
Impact of Olive Trees on the Microclimatic and Edaphic Environment of the Understorey Durum Wheat in an Alley Orchard of the Mediterranean Area

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Microclimate Parameters
2.4. Edaphic Parameters
2.5. Yield Components
2.6. Statistical Analysis
3. Results
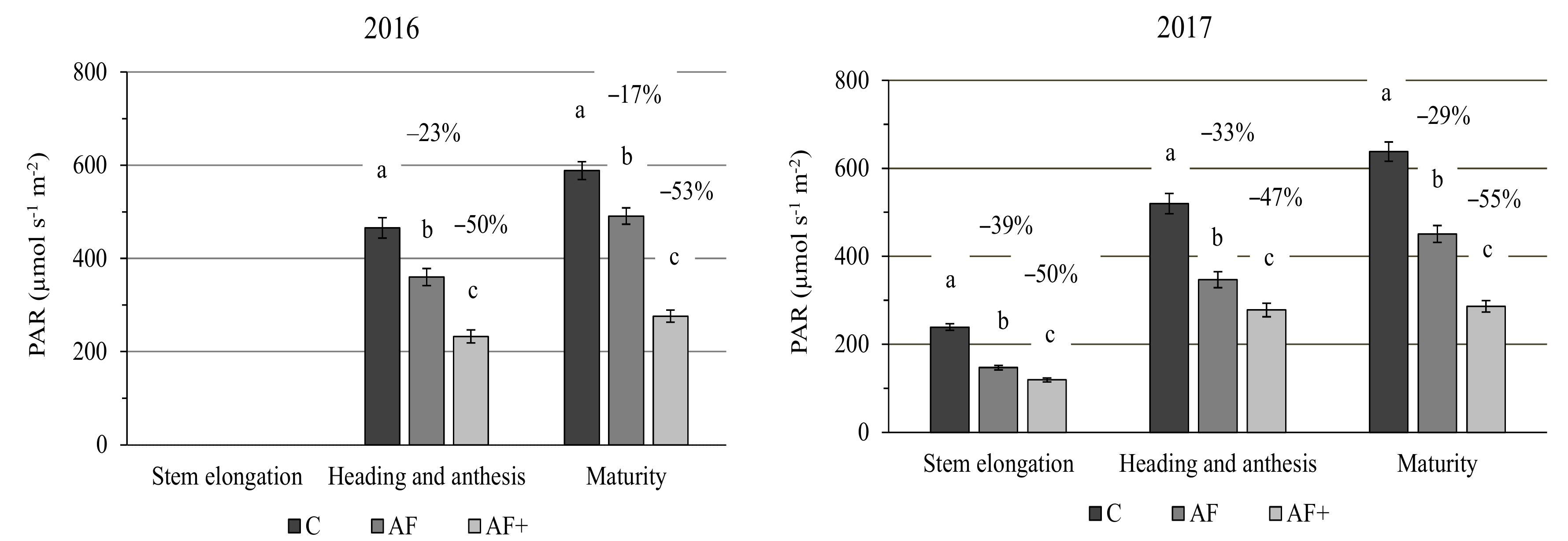
3.1. Solar Radiation
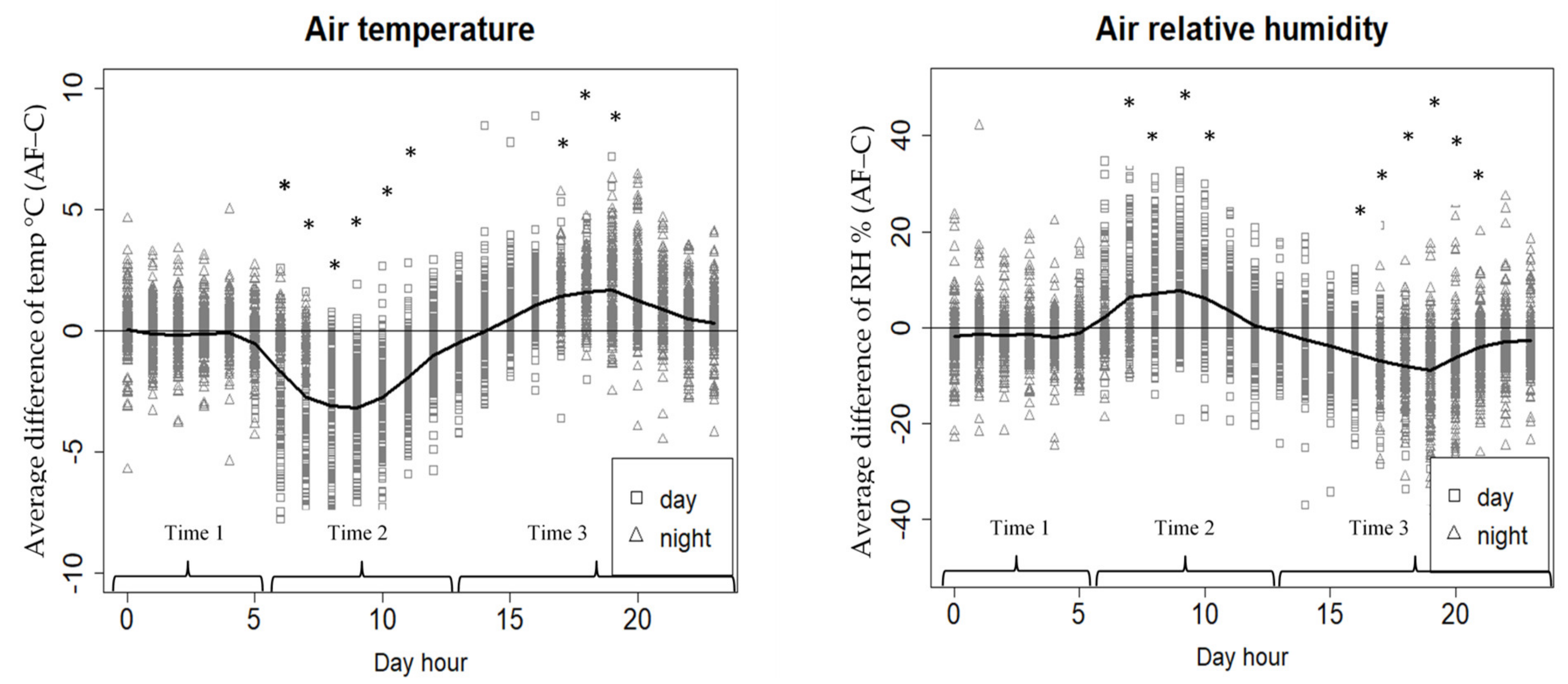
3.2. Air Temperature and Relative Humidity
3.3. Wind Speed
3.4. Edaphic Environment
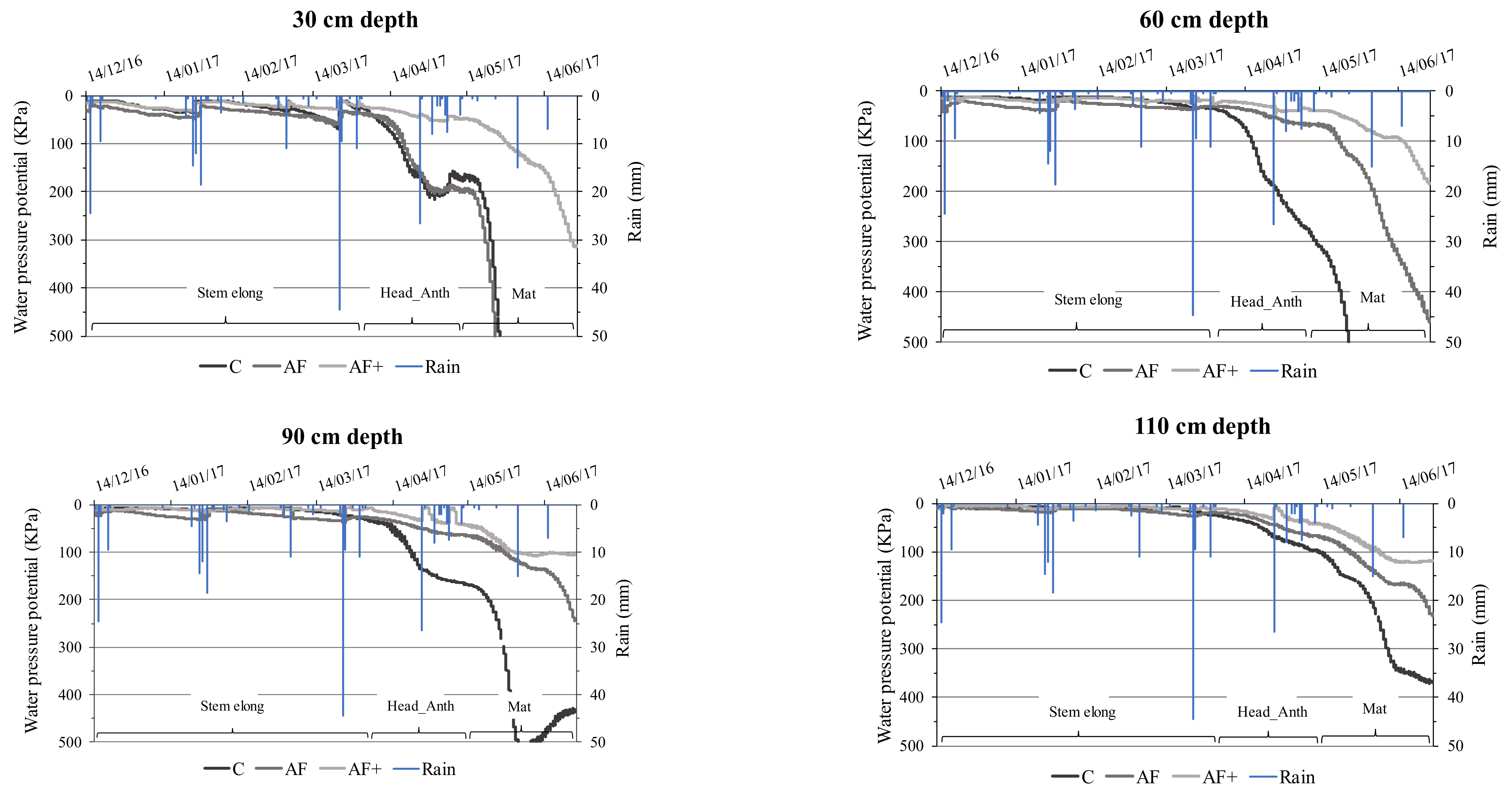
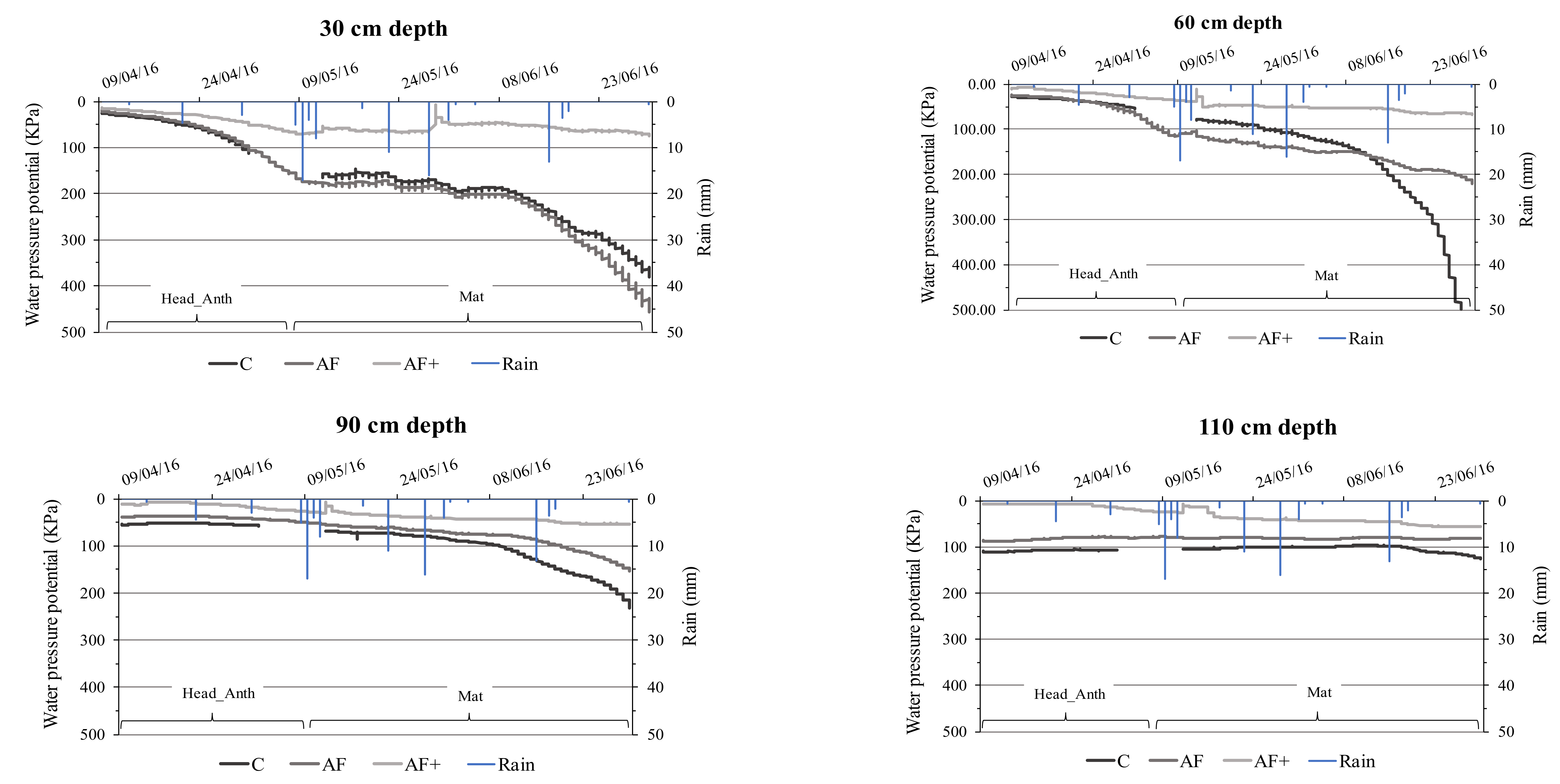
3.4.1. Soil Moisture Content
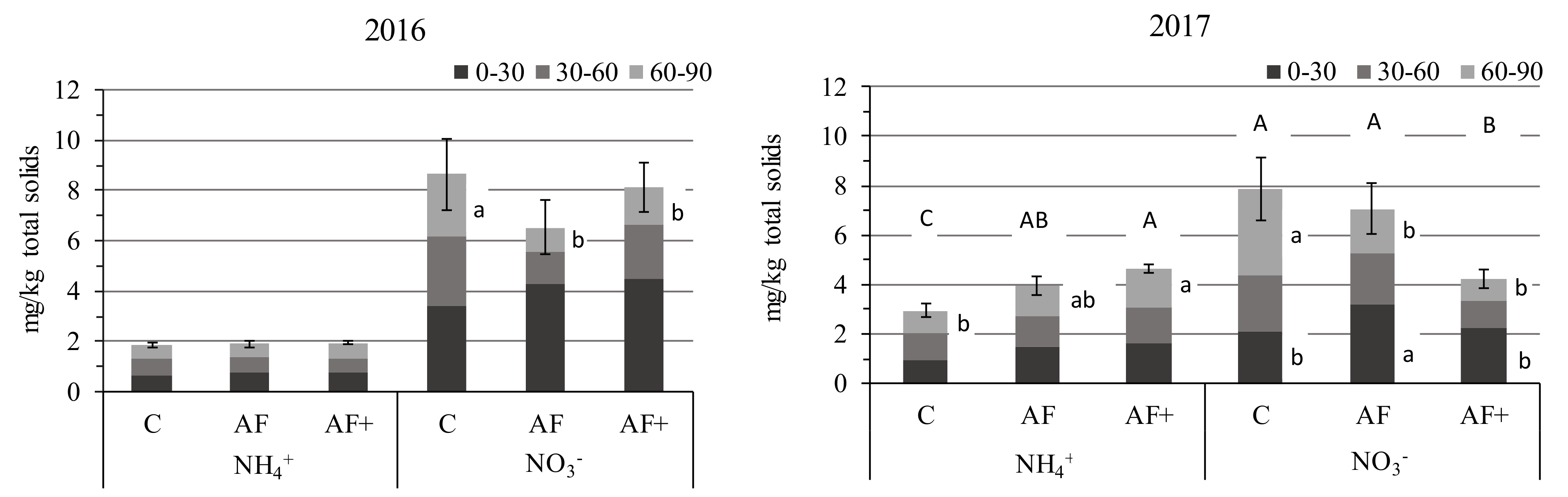
3.4.2. Soil Fertility
3.5. Mycorrhizal Colonization
3.6. Durum Wheat Yield and Yield Components
4. Discussion
4.1. Air Temperature and Relative Humidity
4.2. Wind Speed
4.3. Soil Moisture
4.4. Soil Fertility
4.5. Arbuscular Mycorrhizal Fungi (AMF)
4.6. Durum Wheat Performances
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nerlich, K.; Graeff-Hönninger, S.; Claupein, W. Agroforestry in Europe: A review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agrofor. Syst. 2013, 87, 475–492. [Google Scholar] [CrossRef]
- Nieto, O.M.; Castro, J.; Fernández-Ondoño, E. Conventional tillage versus cover crops in relation to carbon fixation in Mediterranean olive cultivation. Plant Soil. 2013, 365, 321–335. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Xoplaki, E. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Carbone, G.J.; Grego, J.M. Uncertainty and hotspots in 21st century projections of agricultural drought from CMIP5 models. Sci. Rep. 2019, 9, 4922. [Google Scholar] [CrossRef] [PubMed]
- Philandras, C.M.; Nastos, P.T.; Kapsomenakis, J.; Douvis, K.C.; Tselioudis, G.; Zerefos, C.S. Long term precipitation trends and variability within the Mediterranean region. Nat. Hazards Earth Syst. Sci. 2011, 11, 3235–3250. [Google Scholar] [CrossRef] [Green Version]
- Bilbao, J.; Román, R.; De Miguel, A. Temporal and Spatial Variability in Surface Air Temperature and Diurnal Temperature Range in Spain over the Period 1950–2011. Climate 2019, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Georgopoulou, E. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Arenas-Corraliza, M.G.; López-Díaz, M.L.; Moreno, G. Winter cereal production in a Mediterranean silvoarable walnut system in the face of climate change. Agric. Ecosyst. Environ. 2018, 264, 111–118. [Google Scholar] [CrossRef]
- Ribes, A.; Thao, S.; Vautard, R.; Dubuisson, B.; Somot, S.; Colin, J.; Soubeyroux, J.M. Observed increase in extreme daily rainfall in the French Mediterranean. Clim. Dyn. 2019, 52, 1095–1114. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Nguyen, Q.; Hoang, M.H.; Öborn, I.; van Noordwijk, M. Multipurpose agroforestry as a climate change resiliency option for farmers: An example of local adaptation in Vietnam. Clim. Chang. 2013, 117, 241–257. [Google Scholar] [CrossRef]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautasso, M.; Döring, T.F.; Garbelotto, M.; Pellis, L.; Jeger, M.J. Impacts of climate change on plant diseases, opinions and trends. Eur. J. Plant Pathol. 2012, 133, 295–313. [Google Scholar] [CrossRef] [Green Version]
- Inurreta-Aguirre, H.D.; Lauri, P.É.; Dupraz, C.; Gosme, M. Yield components and phenology of durum wheat in a Mediterranean alley-cropping system. Agrofor. Syst. 2018, 1–14. [Google Scholar] [CrossRef]
- Moriondo, M.; Bindi, M. Impact of climate change on the phenology of typical Mediterranean crops. Ital. J. Agrometeorol. 2007, 3, 5–12. [Google Scholar]
- Ren, S.; Qin, Q.; Ren, H. Contrasting wheat phenological responses to climate change in global scale. Sci. Total Environ. 2019, 665, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Gouache, D.; Le Bris, X.; Bogard, M.; Deudon, O.; Pagé, C.; Gate, P. Evaluating agronomic adaptation options to increasing heat stress under climate change during wheat grain filling in France. Eur. J. Agron. 2012, 39, 62–70. [Google Scholar] [CrossRef]
- Xiao, D.; Bai, H.; Liu, D. Impact of future climate change on wheat production: A simulated case for China’s wheat system. Sustainability 2018, 10, 1277. [Google Scholar] [CrossRef] [Green Version]
- Torralba, M.; Fagerholm, N.; Burgess, P.J.; Moreno, G.; Plieninger, T. Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric. Ecosyst. Environ. 2016, 230, 150–161. [Google Scholar] [CrossRef] [Green Version]
- McAdam, J.H.; Burgess, P.J.; Graves, A.R.; Rigueiro-Rodríguez, A.; Mosquera-Losada, M.R. Classifications and functions of agroforestry systems in Europe. In Agroforestry in Europe; Springer: Dordrecht, The Netherlands, 2009; pp. 21–41. [Google Scholar] [CrossRef]
- Bisht, N.; Sah, V.K.; Satyawali, K.; Tewari, S.; Kandpal, G. Assessment of soil quality and wheat yield under open and poplar based farming system in Tarai region of Uttarakhand. Indian J. Agric. Res. 2018, 52, 101–103. [Google Scholar] [CrossRef]
- Kay, S.; Crous-Duran, J.; Ferreiro-Domínguez, N.; De Jalón, S.G.; Graves, A.; Moreno, G.; Szerencsits, E. Spatial similarities between European agroforestry systems and ecosystem services at the landscape scale. Agrofor. Syst. 2018, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.K.; Park, H.M. Effects of Windbreak Planting on Crop Productivity for Agroforestry Practices in a Semi-Arid Region. J. For. Environ. Sci. 2017, 33, 348–354. [Google Scholar] [CrossRef]
- Schroth, G.; Sinclair, F.L. (Eds.) Trees, Crops, and Soil Fertility: Concepts and Research Methods; Cabi: Wallingford, UK, 2003. [Google Scholar]
- Tscharntke, T.; Clough, Y.; Bhagwat, S.A.; Buchori, D.; Faust, H.; Hertel, D.; Scherber, C. Multifunctional shade-tree management in tropical agroforestry landscapes—A review. J. Appl. Ecol. 2011, 48, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Mbow, C.; Smith, P.; Skole, D.; Duguma, L.; Bustamante, M. Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr. Opin. Environ. Sustain. 2014, 6, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Pinho, R.C.; Miller, R.P.; Alfaia, S.S. Agroforestry and the improvement of soil fertility: A view from Amazonia. Appl. Environ. Soil Sci. 2012, 1–11. [Google Scholar] [CrossRef]
- Jezeer, R.E.; Santos, M.J.; Boot, R.G.; Junginger, M.; Verweij, P.A. Effects of shade and input management on economic performance of small-scale Peruvian coffee systems. Agric. Sys. 2018, 162, 179–190. [Google Scholar] [CrossRef]
- Santos, P.Z.F.; Crouzeilles, R.; Sansevero, J.B.B. Can agroforestry systems enhance biodiversity and ecosystem service provision in agricultural landscapes? A meta-analysis for the Brazilian Atlantic Forest. Forest. Ecol. Manag. 2019, 433, 140–145. [Google Scholar] [CrossRef]
- Mbow, C.; Van Noordwijk, M.; Luedeling, E.; Neufeldt, H.; Minang, P.A.; Kowero, G. Agroforestry solutions to address food security and climate change challenges in Africa. Curr. Opin. Environ. Sustai. 2014, 6, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Sida, T.S.; Baudron, F.; Kim, H.; Giller, K.E. Climate-smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. For. Meteorol. 2018, 248, 339–347. [Google Scholar] [CrossRef]
- Kuyah, S.; Whitney, C.W.; Jonsson, M.; Sileshi, G.W.; Öborn, I.; Muthuri, C.W.; Luedeling, E. Agroforestry delivers a win-win solution for ecosystem services in sub-Saharan Africa. A meta-analysis. Agron. Sustain. Dev. 2019, 39, 47. [Google Scholar] [CrossRef] [Green Version]
- Daoui, K.; Fatemi, Z.E.A. Agroforestry systems in Morocco: The case of olive tree and annual crops association in Sais region. In Science, Policy and Politics of Modern Agricultural System; Springer: Dordrecht, The Netherlands, 2014; pp. 281–289. [Google Scholar]
- Rao, K.P.C.; Verchot, L.V.; Laarman, J. Adaptation to climate change through sustainable management and development of agroforestry systems. J. SAT Agric. Res. 2007, 4, 1–30. [Google Scholar]
- Francis, G.; Oliver, J.; Mulpuri, S. High yielding and trait specific genotypes and genetic associations among yield and yield contributing traits in Jatropha curcas L. Agrofor. Syst. 2018, 92, 1417–1436. [Google Scholar] [CrossRef]
- Valencia, V.; García-Barrios, L.; Sterling, E.J.; West, P.; Meza-Jiménez, A.; Naeem, S. Smallholder response to environmental change: Impacts of coffee leaf rust in a forest frontier in Mexico. Land Use Policy 2018, 79, 463–474. [Google Scholar] [CrossRef]
- Ong, C.K.; Black, C.; Wilson, J. (Eds.) Tree-Crop Interactions: Agroforestry in a Changing Climate; CABI: Wallingford, UK, 2015. [Google Scholar]
- Thomas, S.C.; Halim, M.A.; Gale, N.V.; Sujeeun, L. Biochar enhancement of facilitation effects in agroforestry: Early growth and physiological responses in a maize-leucaena model system. Agrofor. Syst. 2018, 1–13. [Google Scholar] [CrossRef]
- Youkhana, A.H.; Idol, T.W. Leucaena-KX2 mulch additions increase growth, yield and soil C and N in a managed full-sun coffee system in Hawaii. Agrofor. Syst. 2016, 90, 325–337. [Google Scholar] [CrossRef]
- Mokgolodi, N.C.; Setshogo, M.P.; Shi, L.L.; Liu, Y.J.; Ma, C. Achieving food and nutritional security through agroforestry: A case of Faidherbia albida in sub-Saharan Africa. For. Stud. China 2011, 13, 123–131. [Google Scholar] [CrossRef]
- Sileshi, G.W. The magnitude and spatial extent of influence of Faidherbia albida trees on soil properties and primary productivity in drylands. J. Arid Environ. 2016, 132, 1–14. [Google Scholar] [CrossRef]
- Suárez, L.R.; Salazar, J.C.S.; Casanoves, F.; Bieng, M.A.N. Cacao agroforestry systems improve soil fertility: Comparison of soil properties between forest, cacao agroforestry systems, and pasture in the Colombian Amazon. Agric. Ecosyst. Environ. 2021, 314, 107349. [Google Scholar] [CrossRef]
- Deheuvels, O.; Avelino, J.; Somarriba, E.; Malezieux, E. Vegetation structure and productivity in cocoa-based agroforestry systems in Talamanca, Costa Rica. Agric. Ecosyst. Environ. 2012, 149, 181–188. [Google Scholar] [CrossRef]
- Deheuvels, O.; Rousseau, G.X.; Quiroga, G.S.; Franco, M.D.; Cerda, R.; Mendoza, S.J.V.; Somarriba, E. Biodiversity is affected by changes in management intensity of cocoa-based agroforests. Agrofor. Syst. 2014, 88, 1081–1099. [Google Scholar] [CrossRef]
- Lin, B.B. The role of agroforestry in reducing water loss through soil evaporation and crop transpiration in coffee agroecosystems. Agric. For. Meteorol. 2010, 150, 510–518. [Google Scholar] [CrossRef]
- Fagerholm, N.; Torralba, M.; Burgess, P.J.; Plieninger, T. A systematic map of ecosystem services assessments around European agroforestry. Ecol. Indic. 2016, 62, 47–65. [Google Scholar] [CrossRef]
- Panozzo, A.; Bernazeau, B.; Desclaux, D. Durum wheat in organic olive orchard: Good deal for the farmers? Agrofor. Syst. 2019, 1–11. [Google Scholar] [CrossRef]
- Panozzo, A.; Huang, H.; Bernazeau, B.; Vamerali, T.; Samson, M.F.; Desclaux, D. Morphology, phenology, yield, and quality of durum wheat cultivated within organic olive orchards of the Mediterranean area. Agronomy 2020, 10, 1789. [Google Scholar] [CrossRef]
- Delteil, A.; Gobbato, E.; Cayrol, B.; Estevan, J.; Michel-Romiti, C.; Dievart, A. Several wall-associated kinases participate positively and negatively in basal defense against rice blast fungus. BMC Plant Biol. 2016, 16, 17. [Google Scholar] [CrossRef]
- Mycocalc Software; Institut National de la Recherche Agronomique. Available online: https://www2.dijon.inra.fr/mychintec/Mycocalc-prg/download.html (accessed on 30 January 2022).
- Huler, S. Defining the Wind: The Beaufort Scale and How a 19th-Century Admiral Turned Science into Poetry; Crown: New York, NY, USA, 2007. [Google Scholar]
- Lin, B.B. Agroforestry management as an adaptive strategy against potential microclimate extremes in coffee agriculture. Agric. For. Meteorol. 2007, 144, 85–94. [Google Scholar] [CrossRef]
- Gosme, M.; Inurreta-Aguirre, H.D.; Dupraz, C. Microclimatic effect of agroforestry on diurnal temperature cycle. In Proceedings of the 3rd European Agroforestry Conference, European Agroforestry Federation, Montpellier, France, 23–25 May 2016; pp. 183–186. [Google Scholar]
- Kanzler, M.; Böhm, C.; Mirck, J.; Schmitt, D.; Veste, M. Microclimate effects on evaporation and winter wheat (Triticum aestivum L.) yield within a temperate agroforestry system. Agrofor. Syst 2019, 93, 1821–1841. [Google Scholar] [CrossRef]
- García, G.A.; Dreccer, M.F.; Miralles, D.J.; Serrago, R.A. High night temperatures during grain number determination reduce wheat and barley grain yield: A field study. Glob. Chang. Boil. 2015, 21, 4153–4164. [Google Scholar] [CrossRef]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [Green Version]
- Bos, H.J.; Neuteboom, J.H. Morphological analysis of leaf and tiller number dynamics of wheat (Triticum aestivum L.): Responses to temperature and light intensity. Ann. Bot. 1998, 81, 131–139. [Google Scholar] [CrossRef]
- Thorne, G.N.; Wood, D.W. Effects of radiation and temperature on tiller survival, grain number and grain yield in winter wheat. Ann. Bot. 1987, 59, 413–426. [Google Scholar] [CrossRef]
- Lin, B.B. Coffee (Cafe arabica var. Bourbon) fruit growth and development under varying shade levels in the Soconusco Region of Chiapas, Mexico. J. Sustain. Agric. 2009, 33, 51–65. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar] [CrossRef]
- Smith, J.; Pearce, B.D.; Wolfe, M.S. Reconciling productivity with protection of the environment: Is temperate agroforestry the answer? Renew. Agric. Food Syst. 2013, 28, 80–92. [Google Scholar] [CrossRef]
- Ong, W.Y.; Mackie, K. A light and electron microscopic study of the CB1 cannabinoid receptor in the primate spinal cord. J. Neurocytol. 1999, 28, 39–45. [Google Scholar] [CrossRef]
- Schoeneberger, M.; Bentrup, G.; De Gooijer, H.; Soolanayakanahally, R.; Sauer, T.; Brandle, J. Branching out: Agroforestry as a climate change mitigation and adaptation tool for agriculture. J. Soil Water Conserv. 2012, 67, 128A–136A. [Google Scholar] [CrossRef] [Green Version]
- Filella, I.; Peñuelas, J. Partitioning of water and nitrogen in co-occurring Mediterranean woody shrub species of different evolutionary history. Oecologia 2003, 137, 51–61. [Google Scholar] [CrossRef]
- Eliades, M.; Bruggeman, A.; Djuma, H.; Lubczynski, M. Tree water dynamics in a semi-arid Pinus brutia forest. Water 2018, 10, 1039. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.J.; Meng, P.; Zhang, J.S.; Wan, X. Hydraulic lift by Juglans regia relates to nutrient status in the intercropped shallow-root crop plant. Plant Soil 2014, 374, 629–641. [Google Scholar] [CrossRef]
- Gillespie, A.R.; Jose, S.; Mengel, D.B.; Hoover, W.L.; Pope, P.E.; Seifert, J.R. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 1. Production physiology. Agrofor. Syst. 2000, 48, 25–40. [Google Scholar] [CrossRef]
- Wanvestraut, R.H.; Jose, S.; Nair, P.R.; Brecke, B.J. Competition for water in a pecan (Carya illinoensis K. Koch)–cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Agrofor. Syst. 2004, 60, 167–179. [Google Scholar] [CrossRef]
- Miller, A.W.; Pallardy, S.G. Resource competition across the crop-tree interface in a maize-silver maple temperate alley cropping stand in Missouri. Agrofor. Syst. 2001, 53, 247–259. [Google Scholar] [CrossRef]
- Dal Cortivo, C.; Barion, G.; Ferrari, M.; Visioli, G.; Dramis, L.; Panozzo, A.; Vamerali, T. Effects of field inoculation with VAM and bacteria consortia on root growth and nutrients uptake in common wheat. Sustainability 2018, 10, 3286. [Google Scholar] [CrossRef] [Green Version]
- Zake, J.; Pietsch, S.A.; Friedel, J.K.; Zechmeister-Boltenstern, S. Can agroforestry improve soil fertility and carbon storage in smallholder banana farming systems? J. Plant Nutr. Soil Sci. 2015, 178, 237–249. [Google Scholar] [CrossRef]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signalling in plants. J. Exp. Bot. 2017, 68, 2501–2512. [Google Scholar] [CrossRef]
- Peichl, M.; Thevathasan, N.V.; Gordon, A.M.; Huss, J.; Abohassan, R.A. Carbon sequestration potentials in temperate tree-based intercropping systems, southern Ontario, Canada. Agrofor. Syst. 2006, 66, 243–257. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Seifert, J.R.; Mengel, D.B.; Pope, P.E. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 3. Competition for nitrogen and litter decomposition dynamics. Agrofor. Syst. 2000, 48, 61–77. [Google Scholar] [CrossRef]
- Benbi, D.K.; Khosa, M.K. Effects of temperature, moisture, and chemical composition of organic substrates on C mineralization in soils. Commun. Soil Sci. Plant Anal. 2014, 45, 2734–2753. [Google Scholar] [CrossRef]
- Querné, A.; Battie-laclau, P.; Dufour, L.; Wery, J.; Dupraz, C. Effects of walnut trees on biological nitrogen fixation and yield of intercropped alfalfa in a Mediterranean agroforestry system. Eur. J. Agron. 2017, 84, 35–46. [Google Scholar] [CrossRef]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Laurindo, L.K.; de Souza, T.A.F.; da Silva, L.J.R.; Casal, T.B.; de Jesus Conceição Pires, K.; Kormann, S.; Siminski, A. Arbuscular mycorrhizal fungal community assembly in agroforestry systems from the Southern Brazil. Biologia 2021, 76, 1099–1107. [Google Scholar] [CrossRef]
- da Silva Maia, R.; Vasconcelos, S.S.; Viana-Junior, A.B.; Castellani, D.C.; Kato, O.R. Oil palm (Elaeis guineensis) shows higher mycorrhizal colonization when planted in agroforestry than in monoculture. Agrofor. Syst. 2021, 95, 731–740. [Google Scholar] [CrossRef]
- Bainard, L.D.; Klironomos, J.N.; Gordon, A.M. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia 2011, 54, 57–61. [Google Scholar] [CrossRef]
- De Carvalho, A.M.X.; de Castro Tavares., R.; Cardoso, I.M.; Kuyper, T.W. Mycorrhizal Associations in Agroforestry Systems. In Soil Biology and Agriculture in the Tropics; Dion, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 21. [Google Scholar] [CrossRef]
- Shukla, A.; Kumar, A.; Chaturvedi, O.P.; Nagori, T.; Kumar, N.; Gupta, A. Efficacy of rhizobial and phosphate-solubilizing bacteria and arbuscular mycorrhizal fungi to ameliorate shade response on six pulse crops. Agrofor. Syst. 2018, 92, 499–509. [Google Scholar] [CrossRef]
- Dobo, B.; Asefa, F.; Asfaw, Z. Effect of tree-enset-coffee based agro-forestry practices on arbuscular mycorrhizal fungi (AMF) species diversity and spore density. Agrofor. Syst. 2018, 92, 525–540. [Google Scholar] [CrossRef]
- Isaac, M.E.; Borden, K.A. Nutrient acquisition strategies in agroforestry systems. Plant Soil 2019, 444, 1–19. [Google Scholar] [CrossRef]
- Chifflot, V.; Rivest, D.; Olivier, A.; Cogliastro, A.; Khasa, D. Molecular analysis of arbuscular mycorrhizal community structure and spores distribution in tree-based intercropping and forest systems. Agric. Ecosyst. Environ. 2009, 131, 32–39. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Bergmann, J.; Verbruggen, E.; Veresoglou, S.D.; Lehmann, A. Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phytol. 2015, 205, 1385–1388. [Google Scholar] [CrossRef]
- Li, F.; Meng, P.; Fu, D.; Wang, B. Light distribution, photosynthetic rate and yield in a Paulownia-wheat intercropping system in China. Agrofor. Syst. 2008, 74, 163–172. [Google Scholar] [CrossRef]
- Dufour, L.; Metay, A.; Talbot, G.; Dupraz, C. Assessing light competition for cereal production in temperate agroforestry systems using experimentation and crop modelling. J. Agron. Crop Sci. 2013, 199, 217–227. [Google Scholar] [CrossRef]
- Yang, T.; Duan, Z.P.; Zhu, Y.; Gan, Y.W.; Wang, B.J.; Hao, X.D.; Li, L.H. Effects of distance from a tree line on photosynthetic characteristics and yield of wheat in a jujube tree/wheat agroforestry system. Agrofor. Syst. 2019, 93, 1545–1555. [Google Scholar] [CrossRef]
- Li, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Effects of shading on morphology, physiology and grain yield of winter wheat. Eur. J. Agron. 2010, 33, 267–275. [Google Scholar] [CrossRef]
- Artru, S.; Garré, S.; Dupraz, C.; Hiel, M.P.; Blitz-Frayret, C.; Lassois, L. Impact of spatio-temporal shade dynamics on wheat growth and yield, perspectives for temperate agroforestry. Eur. J. Agron. 2017, 82, 60–70. [Google Scholar] [CrossRef]
- Dupraz, C.; Blitz-Frayret, C.; Lecomte, I.; Molto, Q.; Reyes, F.; Gosme, M. Influence of latitude on the light availability for intercrops in an agroforestry alley-cropping system. Agrofor. Syst. 2018, 92, 1019–1033. [Google Scholar] [CrossRef]
- Yang, L.; Ding, X.; Liu, X.; Li, P.; Eneji, A.E. Impacts of long-term jujube tree/winter wheat–summer maize intercropping on soil fertility and economic efficiency—A case study in the lower North China Plain. Eur. J. Agron. 2016, 75, 105–117. [Google Scholar] [CrossRef]
- Carrier, M.; Gonzalez, F.A.R.; Cogliastro, A.; Olivier, A.; Vanasse, A.; Rivest, D. Light availability, weed cover and crop yields in second generation of temperate tree-based intercropping systems. Field Crops Res. 2019, 239, 30–37. [Google Scholar] [CrossRef]
- Pardon, P.; Reubens, B.; Mertens, J.; Verheyen, K.; De Frenne, P.; De Smet, G.; Reheul, D. Effects of temperate agroforestry on yield and quality of different arable intercrops. Agric. Syst. 2018, 166, 135–151. [Google Scholar] [CrossRef]
- Desclaux, D. Participatory Ecobreeding for Agroecology. In Proceedings of the “Innovative Research for Organic 3.0” 19th Organic World Congress, New Delhi, India, 9–11 November 2017; Volume 2, p. 654. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycle Period | Wind Speed | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Means | Hourly Mean >4.7 m s−1 | Hourly Max >4.7 m s−1 | |||||||
| m s−1 | n. of Hours | n. of Hours | |||||||
| C | AF | AF+ | C | AF | AF+ | C | AF | AF+ | |
| Stem elongation | 2.17 a | 0.37 b | 0.01 c | 277 a | 11 b | 0 c | 1083 a | 142 b | 0 c |
| Heading and anthesis | 2.03 a | 0.25 b | 0.01 c | 37 a | 0 b | 0 b | 327 a | 13 b | 0 b |
| Maturity | 1.60 a | 0.12 b | 0.01 c | 9 a | 0 b | 0 b | 347 a | 6 b | 0 c |
| Average | 2.02 a | 0.29 b | 0.01 c | 323 a | 11 b | 0 c | 1757 a | 161 b | 0 c |
| Treatment | Frequency | Intensity | Arbuscular Abundance | |||
|---|---|---|---|---|---|---|
| Mean (%) | %var./C | Mean (%) | %var./C | Mean (%) | %var./C | |
| C | 96.4 b | 43.8 b | 17.2 b | |||
| AF | 100.0 a | +4% | 66.0 a | +51% | 30.0 a | +74% |
| AF+ | 99.8 a | +4% | 61.7 a | +41% | 28.6 a | +66% |
| Treatment | Yield | Tillers/Plant | Spikes/Plant | Grains/Spike | TGW | One Plant Biomass | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t ha−1 | % var./C | n. | % var./C | n. | % var./C | n. | % var./C | g | % var./C | g DM | % var./C | |
| C | 1.6 a | 2.4 a | 2.1 a | 27.2 a | 52.0 ab | 119.9 a | ||||||
| AF | 0.8 ab | –50% | 1.8 ab | –26% | 1.5 a | –27% | 4.3 b | –84% | 58.1 a | +12% | 31.1 b | –74% |
| AF+ | 0.4 b | –76% | 1.4 b | –39% | 1.1 a | –49% | 4.6 b | –83% | 36.6 b | –30% | 23.6 b | –80% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panozzo, A.; Huang, H.-Y.; Bernazeau, B.; Meunier, F.; Turc, O.; Duponnois, R.; Prin, Y.; Vamerali, T.; Desclaux, D. Impact of Olive Trees on the Microclimatic and Edaphic Environment of the Understorey Durum Wheat in an Alley Orchard of the Mediterranean Area. Agronomy 2022, 12, 527. https://doi.org/10.3390/agronomy12020527
Panozzo A, Huang H-Y, Bernazeau B, Meunier F, Turc O, Duponnois R, Prin Y, Vamerali T, Desclaux D. Impact of Olive Trees on the Microclimatic and Edaphic Environment of the Understorey Durum Wheat in an Alley Orchard of the Mediterranean Area. Agronomy. 2022; 12(2):527. https://doi.org/10.3390/agronomy12020527
Chicago/Turabian StylePanozzo, Anna, Hsin-Ya Huang, Bruno Bernazeau, Florence Meunier, Olivier Turc, Robin Duponnois, Yves Prin, Teofilo Vamerali, and Dominique Desclaux. 2022. "Impact of Olive Trees on the Microclimatic and Edaphic Environment of the Understorey Durum Wheat in an Alley Orchard of the Mediterranean Area" Agronomy 12, no. 2: 527. https://doi.org/10.3390/agronomy12020527
APA StylePanozzo, A., Huang, H. -Y., Bernazeau, B., Meunier, F., Turc, O., Duponnois, R., Prin, Y., Vamerali, T., & Desclaux, D. (2022). Impact of Olive Trees on the Microclimatic and Edaphic Environment of the Understorey Durum Wheat in an Alley Orchard of the Mediterranean Area. Agronomy, 12(2), 527. https://doi.org/10.3390/agronomy12020527