
Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
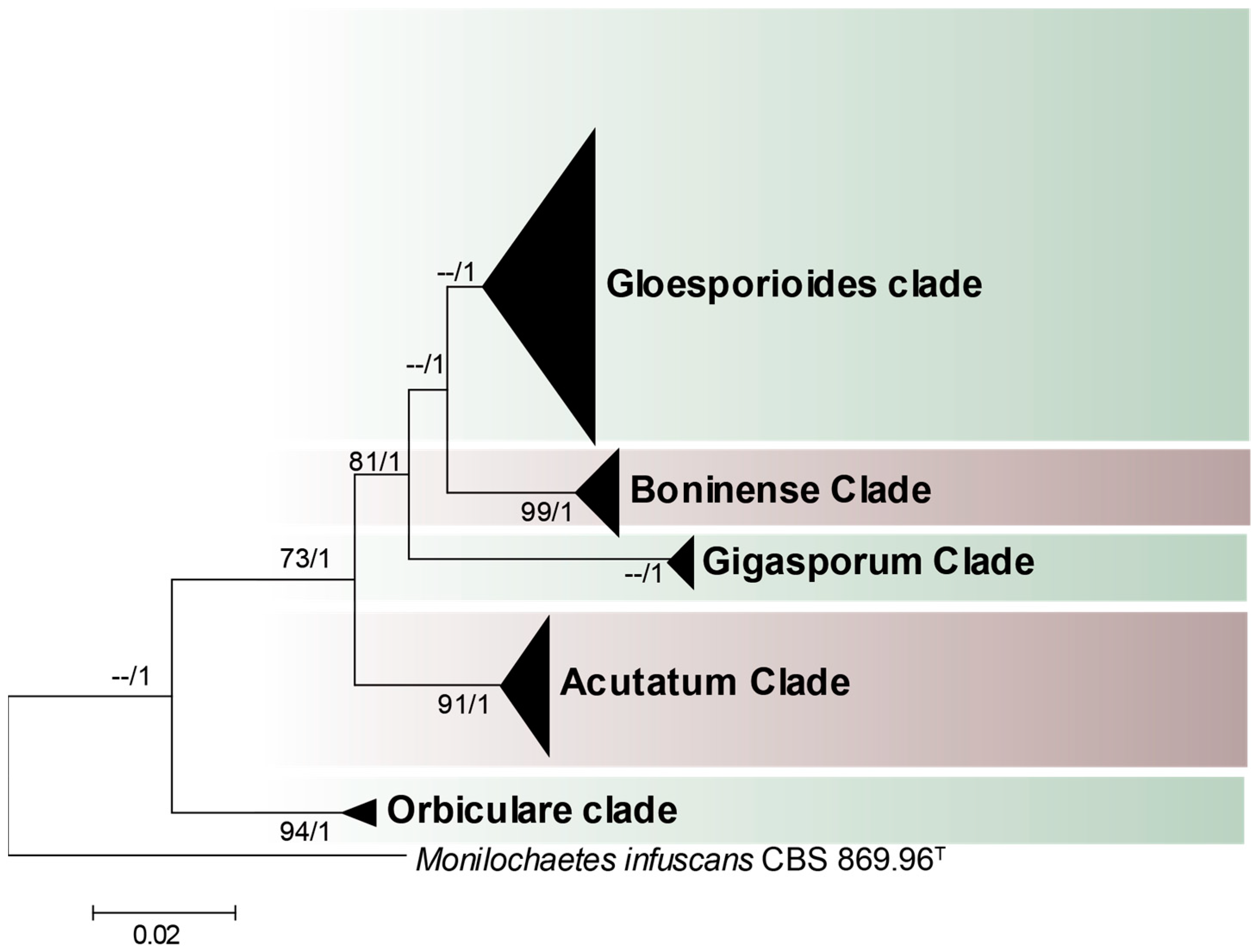
3.1. Species Complexes of Colletotrichum Associated with Crops in Northern South America
3.2. Geographical Distribution of Colletotrichum Complexes in Northern South America
3.3. Phytopathological Aspects
3.3.1. Hosts, Diversity and Botanical Families
3.3.2. Virulence and Aggressiveness
3.3.3. Host Specificity
3.3.4. Strategies for Controlling Colletotrichum Species in Northern South America
3.4. Colletotrichum Species as Endophytes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sutton, B.C. The Genus Glomerella and Its Anamorph Colletotrichum, 1st ed.; CAB International: Wallingford, UK, 1992; pp. 1–26. [Google Scholar]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. ámagnum and C. áorchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cai, L.; Crous, P.W.; Damm, U. The Colletotrichum gigasporum species complex. Persoonia 2014, 33, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum—Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corda, A.C.I. Die pilze Deutschlands. In Deutschlands Flora in Abbildungen nach der Natur mit Beschreibungen, 3rd ed.; Sturm, J., Ed.; Tables 21–32; Sturm: Nürnberg, Germany, 1831; Volume 12, pp. 33–64. [Google Scholar]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhabg, W.; Zhao, W.S.; Yan, J.Y. Notes on currently accepted species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: Geographic distribution, host range and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Damm, U.; Cai, L.; Crous, P.W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Divers. 2013, 61, 89–105. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Nilsson, R.H.; Alias, S.A.; Ariyawansa, H.A.; Blair, J.E.; Cai, L.; de Cock, A.W.A.M.; Dissanayake, J.; Glockling, S.L.; Goonasekara, I.D.; et al. One stop shop: Backbones trees for important phytopathogenic genera: I (2014). Fungal Divers. 2014, 67, 21–125. [Google Scholar] [CrossRef] [Green Version]
- Doyle, V.P.; Oudemans, P.V.; Rehner, S.A.; Litt, A. Habitat and host indicate lineage identity in Colletotrichum gloeosporioides s.l. from wild and agricultural landscapes in North America. PLoS ONE 2013, 8, e62394. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.; Azinheira, H.G.; Talhinhas, P.; Batista, D.; Ramos, A.P.; Silva, M.D.C.; Oliveira, H.; Várzea, V. Pathological, morphological, cytogenomic, biochemical and molecular data support the distinction between Colletotrichum cigarro comb. et stat. nov. and Colletotrichum kahawae. Plants 2020, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- Moreno–Moran, M.; Burbano–Figueroa, O. First report of Colletotrichum gossypii var. cephalosporioides causing cotton ramulosis in Colombia. Plant Dis. 2016, 100, 653. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2018, 42, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [Green Version]
- Afanador–Kafuri, L.; Minz, D.; Maymon, M.; Freeman, S. Characterization of Colletotrichum isolates from tamarillo, Passiflora, and mango in Colombia and identification of a unique species from the genus. Phytopathology 2003, 93, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, E.; Gañán, L.; Rojas–Triviño, A.; Mejía, J.F.; Llano, G.A.; González, A. Diversity and pathogenicity of Colletotrichum species isolated from soursop in Colombia. Eur. J. Plant Pathol. 2014, 139, 325–338. [Google Scholar] [CrossRef]
- Afanador–Kafuri, L.; González, A.; Gañán, L.; Mejía, J.F.; Cardona, N.; Alvarez, E. Characterization of the Colletotrichum species causing anthracnose in Andean blackberry in Colombia. Plant Dis. 2014, 98, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Cannon, P.F.; Liu, F.; Barreto, R.W.; Guatimosim, E.; Crous, P.W. The Colletotrichum orbiculare species complex: Important pathogens of field crops and weeds. Fungal Divers. 2013, 61, 29–59. [Google Scholar] [CrossRef]
- Rakotoniriana, E.F.; Scauflaire, J.; Rabemanantsoa, C.; Urveg–Ratsimamanga, S.; Corbisier, A.M.; Quetin–Leclercq, J.; Declerck, S.; Munaut, F. Colletotrichum gigasporum sp. nov.: A new species of Colletotrichum producing long straight conidia. Mycol. Prog. 2013, 12, 403–412. [Google Scholar] [CrossRef]
- Nitzan, N.; Quick, R.A.; Hutson, W.D.; Bamberg, J.; Brown, C.R. Partial resistance to potato black dot, caused by Colletotrichum coccodes in Solanum tuberosum group Andigena. Am. J. Potato Res. 2010, 87, 502–508. [Google Scholar] [CrossRef]
- Kelemu, S.; Badel, J.L. In vitro inhibition of Colletotrichum gloeosporioides and other phytopathogenic fungi by an Amazonian isolate of Bacillus subtilis and its cell–free culture filtrate. Australas. Plant Pathol. 1994, 23, 41–45. [Google Scholar] [CrossRef]
- Pardo-De la Hoz, C.J.; Calderón, C.; Rincón, A.M.; Cárdenas, M.; Danies, G.; López-Kleine, L.; Restrepo, S.; Jiménez, P. Species from the Colletotrichum acutatum; Colletotrichum boninense and Colletotrichum gloeosporioides species complexes associated with tree tomato and mango crops in Colombia. Plant Pathol. 2016, 65, 227–237. [Google Scholar] [CrossRef]
- Ramírez, J.G.; Tamayo, P.J.; Morales, J.G. Identification and pathogenicity of microorganisms affecting purple passion fruit in Colombia. Rev. Ceres. 2017, 64, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Rueda-Hernández, K.R.; Cardona, A.S.; Cadavid-Restrepo, G.E.; Benjumea, C.I.S.; Gutierrez, G.P.C.; Isaza, R.E.A. Differential organ distribution, pathogenicity and benomyl sensitivity of Colletotrichum spp. from blackberry plants in Northern Colombia. J. Phytopathol. 2013, 161, 246–253. [Google Scholar] [CrossRef]
- Marulanda, M.L.; López, A.M.; Isaza, L.; López, P. Microsatellite isolation and characterization for Colletotrichum spp., causal agent of anthracnose in Andean blackberry. Genet. Mol. Res. 2014, 13, 7673–7685. [Google Scholar] [CrossRef]
- López–Vásquez, J.M.; Castaño–Zapata, J.; Marulanda–Ángel, M.L.; López–Gutiérrez, A.M. Characterization of Anthracnose resistance caused by Glomerella cingulata and productivity of five Andean blackberry genotypes (Rubus glaucus Benth). Acta Agronómica 2013, 62, 174–185. [Google Scholar]
- Vanegas–Berrouet, K.M.; Gutierrez, P.A.; Marín, M.A. Identificación molecular de hongos aislados de tejidos de fríjol con síntomas de antracnosis. Acta Biológica Colomb. 2014, 19, 143–153. [Google Scholar]
- Martínez, L.; Vanegas– Berrouet, K.; Salazar, M.; Gutiérrez, P.; Marín, M. Detección por PCR de Colletotrichum lindemuthianum en cultivos y semillas de frijol en Antioquia, Colombia. Acta Agronómica 2014, 63, 377–387. [Google Scholar] [CrossRef]
- Tofiño–Rivera, A.; Chinchilla–Sánchez, K.; Ortega–Cuadros, M. Evaluación in vitro de compatibilidad de agroquímicos, bioinsumos y aceite esencial sobre el control de Colletotrichum gloeosporioides aislado de ají. Vitae 2016, 23, S478. [Google Scholar]
- Viveros–Folleco, Y.M.; Guzmán–Piedrahita, Ó.A.; Villegas–Estrada, B. Enfermedades en viveros comerciales de Musa AAB “Dominico Hartón” en el departamento de Caldas, Colombia. Bol. Cient. Cent. Mus. Mus. Hist. Nat. 2017, 212, 61–80. [Google Scholar]
- Rojas, P.; Pardo–De la Hoz, C.J.; Calderón, C.; Vargas, N.; Cabrera, L.A.; Restrepo, S.; Jiménez, P. First report of Colletotrichum kahawae subsp. ciggaro causing anthracnose disease on tree tomato in Cundinamarca, Colombia. Plant Dis. 2018, 102, 2031. [Google Scholar] [CrossRef]
- López–Zapata, S.P.; Castaño–Zapata, J.; Arango–Isaza, R.; Vásquez–Barajas, D.A. Characterization of the causal agent of the deformation of the fruits of the grape Vitis vinifera L. var. Red Globe, in La Unión, Valle del Cauca, Colombia. Rev. Acad. Colomb. Cienc. Exact. Fis. Nat. 2019, 43, 241–249. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Peterson, A.T. Current and potential distributions of the most important diseases affecting Hass avocado in Antioquia Colombia. J. Plant Prot. Res. 2019, 59, 214–228. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Morales, J.G. Polyphasic identification of preharvest pathologies and disorders in avocado cv. Hass. Agron. Colomb. 2019, 37, 213–227. [Google Scholar] [CrossRef]
- Botina, A.; García, B.L.M.; Romero, Y. Pre-and post-harvest factors that affect the quality and commercialization of the Tahiti lime. Sci. Hortic. 2019, 257, 108737. [Google Scholar] [CrossRef]
- Castañeda-Novoa, C.D.; Vinchira-Villarraga, D.M.; Romero, I.A.G.; Moreno-Sarmiento, N. Evaluation of the production of antifungal metabolites against Colletotrichum gloeosporioides in Streptomyces 5.1 by random mutagenesis. Acta Sci. Biol. Sci. 2021, 43, e54709. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Osorio, J.G.M. Source of inoculum of pathogens, the origin of disorders and diseases management in avocado nurseries. Australas. Plant Pathol. 2021, 50, 457–468. [Google Scholar] [CrossRef]
- Cedeño, L.; Carrero, C. Antracnosis causada por Colletotrichum acutatum en frutos de fresa en Mérida, Venezuela. Interciencia 1997, 22, 315–319. [Google Scholar]
- Andrades, I.; Yender, F.; Labarca, J.; Ulacio, D.; Paredes, C.; Marín, Y. Evaluación de la antracnosis Colletotrichum sp. en guanábana Annona muricata L. tipo Gigante en el sector Moralito del estado Zulia, Venezuela. Rev. Cient. UDO Agríc. 2009, 9, 148–157. [Google Scholar]
- Domínguez–Guerrero, I.P.; Mohali–Castillo, S.R.; Marín–Montoya, M.A.; Pino–Menesini, H.B. Characterization and genetic variability of Colletotrichum gloeosporioides sensu lato in oil palm (Elaeis guineensis Jacq.) plantations from Venezuela. Trop. Plant Pathol. 2012, 37, 108–122. [Google Scholar]
- Urdaneta, L.; Sanabria, M.; Rodríguez, D.; de Camacaro, M.P. Antracnosis causada por Colletotrichum acutatum Simmonds en frutos de fresa en los estados de Lara y Trujillo, Venezuela. Rev. Fac. Agron. 2013, 30, 504–528. [Google Scholar]
- Arocha, I.; Hernández, S.; Araujo, D.; Colina, A. Hongos asociados al cultivo de la cebolla (Allium fistulosum L.) en los municipios Jesús Enrique Lossada y Maracaibo, estado Zulia, Venezuela. Rev. Fac. Agron. Univ. Zulia. 2020, 37, 78–88. Available online: https://produccioncientificaluz.org/index.php/agronomia/article/view/32993 (accessed on 5 December 2021).
- Kelemu, S.; Skinner, D.Z.; Badel, J.L.; Moreno, C.X.; Rodríguez, M.X.; Fernandes, C.D.; Charchar, M.J.; Chakraborty, S. Genetic diversity in South American Colletotrichum gloeosporioides isolates from Stylosanthes guianensis, a tropical forage legume. Eur. J. Plant Pathol. 1999, 105, 261–272. [Google Scholar] [CrossRef]
- Perez, D.; Iannacone, J. Control Químico de la Antracnosis Causado por Colletotrichum gloeosporioides en el Cultivo del Camu Camu (Myrciaria dubia Myrtaceae) en Ucayali, Perú. Fitopatol. Bras. 2006, 31, 518. [Google Scholar] [CrossRef]
- Aguilar, R.; Rafael-Rutte, R.; Martínez-Santos, H.; Apaza-Apaza, S. Agente causal de la antracnosis en el cultivo de arveja (Pisum sativum L.) en el norte de Perú: Sintomatología, aislamiento e identificación, patogenicidad y control. Sci. Agropecu. 2021, 12, 7–14. [Google Scholar] [CrossRef]
- Sicard, D.; Michalakis, Y.; Dron, M.; Neema, C. Genetic diversity and pathogenic variation of Colletotrichum lindemuthianum in the three centers of diversity of its host, Phaseolus vulgaris. Phytopathology 1997, 87, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Falconi, C.E.; Visser, R.G.; van Heusden, S. Influence of plant growth stage on resistance to anthracnose in Andean lupin (Lupinus mutabilis). Crop Pasture Sci. 2015, 66, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Schütz, L. Survey of agricultural practices and alternatives to pesticide use to conserve water resources in the Mojanda Watershed, Ecuador. Future Food J. Food Agric. Soc. 2014, 2, 56–66. [Google Scholar]
- Valencia-Chamorro, S.; Guevara, J.; Paez, D.; Villaplana, R. Hongos caudantes de podedumbres en la pocosecha de Mora (Rubus glaucus): Aislamiento, identificación y evaluación del género más agresivo. Vitae 2016, 23, S769–S773. [Google Scholar]
- Caicedo, J.D.; Lalangui, K.P.; Pozo, A.N.; Cevallos, P.A.; Arahana, V.S.; Méndez, K.S. Multilocus molecular identification and phylogenetic analysis of Colletotrichum tamarilloi as the causal agent of Tamarillo Solanum betaceum anthracnose in the Ecuadorian highlands. Eur. J. Plant Pathol. 2017, 148, 983–996. [Google Scholar] [CrossRef]
- Riera, N.; Ramirez–Villacis, D.; Barriga–Medina, N.; Alvarez–Santana, J.; Herrera, K.; Ruales, C.; Leon–Reyes, A. First report of banana anthracnose caused by Colletotrichum gloeosporioides in Ecuador. Plant Dis. 2019, 103, 763. [Google Scholar] [CrossRef]
- Guevara, Y.; Aponte, A.; Masselli, A. Enfermedades del cultivo de la fresa en dos localidades agrícolas de Aragua y Miranda. INIA Divulga 2004, 3, 22–24. [Google Scholar]
- Farrera, P.; Zambrano, V.; Ortiz, M. Identificación de hongos asociados a enfermedades del fruto de la fresa en el municipio Jáuregui del estado Táchira. Rev. Fac. Agron. 2007, 24, 269–281. [Google Scholar]
- Cedeño, L.; Carrero, C. Antracnosis en plántulas de macadamia causada por Glomerella cingulata. Interciencia 2002, 27, 559–562. [Google Scholar]
- Castro-Navarro, O.M.; Riascos-Ortiz, D.H.; Hoyos-Carvajal, L.M.; Tapiero-Ortiz, A.L. Disturbios histológicos causados por Microcyclus ulei y el complejo Colletotrichum spp. en caucho (Hevea brasiliensis). Summa Phytopathol. 2020, 46, 105–112. [Google Scholar] [CrossRef]
- Bailey, J.A.; O’connell, R.J.; Pring, R.J.; Nash, C. Infection Strategies of Colletotrichum Species; CAB International: Wallingford, UK, 1992; pp. 88–120. [Google Scholar]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, L.; Rojas, P.; Rojas, S.; Pardo-De la Hoz, C.J.; Mideros, M.F.; Danies, G.; Lopez-Kleine, L.; Jiménez, P.; Restrepo, S. Most Colletotrichum species associated with tree tomato Solanum betaceum and mango Mangifera indica crops are not host-specific. Plant Pathol. 2018, 67, 1022–1030. [Google Scholar] [CrossRef]
- Viera, W.; Sotomayor, A.; Tamba Sandoval, M.V.; Vásquez, C.; Martínez, A.; Ron, L. Estimación de parámetros de calidad del fruto para segregantes interespecíficos de tomate de árbol Solanum betaceum Cav. en respuesta de resistencia a la Antracnosis (Colletotrichum acutatum J.H. Simmonds). Acta Agronómica 2016, 65, 304–311. [Google Scholar] [CrossRef]
- Morales, V.; Rodriguez, M. Micobiota endofítica asociada al cultivo del mango ‘Haden’ Mangifera indica L. en el oriente de Venezuela. Rev. UDO Agríc. 2009, 9, 393–402. [Google Scholar]
- Mogollón, Á.M.; López, C.N.; Orduz, J.O. Efecto de las variables meteorológicas sobre la antracnosis (Colletotrichum acutatum) de la lima ácida Tahití en el piedemonte llanero, Colombia. Rev. Acad. Colomb. Cienc. Ex. Fis. Nat. 2021, 45, 250–259. [Google Scholar] [CrossRef]
- Ramírez-Pelayo, C.; Martínez-Quiñones, J.; Gil, J.; Durango, D. Coumarins from the peel of citrus grown in Colombia: Composition, elicitation and antifungal activity. Heliyon 2019, 5, e01937. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.; Gomez, A.; Ramirez, C.; Gil, J.; Durango, D. Controlling anthracnose by means of extracts, and their major constituents, from Brosimum rubescens Taub. Biotechnol. Rep. 2020, 25, e00405. [Google Scholar] [CrossRef]
- Balardin, R.S.; Kelly, J.D. Interaction between Colletotrichum lindemuthianum races and gene pool diversity in Phaseolus vulgaris. J. Am. Soc. Hortic. Sci. 1998, 123, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Weeds, P.L.; Chakraborty, S.; Fernandes, C.D.; Charchar, M.J.D.; Ramesh, C.R.; Kexian, Y.; Kelemu, S. Genetic diversity in Colletotrichum gloeosporioides from Stylosanthes spp. at centers of origin and utilization. Phytopathology 2003, 93, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Falconí, C.E.; Visser, R.G.; van Heusden, A.W. Phenotypic, molecular, and pathological characterization of Colletotrichum acutatum associated with Andean lupine and tamarillo in the Ecuadorian Andes. Plant Dis. 2013, 97, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Correa, G.; Lavalett, L.; Galindo, M.P.; Afanador–Kafuri, L. Uso de métodos multivariantes para la agrupación de aislamientos de Colletotrichum spp. con base en características morfológicas y culturales. Rev. Fac. Nal. Agr. Medellín 2007, 60, 3671–3690. [Google Scholar]
- Mena–Rodríguez, E.; Ortega–Cuadros, M.; Merini, L.; Melo–Ríos, A.E.; Tofiño–Rivera, A. Effect of agricultural inputs and essential oils on the soil of vegetables in Colombia’s Caribbean region. Corpoica Cienc. Tecnol. Agropecu. 2018, 19, 103–124. [Google Scholar] [CrossRef] [Green Version]
- Gamboa–Gaitán, M.Á. Vainillas colombianas y su microbiota. II. Diversidad, cultivo y microorganismos endófitos. Univ. Sci. 2014, 19, 287–300. [Google Scholar] [CrossRef]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal endophyte diversity in coffee plants from Colombia, Hawaii, Mexico and Puerto Rico. Fungal. Ecol. 2010, 3, 122–138. [Google Scholar] [CrossRef]
- Villavicencio–Vásquez, M.; Espinoza–Lozano, R.F.; Pérez–Martínez, S.; Sosa del Castillo, D. Hongos endófitos foliares como candidatos a biocontroladores contra Moniliophthora spp. de Theobroma cacao (Malvaceae) en Ecuador. Acta Biol. Colomb. 2018, 23, 235–241. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Chethana, K.W.T.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X. Mycosphere essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- D’arcy, C.J.; Eastburn, D.M.; Schumann, G.L. Illustrated glossary of plant pathology. In The Plant Health Instructor; The American Phytopathological Society: St. Paul, MN, USA, 2001. [Google Scholar]
- Pariaud, B.; Ravigné, V.; Halkett, F.; Goyeau, H.; Carlier, J.; Lannou, C. Aggressiveness and its role in the adaptation of plant pathogens. Plant Pathol. 2009, 58, 409–424. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Academic Press: Burlington, UK, 2005; p. 952. [Google Scholar]
- Van Wyk, M.; Heath, R.N.; Tarigan, M.; Vermeulen, M.; Wingfield, M.J. Comparison of procedures to evaluate the pathogenicity of Ceratocystis fimbriata sensu lato isolates from Eucalyptus in South Africa. South. For. 2010, 72, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Baroncelli, R.; Amby, D.B.; Zapparata, A.; Sarrocco, S.; Vannacci, G.; Le Floch, G.; Harrison, R.J.; Holub, E.; Sukno, S.; Sreenivasaprasad, S.; et al. Gene family expansions and contractions are associated with host range in plant pathogens of the genus Colletotrichum. BMC Genom. 2016, 17, 555. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Wang, B.; Dong, Q.; Li, L.; Rollins, J.A.; Zhang, R.; Sun, G. Pathogenic adaptations of Colletotrichum fungi revealed by genome wide gene family evolutionary analyses. PLoS ONE 2018, 13, e0196303. [Google Scholar] [CrossRef]
- Russell, P.E. A century of fungicide evolution. J. Agric. Sci. 2005, 143, 11–25. [Google Scholar] [CrossRef]
- Carvalho, F.P. Agriculture, pesticides, food security and food safety. Environ. Sci. Policy 2006, 9, 685–692. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K. Use of endophytes as biocontrol agents. Fungal. Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Suárez–Estrella, F.; Arcos–Nievas, M.A.; López, M.J.; Vargas–García, M.C.; Moreno, J. Biological control of plant pathogens by microorganisms isolated from agroindustrial composts. Biol. Control 2013, 67, 509–515. [Google Scholar] [CrossRef]
- Urrea, R.; Cabezas, L.; Sierra, R.; Cárdenas, M.; Restrepo, S.; Jiménez, P. Selection of antagonistic bacteria isolated from the Physalis peruviana rhizosphere against Fusarium oxysporum. J. Appl. Microbiol. 2011, 111, 707–716. [Google Scholar] [CrossRef]
- Diaz–Herrera, S.; Grossi, C.; Zawoznik, M.; Groppa, M.D. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiol. Res. 2016, 186, 37–43. [Google Scholar] [CrossRef]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Melo, R.A.; Ariza, P.; Lissbrant, S.; Tofiño, A. Evaluation of agrochemicals and bioinputs for sustainable bean management on the Caribbean coast of Colombia. Agron. Colomb. 2015, 3, 203–211. [Google Scholar] [CrossRef]
- Alzate, D.A.; Afanador, L.; Durango, D.L.; García, C.M. Evaluación de la fitotoxicidad y la actividad antifúngica contra Colletotrichum acutatum de los aceites esenciales de tomillo Thymus vulgaris, limoncillo Cymbopogon citratus, y sus componentes mayoritarios. Vitae 2009, 16, 116–125. [Google Scholar]
- Albarracín, L.T.; Delgado, W.; Cuca, L.E.; Ávila, M.C. New butyrolactone and other metabolites from the bark of Endlicheria arenosa against of the phytopathogen Colletotrichum tamarilloi. Nat. Prod. Res. 2019, 33, 687–694. [Google Scholar] [CrossRef]
- España, M.D.; Arboleda, J.W.; Ribeiro, J.A.; Abdelnur, P.V.; Guzman, J.D. Eucalyptus leaf byproduct inhibits the anthracnose–causing fungus Colletotrichum gloeosporioides. Ind. Crop. Prod. 2017, 108, 793–797. [Google Scholar] [CrossRef]
- Martinez, J.; García, C.; Durango, D. Antifungal activity against Colletotrichum acutatum and Colletotrichum gloeosporioides of the major constituents from wood sawdust of Platymiscium gracile Benth. Boletín Latinoam. Caribe Plantas Med. Aromáticas 2017, 161, 14–25. [Google Scholar]
- Pineda, R.; Vizcaíno, S.; García, C.M.; Gil, J.H.; Durango, D. Antifungal activity of extracts, essential oil and constituents from Petroselinum crispum against Colletotrichum acutatum. Rev. Fac. Nac. Agron. Medellín 2018, 71, 8563–8572. [Google Scholar] [CrossRef]
- Osorio, J.; Martínez, E.P.; Hío, J.C. Screening of microbial culture filtrates, plant extracts and fungicides for control of mango anthracnose. Agron. Colomb. 2012, 30, 222–229. [Google Scholar]
- Ramírez, S.; Arias, J.D.; Bedoya, J.C.; Rueda, E.A.; Sánchez, C.Y.; Granada, S.D. Metabolites produced by antagonistic microbes inhibit the principal avocado pathogens in vitro. Agron. Colomb. 2015, 33, 58–63. [Google Scholar] [CrossRef]
- Pastrana–Camacho, N.; Suárez, Z.; Acosta–González, A.; Arango, C.; Haltli, B.; Correa, H.; Kerr, R.; Duque, C.; Díaz, L.E. Bioprospecting for culturable actinobacteria with antimicrobial properties isolated from rivers in Colombian Orinoquia. Trop. J. Pharm. Res. 2016, 15, 1259–1265. [Google Scholar] [CrossRef] [Green Version]
- López–Vásquez, J.M.; Angel, M.L.M.; Gutierrez, A.M.L. Factores climáticos y su influencia en la expresión de enfermedades fúngicas en cultivares de Heliconias. Rev. Univ. Sci. 2013, 18, 331–345. [Google Scholar] [CrossRef]
- Bolívar, K.; Sanabria, M.E.; Rodríguez, D.; de Camacaro, M.P.; Ulacio, D.; Cumana, L.J.; Crescente, O. Potencial efecto fungicida de extractos vegetales en el desarrollo in vitro del hongo Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. y de la antracnosis en frutos de mango. Rev. Cient. UDO Agríc. 2009, 91, 175–181. [Google Scholar]
- Pineda, J.; Principal, J.; Barrios, C.; Milla, D.; Solano, Y.; Gil, E. In vitro fungistatic property of propolis on three Colletotrichum gloeosporioides isolates. Zootec. Trop. 2010, 281, 83–91. [Google Scholar]
- Mochizuki, M.; Yamamoto, S.; Aoki, Y.; Suzuki, S. Isolation and characterisation of Bacillus amyloliquefaciens S13–3 as a biological control agent for anthracnose caused by Colletotrichum gloeosporioides. Biocontrol. Sci. Technol. 2012, 22, 697–709. [Google Scholar] [CrossRef]
- Thilagam, R.; Hemalatha, N. Plant growth promotion and chilli anthracnose disease suppression ability of rhizosphere soil actinobacteria. J. Appl. Microbiol. 2019, 126, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Khakvar, R.; AliasgarZad, N. Application of endophytic bacteria for controlling anthracnose disease Colletotrichum lindemuthianum on bean plants. Arch. Phytopathol. Plant Prot. 2013, 46, 1831–1838. [Google Scholar] [CrossRef]
- Damasceno, C.L.; Duarte, E.A.; dos Santos, L.B.P.R.; de Oliveira, T.A.S.; de Jesus, F.N.; de Oliveira, L.M.; Góes–Neto, A.; Soares, A.C.F. Postharvest biocontrol of anthracnose in bananas by endophytic and soil rhizosphere bacteria associated with sisal (Agave sisalana) in Brazil. Biol. Control. 2019, 137, 104016. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Udayanga, D.; Cai, L.; Chukeatirote, E.; Hyde, K.D. Endophytic Colletotrichum from tropical grasses with a new species C. endophytica. Fungal Divers. 2013, 61, 107–115. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Urdaneta, L.; Araujo, D.; Quirós, M.; Rodríguez, D.; Colmenares, C.B.; Poleo, N.; Petit, Y.; Dorado, I. Micobiota endófita asociada a estadios preflorales del guayabo (Psidium guajava L.) y al ácaro plano (Brevipalpus phoenicis) (Geijskes) (Acari: Tenuipalpidae). Rev. Cient. UDO Agríc. 2009, 9, 166–174. [Google Scholar]
- Quintero–Mercado, A.; Dangon–Bernier, F.; Páez–Redondo, A. Aislamientos endofíticos de Colletotrichum spp. a partir de hojas y ramas de mango Mangifera indica L. cultivar Azúcar en el municipio de Ciénaga, Magdalena, Colombia. Rev. Acad. Colomb. Cienc. Exact. Fis. Nat. 2019, 43, 65–77. [Google Scholar] [CrossRef]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, P.; Chen, R.; Pang, J.; Wang, R.; Zhang, Y.; et al. Colletotrichum gloeosporioides sl associated with Theobroma cacao and other plants in Panama: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338. [Google Scholar] [CrossRef] [Green Version]
- Lenné, J.M.; Brown, A.E. Factors influencing the germination of pathogenic and weakly pathogenic isolates of Colletotrichum gloeosporiodes on leaf surfaces of Stylosanthes guianensis. Mycol. Res. 1991, 95, 227–232. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Species | Countries | Observations |
|---|---|---|---|
| C. acutatum species complex | C. acutatum | Colombia, Ecuador | |
| C. godetiae | Colombia | ||
| C. laticiphilum | Colombia | ||
| C. tamarilloi | Colombia, Ecuador | ||
| C. boninense species complex | C. annellatum | Colombia | |
| C. colombiense | Colombia | ||
| C. boninense | Colombia | ||
| C. brassicicola | Colombia | ||
| C. karstii | Colombia | ||
| C. gigasporum species complex | C. gigasporum | Colombia | Endophyte |
| C. gloeosporioides species complex | C. aenigma | Colombia | |
| C. asianum | Colombia | ||
| C. cigarro | Colombia | Formerly labeled C. kahawae | |
| C. fructicola | Colombia | ||
| C. gloeosporioides ss. | Colombia | ||
| C. gloeosporioides sl. | Colombia, Ecuador | ||
| C. gossypii | Colombia | Unverified | |
| C. siamense | Colombia | ||
| C. theobromicola | Colombia | ||
| C. tropicale | Colombia | ||
| C. orbiculare species complex | C. lindemuthianum | Colombia, Peru | Peru unverified |
| Singleton species | C. coccodes | Colombia | Unverified |
| Family | Species | Common Name |
|---|---|---|
| Anacardiaceae | Manguifera indica L. | Mango |
| Annonaceae | Annona muricata L. | Soursop |
| Areacaceae | Elaeis guinensis Jacq. | Oil palm, African palm |
| Caricaceae | Carica papaya L. | Papaya |
| Euphorbiaceae | Hevea brasiliensis Muell. | Rubber |
| Fabaceae | Phaseolus vulgaris L. | Common bean |
| Lupinus mutabilis Sweet | Tarwi, Lupin, Chocho, Altramuz | |
| Stylosanthes guianensis Aubl. | Stylosanthes | |
| Heliconiaceae | Heliconia spp. | Heliconia, platanillo |
| Lauraceae | Persea americana Mill. | Avocado |
| Malvaceae | Theobroma cacao L. | Cocoa |
| Gossypium spp. | Cotton | |
| Musaceae | Musa AAB Simmonds | Platane |
| Passifloraceae | Passiflora edulis f. edulis Sims | Gulupa, Poro poro, purple passion fruit |
| Proteaceae | Macadamia integrifolia | Macadamia |
| Rosaceae | Rubus glaucus Benth. | Andean raspberry |
| Fragaria × ananassa | Strawberry | |
| Rubiaceae | Coffea arabica L. | Coffee |
| Rutaceae | Citrus latifolia Tanaka | Tahiti Lima |
| Solanaceae | Capsicum spp. L. | Pepper |
| Solanum melongena L. | Eggplant | |
| Solanum betaceum Cav. | Tamarillo, tree tomato | |
| Solanum tuberosum | Potato Group Andigena |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guevara-Suarez, M.; Cárdenas, M.; Jiménez, P.; Afanador-Kafuri, L.; Restrepo, S. Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review. Agronomy 2022, 12, 548. https://doi.org/10.3390/agronomy12030548
Guevara-Suarez M, Cárdenas M, Jiménez P, Afanador-Kafuri L, Restrepo S. Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review. Agronomy. 2022; 12(3):548. https://doi.org/10.3390/agronomy12030548
Chicago/Turabian StyleGuevara-Suarez, Marcela, Martha Cárdenas, Pedro Jiménez, Lucía Afanador-Kafuri, and Silvia Restrepo. 2022. "Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review" Agronomy 12, no. 3: 548. https://doi.org/10.3390/agronomy12030548
APA StyleGuevara-Suarez, M., Cárdenas, M., Jiménez, P., Afanador-Kafuri, L., & Restrepo, S. (2022). Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review. Agronomy, 12(3), 548. https://doi.org/10.3390/agronomy12030548