [CrossRef]
- Fahr, M.; Laplaze, L.; Bendaou, N.; Hocher, V.; Mzibri, M.E.; Bogusz, D.; Smouni, A. Effect of lead on root growth. Front. Plant Sci. 2013, 4, 175. [Google Scholar] [CrossRef] [Green Version]
- Kiniry, J.R.; Lynd, L.; Greene, N.; Johnson, M.V.V.; Casler, M.; Laser, M.S. Biofuels and water use: Comparison of maize and switchgrass and general perspectives. In New Research on Biofuels; Wright, J.H., Evans, D.A., Eds.; Nova Science Publishers: New York, NY, USA, 2008; pp. 17–30. [Google Scholar]
- Azoulay-Shemer, T.; Palomares, A.; Bagheri, A.; Israelsson-Nordstrom, M.; Engineer, C.B.; Bargmann, B.O.; Stephan, A.B.; Schroeder, J.I. Guard cell photosynthesis is critical for stomatal turgor production, yet does not directly mediate CO2-and ABA-induced stomatal closing. Plant J. 2015, 83, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Hills, A.; Batz, U.; Amtmann, A.; Lew, V.L.; Blatt, M.R. Systems dynamic modeling of the stomatal guard cell predicts emergent behaviors in transport, signaling, and volume control. Plant Physiol. 2012, 159, 1235–1251. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Pompeu, G.B.; Vilhena, M.B.; Gratao, P.L.; Carvalho, R.F.; Rossi, M.L.; Martinelli, A.P.; Azevedo, R.A. Abscisic acid-deficient sit tomato mutant responses to cadmium-induced stress. Protoplasma 2017, 254, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Wang, S.; Zhao, N.; Deng, S.; Zhao, C.; Li, N.; Sun, J.; Zhao, R.; Yi, H.; Shen, X.; et al. Exogenous Abscisic Acid Alleviates Cadmium Toxicity by Restricting Cd2+ Influx in Populus euphratica Cells. J. Plant Growth Regul. 2016, 35, 827–837. [Google Scholar] [CrossRef]
- Li, S.W.; Leng, Y.; Feng, L.; Zeng, X.Y. Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean (Vigna radiata L.) Wilczek seedlings under cadmium stress. Environ. Sci. Pollut. Res. 2014, 21, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Filardo, F.; Hu, X.; Zhao, X.; Fu, D. Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ. Sci. Pollut. Res. 2016, 23, 3758–3769. [Google Scholar] [CrossRef]
- Shpak, E.D.; McAbee, J.M.; Pillitteri, L.J.; Torii, K.U. Stomatal patterning and differentiation by synergistic interactions of receptor kinases. Science 2005, 309, 290–293. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, X.; Zhu, J.-K.; Dong, J. Demethylation of ERECTA receptor genes by IBM1 histone demethylase affects stomatal development. Development 2016, 143, 4452–4461. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.M.; Paciorek, T.; Abrash, E.; Bergmann, D.C. Modulators of stomatal lineage signal transduction alter membrane contact sites and reveal specialization among ERECTA kinases. Dev. Cell 2016, 38, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Zanten, M.; Snoek, L.B.; Proveniers, M.C.; Peeters, A.J. The many functions of ERECTA. Trends Plant Sci. 2009, 14, 214–218. [Google Scholar] [CrossRef]
- Torii, K.U.; Mitsukawa, N.; Oosumi, T.; Matsuura, Y.; Yokoyama, R.; Whittier, R.F.; Komeda, Y. The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell 1996, 8, 735–746. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Cultivar | Ecotype | No. | Cultivar | Ecotype |
|---|---|---|---|---|---|
| 1 | Alamo | Lowland | 8 | Forestburg | Upland |
| 2 | Bomaster | Lowland | 9 | Long Island | Upland |
| 3 | Kanlow | Lowland | 10 | New York | Upland |
| 4 | Blackwell | Upland | 11 | Shawnee | Upland |
| 5 | Carthage | Upland | 12 | Shelter | Upland |
| 6 | Cave-in-Rock | Upland | 13 | Sunburst | Upland |
| 7 | Dacotah | Upland | 14 | Trailblazer | Upland |
| Cultivar | RWCcontrol (%) | RWCCd (%) | RWC Reducing Rate (%) |
|---|---|---|---|
| Alamo | 90.06 ± 0.95 ab | 88.13 ± 0.33 ab | 1.91 ± 0.56 cd |
| Bomaster | 89.18 ± 0.62 abc | 87.40 ± 0.57 ab | 1.99 ± 0.47 cd |
| Kanlow | 90.23 ± 0.49 a | 88.88 ± 0.26 a | 1.71 ± 0.31 d |
| Blackwell | 87.11 ± 0.69 cd | 83.82 ± 0.70 cde | 3.76 ± 0.56 abc |
| Carthage | 84.63 ± 0.87 d | 82.58 ± 0.3 de | 2.39 ± 0.53 bcd |
| Cave-in-Rock | 90.7 ± 0.57 a | 86.97 ± 0.98 b | 4.10 ± 0.62 ab |
| Dacotah | 86.12 ± 1.07 d | 84.00 ± 0.58 cde | 2.85 ± 0.63 bcd |
| Forestburg | 86.38 ± 0.95 cd | 83.42 ± 0.35 cde | 3.41 ± 0.57 bcd |
| Long Island | 87.43 ± 1.25 bcd | 84.62 ± 0.20 c | 3.17 ± 0.70 bcd |
| New York | 87.23 ± 1.30 cd | 84.48 ± 0.33 cd | 3.51 ± 0.72 bcd |
| Shawnee | 90.54 ± 0.30 a | 88.77 ± 0.51 ab | 1.95 ± 0.33 cd |
| Shelter | 86.58 ± 0.81 cd | 83.08 ± 0.65 cde | 4.03 ± 0.59 ab |
| Sunburst | 87.08 ± 0.58 cd | 82.41 ± 0.92 e | 5.36 ± 0.62 a |
| Trailblazer | 87.32 ± 1.09 cd | 83.83 ± 0.82 cde | 3.97 ± 0.76 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Fang, Z.; Hu, B.; Wen, X.; Lou, L.; Cai, Q. Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation. Agronomy 2022, 12, 507. https://doi.org/10.3390/agronomy12020507
Hu Z, Fang Z, Hu B, Wen X, Lou L, Cai Q. Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation. Agronomy. 2022; 12(2):507. https://doi.org/10.3390/agronomy12020507
Chicago/Turabian StyleHu, Zhaoyang, Zhigang Fang, Bingyu Hu, Xuehui Wen, Laiqing Lou, and Qingsheng Cai. 2022. "Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation" Agronomy 12, no. 2: 507. https://doi.org/10.3390/agronomy12020507
APA StyleHu, Z., Fang, Z., Hu, B., Wen, X., Lou, L., & Cai, Q. (2022). Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation. Agronomy, 12(2), 507. https://doi.org/10.3390/agronomy12020507