
‘Breathing Out’ under Heat Stress—Respiratory Control of Crop Yield under High Temperature


 ,
, 
Abstract
:1. Introduction
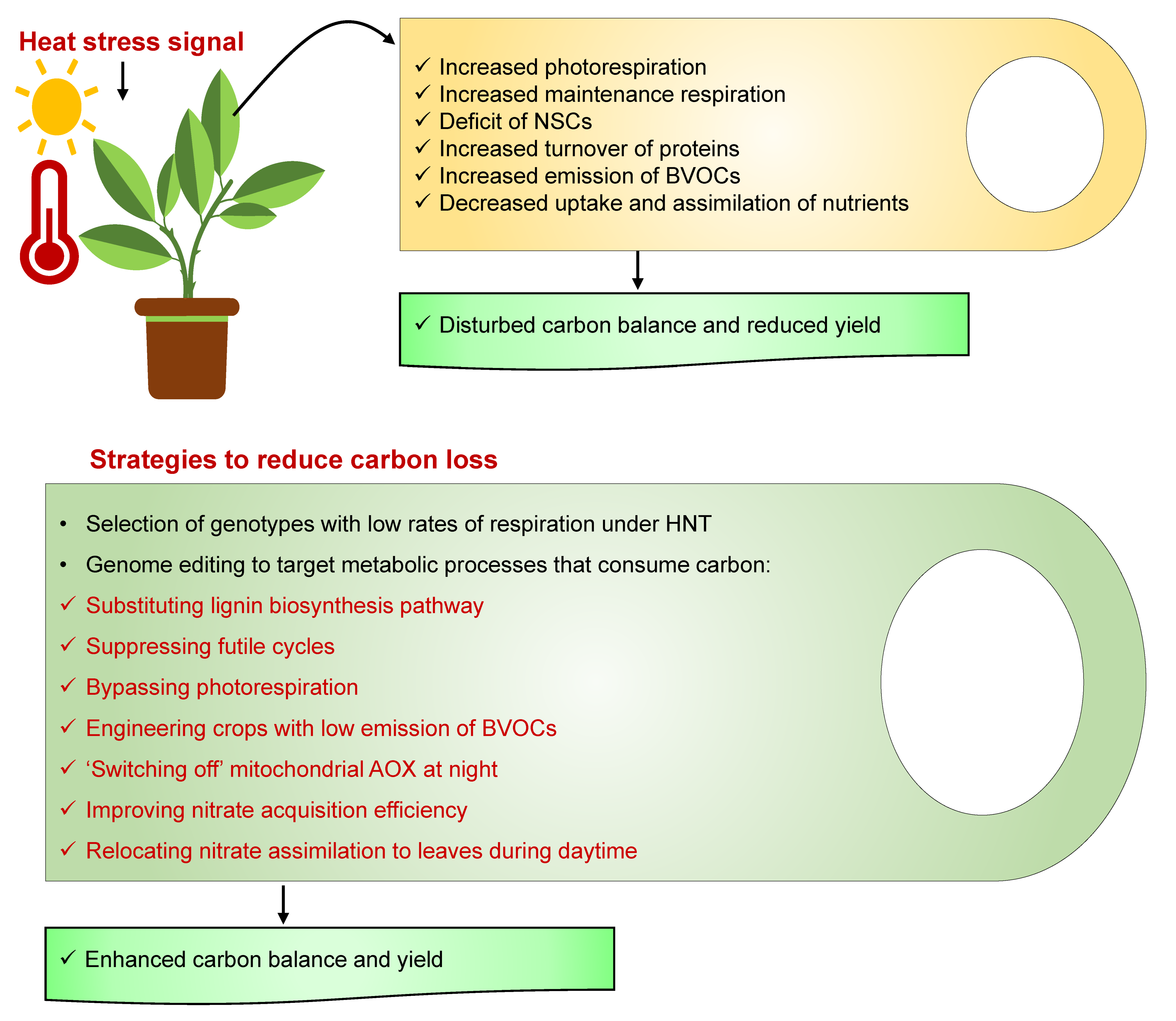
2. Respiratory Carbon Loss-A Constraint to Crop Yield
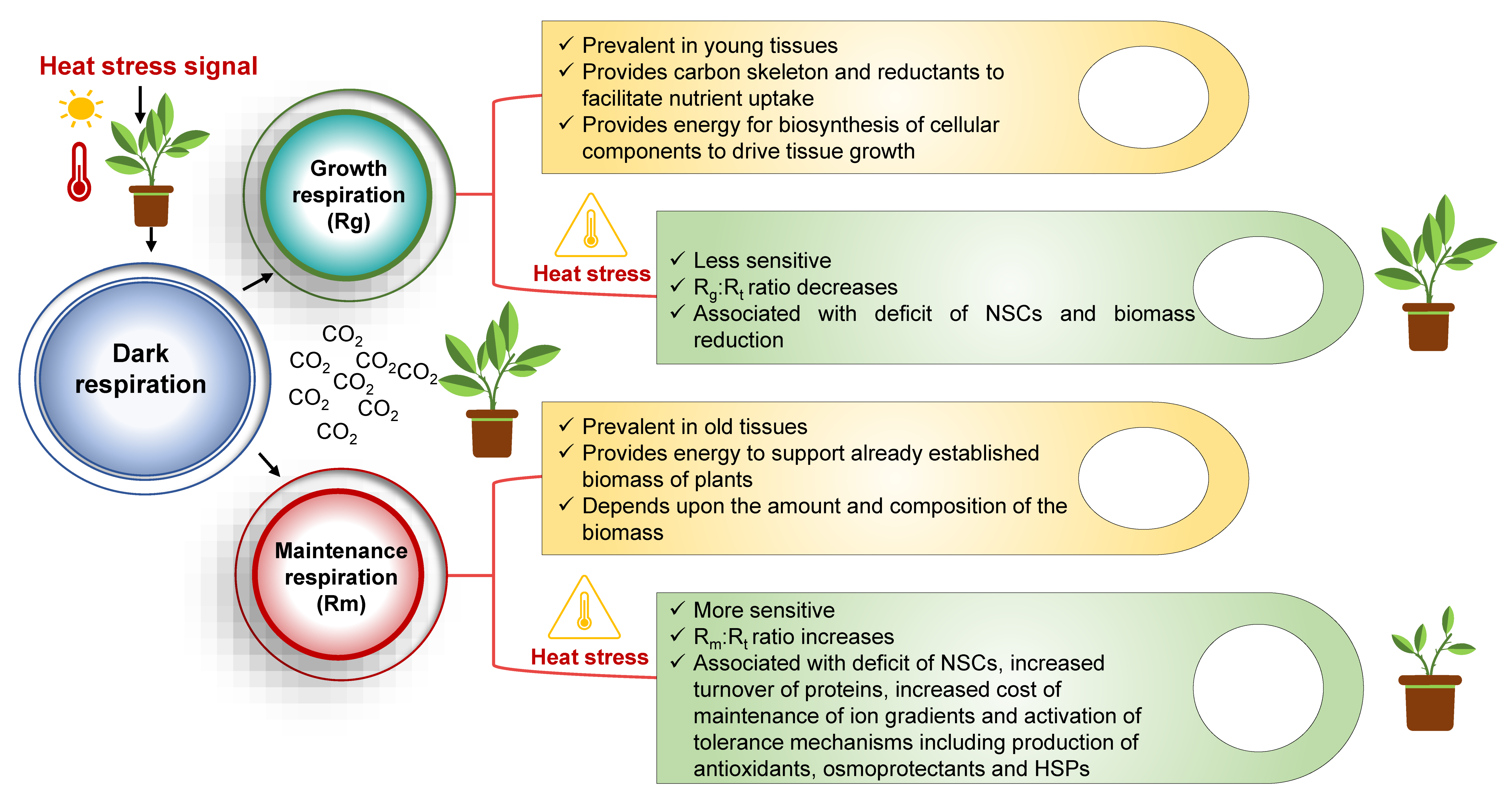
3. Heat-Induced Changes in the Proportion of Maintenance Respiration
4. Substrate Availability for Respiration under High Temperature
5. Regulation of Respiratory Flux at High Temperature
6. Positive Correlation between Protein Turnover Cost and Respiratory Cost at High Temperature
7. Diurnal Dynamics of Respiration
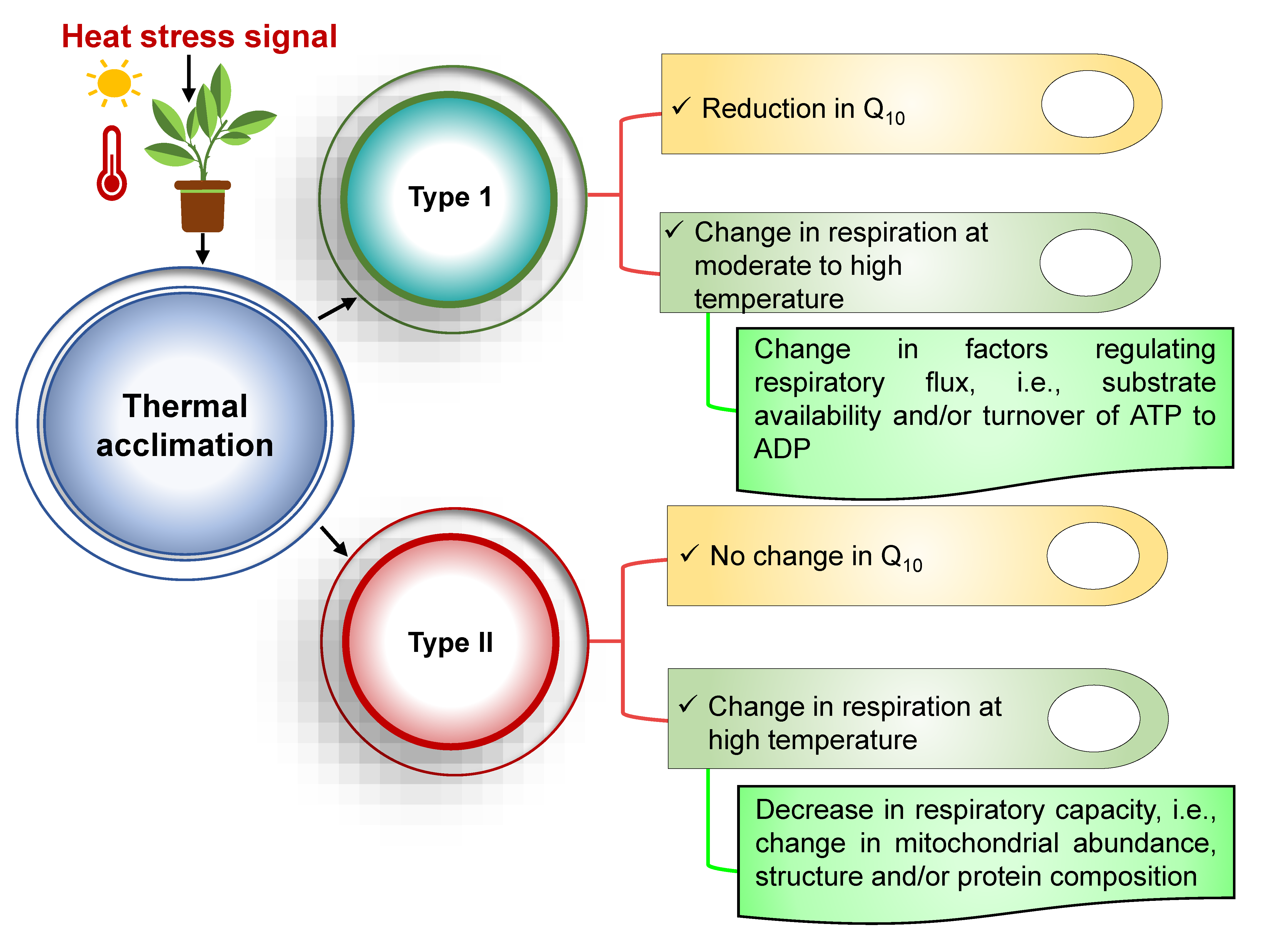
8. Thermal Acclimation of the Respiration Response in Plants under Heat Stress
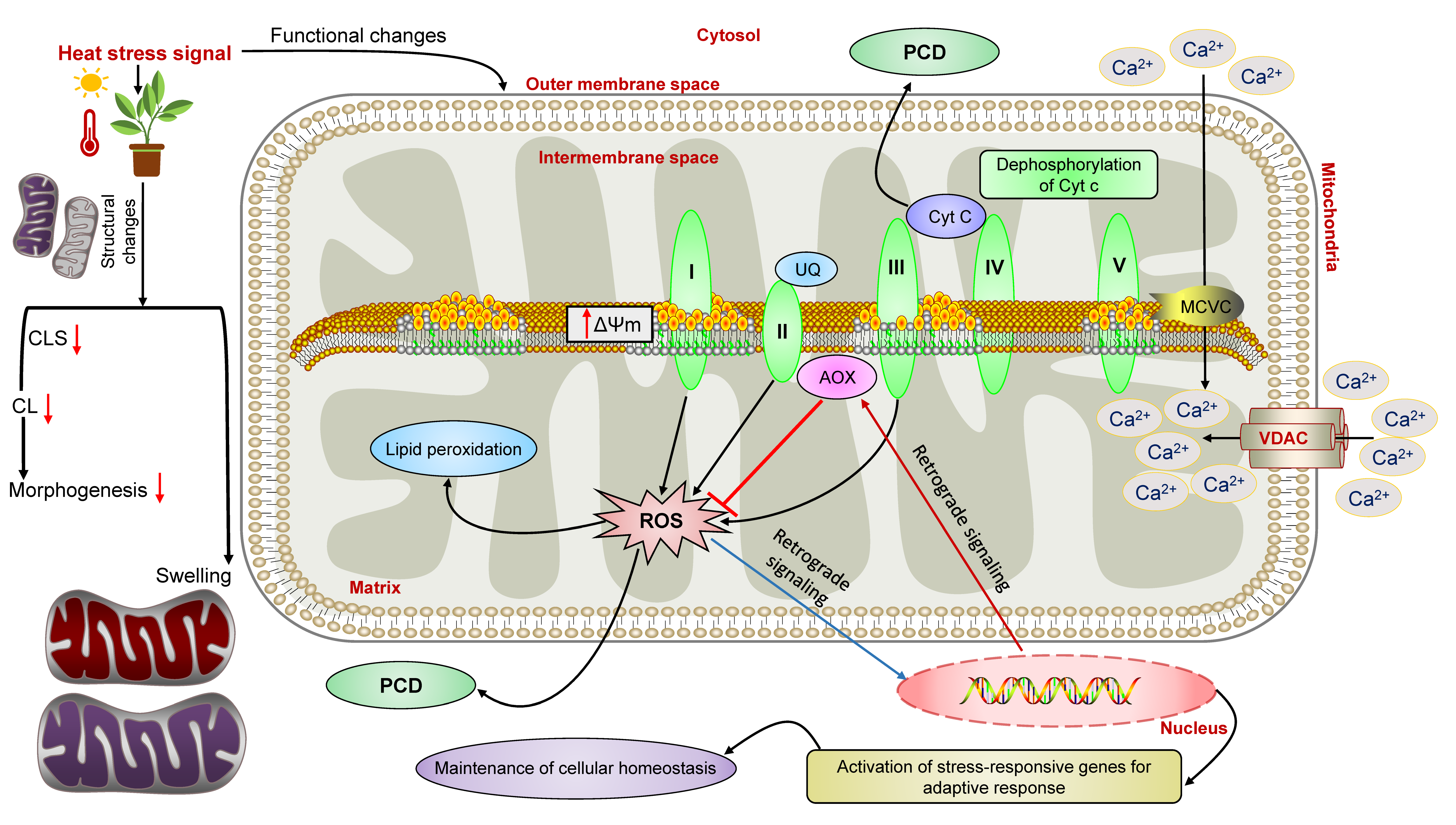
9. Mitochondrial Physiology under High Temperature
10. Hormonal Regulation of Respiratory Metabolism under High Temperature
11. Strategies to Reduce Carbon Loss
11.1. Selection of Genotypes with Low Rates of Respiration under High Night Temperature
11.2. Genome Editing to Target the Metabolic Processes Consuming Carbon
11.2.1. Substitution of the Lignin Biosynthesis Pathway
11.2.2. Suppression of Futile Cycles
11.2.3. Designing Carbon Conserving Photorespiration
11.2.4. Engineering for Low Emission of Biogenic Volatile Organic Compounds
11.2.5. “Switching Off” Mitochondrial AOX at Night
11.2.6. Improving Nitrate Acquisition and Relocating Nitrate Assimilation
12. Conclusions and Future Outlooks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hüve, K.; Bichele, I.; Ivanova, H.; Keerberg, O.; Pärnik, T.; Rasulov, B.; Tobias, M.; Niinemets, U. Temperature responses of dark respiration in relation to leaf sugar concentration. Physiol. Plant. 2012, 144, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Heskel, M.A.; O’sullivan, O.S.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, L.K.; Penillard, A.; Atkin, O.K. Convergence in the temperature response of leaf respiration across biomes and plant functional types. Proc. Natl. Acad. Sci. USA 2016, 113, 3832–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadok, W.; Jagadish, S.K. The hidden costs of nighttime warming on yields. Trends Plant Sci. 2020, 25, 644–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, W.H. Biogeochemistry: An Analysis of Global Change, 2nd ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Amthor, J.S. Terrestrial higher-plant response to increasing atmospheric [CO2] in relation to the global carbon cycle. Glob. Chang. Biol. 1995, 1, 243–274 . [Google Scholar] [CrossRef]
- Waring, R.H.; Landsberg, J.J.; Williams, M. Net primary production of forests: A constant fraction of gross primary production? Tree Physiol. 1998, 18, 129–134. [Google Scholar] [CrossRef]
- Gifford, R.M. The global carbon cycle: A viewpoint on the missing sink. Aust. J. Plant Physiol. 1994, 21, 1–15. [Google Scholar] [CrossRef]
- Gifford, R.M.; Thorne, J.H.; Hitz, W.D.; Giaquinta, R.T. Crop productivity and photoassimilate partitioning. Science 1984, 225, 801–808. [Google Scholar] [CrossRef]
- Amthor, J.S. Evolution and applicability of a whole plant respiration model. J. Theor. Biol. 1986, 122, 473–490 . [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Pean, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. 2022; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Li, G.; Chen, T.; Feng, B.; Peng, S.; Tao, L.; Fu, G. Respiration, rather than photosynthesis, determines rice yield loss under moderate high-temperature conditions. Front. Plant Sci. 2021, 12, 1287. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Reich, P.B. Modelling respiration of vegetation: Evidence for a general temperature-dependent Q10. Glob. Chang. Biol. 2001, 7, 223–230. [Google Scholar] [CrossRef]
- Heskel, M.A.; Bitterman, D.; Atkin, O.K.; Turnbull, M.H.; Griffin, K.L. Seasonality of foliar respiration in two dominant plant species from the Arctic tundra: Response to long-term warming and short-term temperature variability. Funct. Plant Biol. 2014, 41, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, L.K.; Creek, D.; Crous, K.Y.; Xiang, S.; Liddell, M.J.; Turnbull, M.H.; Atkin, O.K. Canopy position affects the relationships between leaf respiration and associated traits in a tropical rainforest in Far North Queensland. Tree Physiol. 2014, 34, 564–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, O.S.; Heskel, M.A.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, L.K.; Penillard, A.; Zhu, L.L.; Egerton, J.J.G.; Bloomfield, K.J.; Creek, D.; et al. Thermal limits of leaf metabolism across biomes. Glob. Chang. Biol. 2017, 23, 209–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kromer, S. Respiration during photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 45–70. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2018, 221, 32–49. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, A.P.; South, P.F.; Bernacchi, C.J.; Ort, D.R. Alternative pathway to photorespiration protects growth and productivity at elevated temperatures in a model crop. Plant Biotechnol. J. 2021. [CrossRef]
- Nunes-Nesi, A.; Carrari, F.; Lytovchenko, A.; Smith, A.M.; Loureiro, M.E.; Ratcliffe, R.G.; Sweetlove, L.J.; Fernie, A.R. Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants. Plant Physiol. 2005, 137, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Hauben, M.; Haesendonckx, B.; Standaert, E.; Van Der Kelen, K.; Azmi, A.; Akpo, H.; Van Breusegem, F.; Guisez, Y.; Bots, M.; Lambert, B.; et al. Energy use efficiency is characterized by an epigenetic component that can be directed through artificial selection to increase yield. Proc. Natl. Acad. Sci. USA 2009, 106, 20109–20114. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; VanLoocke, A.; Bernacchi, C.J.; Ort, D.R. The costs of photorespiration to food production now and in the future. Annu. Rev. Plant Biol. 2016, 67, 107–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Yin, X.; He, S.; Jiang, W.; Si, C.; Struik, P.C.; Luo, W.; Li, G.; Xie, Y.; Xiong, Y.; et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Glob. Chang. Biol. 2016, 22, 856–874. [Google Scholar] [CrossRef] [PubMed]
- Impa, S.M.; Raju, B.; Hein, N.T.; Sandhu, J.; Prasad, P.V.V.; Walia, H.; Jagadish, S.K. High night temperature effects on wheat and rice: Current status and way forward. Plant Cell Environ. 2021, 44, 2049–2065. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Misra, G.; Sreenivasulu, N.; Henry, A. What happens at night? Physiological mechanisms related to maintaining grain yield under high night temperature in rice. Plant Cell Environ. 2021, 44, 2245–2261. [Google Scholar] [CrossRef] [PubMed]
- Tombesi, S.; Cincera, I.; Frioni, T.; Ughini, V.; Gatti, M.; Palliotti, A.; Poni, S. Relationship among night temperature, carbohydrate translocation and inhibition of grapevine leaf photosynthesis. Environ. Exp. Bot. 2019, 157, 293–298 . [Google Scholar] [CrossRef]
- Glaubitz, U.; Erban, A.; Kopka, J.; Hincha, D.K.; Zuther, E. High night temperature strongly impacts TCA cycle, amino acid and polyamine biosynthetic pathways in rice in a sensitivity-dependent manner. J. Exp. Bot. 2015, 66, 6385–6397. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Henry, A.; Sreenivasulu, N. Rice yield formation under high day and night temperatures-A prerequisite to ensure future food security. Plant Cell Environ. 2020, 43, 1595–1608. [Google Scholar] [CrossRef]
- Lambers, H. Respiration in intact plants and tissues: Its regulation and dependence on environmental factors, metabolism and invaded organisms. In Higher Plant Cell Respiration; Douce, R., Day, D.A., Eds.; (Encyclopedia of Plant Physiology), New Series; Springer: Berlin, Germany, 1985; Volume 18. [Google Scholar]
- Slot, M.; Kitajima, K. General patterns of acclimation of leaf respiration to elevated temperatures across biomes and plant types. Oecologia 2015, 177, 885–900. [Google Scholar] [CrossRef] [Green Version]
- Amthor, J.S.; Bar-Even, A.; Hanson, A.D.; Millar, A.H.; Stitt, M.; Sweetlove, L.J.; Tyerman, S.D. Engineering strategies to boost crop productivity by cutting respiratory carbon loss. Plant Cell 2019, 31, 297–314. [Google Scholar] [CrossRef] [Green Version]
- Amthor, J.S. The McCree-de Wit-Penning de Vries-Thornley respiration paradigms: 30 years later. Ann. Bot. 2000, 86, 1–20 . [Google Scholar] [CrossRef] [Green Version]
- Adu-Bredu, S.; Yokota, T.; Hagihara, A. Temperature effect on maintenance and growth respiration coefficients of young, field-grown hinoki cypress (Chamaecyparisobtusa). Ecol. Res. 1997, 12, 357–362. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bloomfield, K.J.; Reich, P.B.; Tjoelker, M.G.; Asner, G.P.; Bonal, D.; Bönisch, G.; Bradford, M.G.; Cernusak, L.; Cosio, E.; et al. Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 2015, 206, 614–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, K.Y.; Uddling, J.; De Kauwe, M.G. Temperature responses of photosynthesis and respiration in evergreen trees from boreal to tropical latitudes. New Phytol. 2022, 234, 353–374. [Google Scholar] [CrossRef]
- Thornley, J.H.M. Respiration, growth and maintenance in plants. Nature 1970, 227, 304–305. [Google Scholar] [CrossRef] [PubMed]
- McCree, K.J. Maintenance requirements of white clover at high and low growth rates. Crop Sci. 1982, 22, 345–351. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; Johnson, I.R. Plant and Crop Modeling. Clarendon Press Oxford: Oxford, UK, 1990. [Google Scholar]
- Van Iersel, M.W. Respiratory Q10 of marigold (Tagetes patula) in response to long-term temperature differences and its relationship to growth and maintenance respiration. Physiol. Plant. 2006, 128, 289–301. [Google Scholar] [CrossRef]
- Winzeler, H.; Hunt, L.A.; Mahon, J.D. Ontogenetic changes in respiration and photosynthesis in a uniculm barley. Crop Sci. 1976, 16, 786–790. [Google Scholar] [CrossRef]
- McCree, K.J.; Silsbury, J.H. Growth and maintenance requirements of subterranean clover. Crop Sci. 1978, 18, 13–18. [Google Scholar] [CrossRef]
- Mariko, S.; Koizumi, H. Respiration for maintenance and growth in Reynoutria japonica ecotypes from different altitudes on Mt Fuji. Ecol. Res. 1993, 8, 241–246. [Google Scholar] [CrossRef]
- Gifford, R.M. Whole plant respiration and photosynthesis of wheat under increased CO2 concentration and temperature: Long-term vs. short-term distinctions for modelling. Glob. Chang. Biol. 1995, 1, 385–396. [Google Scholar] [CrossRef]
- Tan, K.; Zhou, G.; Ren, S. Response of leaf dark respiration of winter wheat to changes in CO2 concentration and temperature. Chin. Sci. Bull. 2013, 58, 1795–1800. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Bruhn, D.; Tjoelker, M.G. Response of plant respiration to changes in temperature: Mechanisms and consequences of variations in Q10 values and acclimation. In Advances in Photosynthesis and Respiration, Plant Respiration; Lambers, H., Ribas-Carbo, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 18. [Google Scholar] [CrossRef]
- Atkin, O.K.; Macherel, D. The crucial role of plant mitochondria in orchestrating drought tolerance. Ann. Bot. 2009, 103, 581–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, M.; Sharma, P.; Anand, A.; Pandita, V.K.; Bhatia, A.; Pushkar, S. Raffinose and hexose sugar content during germination are related to infrared thermal fingerprints of primed onion (Allium cepa L.) seeds. Front. Plant Sci. 2020, 11, 579037. [Google Scholar] [CrossRef] [PubMed]
- Ögren, E. Maintenance respiration correlates with sugar but not nitrogen concentration in dormant plants. Physiol. Plant. 2000, 108, 295–299. [Google Scholar] [CrossRef]
- Whitehead, D.; Griffin, K.L.; Turnbull, M.H.; Tissue, D.T.; Engel, V.C.; Brown, K.J.; Schuster, W.S.; Walcroft, A.S. Response of total night-time respiration to differences in total daily photosynthesis for leaves in a Quercus rubra L. canopy: Implications for modelling canopy CO2 exchange. Glob. Chang. Biol. 2004, 10, 925–938. [Google Scholar] [CrossRef]
- Noguchi, K.; Terashima, I. Different regulation of leaf respiration between Spinacia oleracea, a sun species, and Alocasia odora, a shade species. Physiol. Plant. 1997, 101, 1–7. [Google Scholar] [CrossRef]
- Hoefnagel, M.H. Interdependence between chloroplasts and mitochondria in the light and the dark. Biochim. Biophys. ActaBioenerg. 1998, 1366, 235–255. [Google Scholar] [CrossRef] [Green Version]
- Douce, R.; Neuburger, M. The uniqueness of plant mitochondria. Annu. Rev. Plant Biol. 1980, 40, 371–414. [Google Scholar] [CrossRef]
- Zerihun, A.; Mckenzie, B.A.; Morton, J.D. Photosynthate costs associated with the utilization of different nitrogen-forms: Influence on the carbon balance of plants and shoot-root biomass partitioning. New Phytol. 1998, 138, 1–11. [Google Scholar] [CrossRef]
- Williams, K.; Percival, F.; Merino, J.; Mooney, H.A. Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant Cell Environ. 1987, 10, 725–734. [Google Scholar] [CrossRef]
- Hachiya, T.; Terashima, I.; Noguchi, K. Increase in respiratory cost at high growth temperature is attributed to high protein turnover cost in Petunia x hybrida petals. Plant Cell Environ. 2007, 30, 1269–1283. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.P.; Eubel, H.; Millar, A.H. Diurnal changes in mitochondrial function reveal daily optimization of light and dark respiratory metabolism in Arabidopsis. Mol. Cell. Proteom. 2010, 9, 2125–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasing, O.E.; Gibon, Y.; Günther, M.; Höhne, M.; Morcuende, R.; Osuna, D.; Thimm, O.; Usadel, B.; Scheible, W.R.; Stitt, M. Sugars and circadian regulation make major contributions to the global regulation of diurnal gene expression in Arabidopsis. Plant Cell 2005, 17, 3257–3281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shameer, S.; Ratcliffe, R.G.; Sweetlove, L.J. Leaf energy balance requires mitochondrial respiration and export of chloroplast NADPH in the light. Plant Physiol. 2019, 180, 1947–1961 . [Google Scholar] [CrossRef]
- Gibon, Y.; Pyl, E.-T.; Sulpice, R.; Lunn, J.E.; Hohne, M.; Gunther, M.; Stitt, M. Adjustment of growth, starch turnover, protein content and central metabolism to a decrease of the carbon supply when Arabidopsis is grown in very short photoperiods. Plant Cell Environ. 2009, 32, 859–874. [Google Scholar] [CrossRef]
- O’Leary, B.M.; Lee, C.P.; Atkin, O.K.; Cheng, R.; Brown, T.B.; Millar, A.H. Variation in leaf respiration rates at night correlates with carbohydrate and amino acid supply. Plant Physiol. 2017, 174, 2261–2273. [Google Scholar] [CrossRef] [Green Version]
- Gibon, Y.; Usadel, B.; Blaesing, O.E.; Kamlage, B.; Hoehne, M.; Trethewey, R.; Stitt, M. Integration of metabolite with transcript and enzyme activity profiling during diurnal cycles in Arabidopsis rosettes. Genome Biol. 2006, 7, R76. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, C.; Sato, S.; Yanagisawa, S.; Uesono, Y.; Terashima, I.; Noguchi, K. Effects of elevated CO2 on levels of primary metabolites and transcripts of genes encoding respiratory enzymes and their diurnal patterns in Arabidopsis thaliana: Possible relationships with respiratory rates. Plant Cell Physiol. 2014, 55, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Rashid, F.A.A.; Crisp, P.A.; Zhang, Y.; Berkowitz, O.; Pogson, B.J.; Day, D.A.; Masle, J.; Dewar, R.C.; Whelan, J.; Atkin, O.K. Molecular and physiological responses during thermal acclimation of leaf photosynthesis and respiration in rice. Plant Cell Environ. 2020, 43, 594–610. [Google Scholar] [CrossRef]
- Cook, D.; Fowler, S.; Fiehn, O.; Thomashow, M. A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis. Proc. Natl. Acad. Sci.USA 2004, 101, 15243–15248. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Bruhn, D.; Hurry, V.M.; Tjoelker, M. The hot and the cold: Unraveling the variable response of plant respiration to temperature. Funct. Plant Biol. 2005, 32, 87–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Wei, X.; Rich, R.L.; Montgomery, R.A. Boreal and temperate trees show strong acclimation of respiration to warming. Nature 2016, 531, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.G.; Dukes, J.S. Short-term acclimation to warmer temperatures accelerates leaf carbon exchange processes across plant types. Glob. Chang. Biol. 2017, 23, 4840–4853. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.G.; Dukes, J.S. Plant respiration and photosynthesis in global-scale models: Incorporating acclimation to temperature and CO2. Glob. Chang. Biol. 2013, 19, 45–63. [Google Scholar] [CrossRef]
- Armstrong, A.F.; Logan, D.C.; Atkin, O.K. On the developmental dependence of leaf respiration: Responses to short-and long-term changes in growth temperature. Am. J. Bot. 2006, 93, 1633–1639. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Reich, P.B. Acclimation of respiration to temperature and CO2 in seedlings of boreal tree species in relation to plant size and relative growth rate. Glob. Chang. Biol. 1999, 5, 679–691. [Google Scholar] [CrossRef]
- Rurek, M. Plant mitochondria under a variety of temperature stress conditions. Mitochondrion 2014, 19, 289–294. [Google Scholar] [CrossRef]
- Logan, D.C. Mitochondrial fusion, division and positioning in plants. Biochem. Soc. Trans. 2010, 38, 789–795. [Google Scholar] [CrossRef]
- Taylor, N.L.; Heazlewood, J.L.; Day, D.A.; Millar, A.H. Differential impact of environmental stresses on the pea mitochondrial proteome. Mol. Cell. Proteom. 2005, 4, 1122–1133. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.H.; Xiao, Y.A.; Zeng, J.J.; Hu, X.H. Photosynthesis, respiration and antioxidant enzymes in pepper leaves under drought and heat stresses. Biol. Plant 2010, 54, 761–765. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Crawford, T.; Lehotai, N.; Strand, A. The role of retrograde signals during plant stress responses. J. Exp. Bot. 2018, 69, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Osman, C.; Voelker, D.R.; Langer, T. Making heads or tails of phospholipids in mitochondria. J. Cell Biol. 2011, 192, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.H.; Jones, A.D.; Hu, J.P. Cardiolipin-mediated mitochondrial dynamics and stress response in Arabidopsis. Plant Cell 2014, 26, 391–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welchen, E.; Gonzalez, D.H. Cytochrome c, a hub linking energy, redox, stress and signaling pathways in mitochondria and other cell compartments. Physiol. Plant. 2016, 157, 310–321 101111/ppl12449. [Google Scholar] [CrossRef]
- Zancani, M.; Casolo, V.; Petrussa, E.; Peresson, C.; Patui, S.; Bertolini, A.; De Col, V.; Braidot, E.; Boscutti, F.; Vianello, A. The permeability transition in plant mitochondria: The missing link. Front. Plant Sci. 2015, 6, 1120. [Google Scholar] [CrossRef] [Green Version]
- Petereit, J.; Katayama, K.; Lorenz, C.; Ewert, L.; Schertl, P.; Kitsche, A.; Wada, H.; Frentzen, M.; Braun, H.-P.; Eubel, H. Cardiolipin supports respiratory enzymes in plants in different ways. Front. Plant Sci. 2017, 8, 72 . [Google Scholar] [CrossRef] [Green Version]
- Van Aken, O.; Van Breusegem, F. Licensed to Kill: Mitochondria, Chloroplasts, and Cell Death. Trends Plant Sci. 2015, 20, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Pedrotti, L.; Weiste, C.; Nagele, T.; Wolf, E.; Lorenzin, F.; Dietrich, K.; Mair, A.; Weckwerth, W.; Teige, M.; Baena-Gonzalez, E.; et al. Snf1-RELATED KINASE1-controlled C/S1- bZIP signaling activates alternative mitochondrial metabolic pathways to ensure plant survival in extended darkness. Plant Cell 2018, 30, 495–509. [Google Scholar] [CrossRef] [Green Version]
- Dourmap, C.; Roque, S.; Morin, A.; Caubriere, D.; Kerdiles, M.; Beguin, K.; Perdoux, R.; Reynoud, N.; Bourdet, L.; Audebert, P.-A.; et al. Stress signalling dynamics of the mitochondrial electron transport chain and oxidative phosphorylation system in higher plants. Ann. Bot. 2020, 125, 721–736. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jardim-Messeder, D.; Caverzan, A.; Rauber, R.; De souza Ferreira, E.; Margis-pinheiro, M.; Galina, A. Succinate dehydrogenase (mitochondrial complex II) is a source of reactive oxygen species in plants and regulates development and stress responses. New Phytol. 2015, 208, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Fedyaeva, A.V.; Stepanov, A.V.; Lyubushkina, I.V.; Pobezhimova, T.P.; Rikhvanov, E.G. Heat shock induces production of reactive oxygen species and increases inner mitochondrial membrane potential in winter wheat cells. Biochemistry 2014, 79, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Rikhvanov, E.G.; Fedoseeva, I.V.; Pyatrikas, D.V.; Borovskii, G.B.; Voinikov, V.K. Role of mitochondria in the operation of calcium signaling system in heat-stressed plants. Russ. J. Plant Physiol. 2014, 61, 141–153. [Google Scholar] [CrossRef]
- Vothknecht, U.C.; Szabo, I. Channels and transporters for inorganic ions in plant mitochondria: Prediction and facts. Mitochondrion 2020, 53, 224–233. [Google Scholar] [CrossRef]
- Jazwinski, S.M. The retrograde response: When mitochondrial quality control is not enough. Biochim. Biophys. Acta-Mol. Cell Res. 2013, 1833, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Giraud, E.; Van Aken, O.; Ho, L.H.; Whelan, J. The transcription factor ABI4 is a regulator of mitochondrial retrograde expression of ALTERNATIVE OXIDASE1a. Plant Physiol. 2009, 150, 1286–1296. [Google Scholar] [CrossRef] [Green Version]
- Parsons, H.L.; Yip, J.Y.H.; Vanlerberghe, G.C. Increased respiratory restriction during phosphate-limited growth in transgenic tobacco cells lacking alternative oxidase. Plant Physiol. 1999, 121, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase modulates leaf mitochondrial concentrations of superoxide and nitric oxide. New Phytol. 2012, 195, 32–39. [Google Scholar] [CrossRef]
- Berkowitz, O.; Clercq, I.D.; Breusegem, F.V.; Whelan, J. Interaction between hormonal and mitochondrial signalling during growth, development and in plant defense responses. Plant Cell Environ. 2016, 39, 1127–1139. [Google Scholar] [CrossRef]
- Maestri, E.; Klueva, N.; Perrotta, C.; Gulli, M.; Nguyen, H.T.; Marmiroli, N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol. 2002, 48, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, A.; Pieterse, C. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liang, X.; Huang, J.; Zhang, D.; Lu, H.; Liu, Z.; Bi, Y. Involvement of ethylene and hydrogen peroxide in induction of alternative respiratory pathway in salt-treated Arabidopsis calluses. Plant Cell Physiol. 2010, 51, 1754–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verslues, P.E. ABA and cytokinins: Challenge and opportunity for plant stress research. Plant Mol. Biol. 2016, 91, 629–640. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Norman, C.; Howell, K.A.; Millar, A.H.; Whelan, J.M.; Day, D.A. Salicylic acid is an uncoupler and inhibitor of mitochondrial electron transport. Plant Physiol. 2004, 134, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Czarnocka, W.; Karpinski, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Van den Bergen, C.W.; Wagner, A.M.; Krab, K.; Moore, A.L. The relationship between electron flux and the redox poise of the quinone pool in plant mitochondria: Interplay between quinol-oxidizing and quinone reducing pathways. Eur. J. Biochem. 1994, 226, 1071–1078. [Google Scholar] [CrossRef]
- Millar, A.H.; Atkin, O.K.; Lambers, H.; Wiskich, J.T.; Day, D.A. A critique of the use of inhibitors to estimate partitioning of electrons between mitochondrial respiratory pathways in plants. Physiol. Plant. 1995, 95, 523–532. [Google Scholar] [CrossRef]
- Liao, Y.; Tian, M.; Zhang, H.; Li, X.; Wang, Y.; Xia, X.; Zhou, J.; Zhou, Y.; Yu, J.; Shi, K.; et al. Salicylic acid binding of mitochondrial alpha-ketoglutarate dehydrogenase E2 affects mitochondrial oxidative phosphorylation and electron transport chain components and plays a role in basal defense against tobacco mosaic virus in tomato. New Phytol. 2015, 205, 1296–1307. [Google Scholar] [CrossRef]
- Miller, C.O. Cytokinin modification of mitochondrial function. Plant Physiol. 1982, 69, 1274–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberto, D.; Couee, I.; Sulmon, C.; Gouesbet, G. Root-level exposure reveals multiple physiological toxicity of triazine xenobiotics in Arabidopsis thaliana. J. Plant Physiol. 2017, 212, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, D.; Yang, C. AtFtsH4 perturbs the mitochondrial respiratory chain complexes and auxin homeostasis in Arabidopsis. Plant Signal. Behav. 2014, 9, e29709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerchev, P.I.; Clercq, I.D.; Denecker, J.; Muhlenbock, P.; Kumpf, R.; Nguyen, L.; Audenaert, D.; Dejonghe, W.; Breusegem, F.V. Mitochondrial perturbation negatively affects auxin signaling. Mol. Plant. 2014, 7, 1138–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Murayama, M.; Hayashi, S.; Nishimura, N.; Ishide, M.; Kobayashi, K.; Yagi, Y.; Asami, T.; Nakamura, T.; Shinozaki, K.; Hirayama, T. Isolation of Arabidopsis ahg11, a weak ABA hypersensitive mutant defective in nad4 RNA editing. J. Exp. Bot. 2012, 63, 5301–5310. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Dugardeyn, J.; Zhang, C.; Muhlenbock, P.; Eastmond, P.J.; Valcke, R.; De Coninck, B.; Oden, S.; Karampelias, M.; Cammue, B.P.A.; et al. The Arabidopsis thaliana RNA editing factor SLO2, which affects the mitochondrial electron transport chain, participates in multiple stress and hormone responses. Mol Plant. 2014, 7, 290–310. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, L.-J.; Gong, Z. DEXH box RNA helicase-mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar] [CrossRef] [Green Version]
- Sechet, J.; Roux, C.; Plessis, A.; Effroy, D.; Frey, A.; Perreau, F.; Biniek, C.; Krieger-Liszkar, A.; Macherel, D.; North, H.N.; et al. The ABA-deficiency suppressor locus HAS2 encodes the PPR protein LOI1/MEF11 involved in mitochondrial RNA editing. Mol. Plant. 2015, 8, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Watkins, C.B. Advances in the use of 1-MCP. In Advances in Postharvest Fruit and Vegetable Technology; CRC Press: Boca Raton, FL, USA, 2015; pp. 117–145. [Google Scholar] [CrossRef]
- Deng, X.G.; Zhu, T.; Zhang, D.W.; Lin, H.H. The alternative respiratory pathway is involved in brassinosteroid-induced environmental stress tolerance in Nicotiana benthamiana. J. Exp. Bot. 2015, 66, 6219–6232. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Yadav, A.; Khetarpal, S.; Anand, A.; Sathee, L.; Kumar, R.R.; Singh, B.; Soora, N.K.; Pushkar, S. High day-night transition temperature alters nocturnal starch metabolism in rice (Oryza sativa L.). Acta Physiol. Plant 2017, 39, 74. [Google Scholar] [CrossRef]
- Shi, W.; Yin, X.; Struik, P.C.; Solis, C.; Xie, F.; Schmidt, R.C.; Huang, M.; Zou, Y.; Ye, C.; Jagadish, S.K. High day-and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J. Exp. Bot. 2017, 68, 5233–5245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahuguna, R.N.; Solis, C.A.; Shi, W.; Jagadish, K.S. Post-flowering night respiration and altered sink activity account for high night temperature-induced grain yield and quality loss in rice (Oryza sativa L.). Physiol.Plant. 2017, 159, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Yadav, A.; Anand, A.; Khetarpal, S.; Kumar, D.; Trivedi, S.M. Adverse effect of increase in minimum temperature during early grain filling period on grain growth and quality in indica rice (Oryza sativa) cultivars. Indian J. Agric. Sci. 2017, 87, 883–888. [Google Scholar]
- Barros, J.; Serrani-Yarce, J.C.; Chen, F.; Baxter, D.; Venables, B.J.; Dixon, R.A. Role of bifunctional ammonia-lyase in grass cell wall biosynthesis. Nat.Plants 2016, 2, 16050. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, O.; Ishihara, H.; George, G.M.; Mengin, V.; Flis, A.; Sumner, D.; Arrivault, S.; Feil, R.; Lunn, J.E.; Zeeman, S.C.; et al. Leaf starch turnover occurs inlong days and in falling light at the end of the day. Plant Physiol. 2017, 174, 2199–2212. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.P.; Vigeolas, H.; Raymond, P.; Rolin, D.; Dieuaide-Noubhani, M. A new substrate cycle in plants. Evidence fora high glucose-phosphate-to-glucose turnover from in vivo steady-state and pulse-labeling experiments with [13C] glucose and [14C] glucose. Plant Physiol. 2005, 138, 2220–2232 . [Google Scholar] [CrossRef] [Green Version]
- Hill, S.A.; Ap Rees, T. Fluxes of carbohydrate metabolism in ripening bananas. Planta 1994, 192, 52–60. [Google Scholar] [CrossRef]
- Trudeau, D.L.; Edlich-Muthm, C.; Zarzycki, J.; Scheffen, M.; Goldsmith, M.; Khersonsky, O.; Avizemer, Z.; Fleishman, S.J.; Cotton, C.A.R.; Erb, T.J.; et al. Design and in vitro realization of carbon-conserving photorespiration. Proc. Natl. Acad. Sci. USA 2018, 115, E11455–E11464. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Singsaas, E.L. Why plants emit isoprene. Nature 1995, 374, 769. [Google Scholar] [CrossRef]
- Loreto, F.; Forster, A.; Durr, M.; Csiky, O.; Seufert, G. On the monoterpene emission under heat stress and on the increased thermotolerance of leaves of Quercus ilex L. fumigated with selected monoterpenes. Plant Cell Environ. 1998, 21, 101–107. [Google Scholar] [CrossRef]
- Cheah, M.H.; Millar, A.H.; Myers, R.C.; Day, D.A.; Roth, J.; Hillier, W.; Badger, M.R. Online oxygen kinetic isotope effects using membrane inlet mass spectrometry can differentiate between oxidases for mechanistic studies and calculation of their contributions to oxygen consumption in whole tissues. Anal. Chem. 2014, 86, 5171–5178. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Cannell, M.G.R.; Thornley, J.H.M. Modeling the components of plant respiration: Some guiding principles. Ann. Bot. 2000, 85, 45–54. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Experimental Temperature | Q10 Value | Reference |
|---|---|---|---|
| Marigold (Tagetes patula) | 20 °C (Control) 30 °C (Elevated) | 1.35–1.55 | [40] |
| Barley (Hordeum vulgare) | 15 °C (Control) 28 °C (Elevated) | 3.00 | [41] |
| Subterranean clover (Trifolium subterraneum) | 10 °C (Control) 35 °C (Elevated) | 1.85 | [42] |
| Japanese knotweed (Reynoutria japonica) | 15 °C (Control) 25 °C (Elevated) | 1.90 | [43] |
| Wheat (Triticum aestivum) | 15 °C (Control) 20 °C (Elevated) | 1.80 | [44] |
| 10 °C (Control-Night temperature) 21 °C (Elevated-Night temperature) | 1.97 | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, N.; Thakur, M.; Suryakumar, P.; Mukherjee, P.; Raza, A.; Prakash, C.S.; Anand, A. ‘Breathing Out’ under Heat Stress—Respiratory Control of Crop Yield under High Temperature. Agronomy 2022, 12, 806. https://doi.org/10.3390/agronomy12040806
Sharma N, Thakur M, Suryakumar P, Mukherjee P, Raza A, Prakash CS, Anand A. ‘Breathing Out’ under Heat Stress—Respiratory Control of Crop Yield under High Temperature. Agronomy. 2022; 12(4):806. https://doi.org/10.3390/agronomy12040806
Chicago/Turabian StyleSharma, Nitin, Meenakshi Thakur, Pavithra Suryakumar, Purbali Mukherjee, Ali Raza, Channapatna S. Prakash, and Anjali Anand. 2022. "‘Breathing Out’ under Heat Stress—Respiratory Control of Crop Yield under High Temperature" Agronomy 12, no. 4: 806. https://doi.org/10.3390/agronomy12040806
APA StyleSharma, N., Thakur, M., Suryakumar, P., Mukherjee, P., Raza, A., Prakash, C. S., & Anand, A. (2022). ‘Breathing Out’ under Heat Stress—Respiratory Control of Crop Yield under High Temperature. Agronomy, 12(4), 806. https://doi.org/10.3390/agronomy12040806