
Antifungal Activity of Methylxanthines against Grapevine Trunk Diseases

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemical Reagents
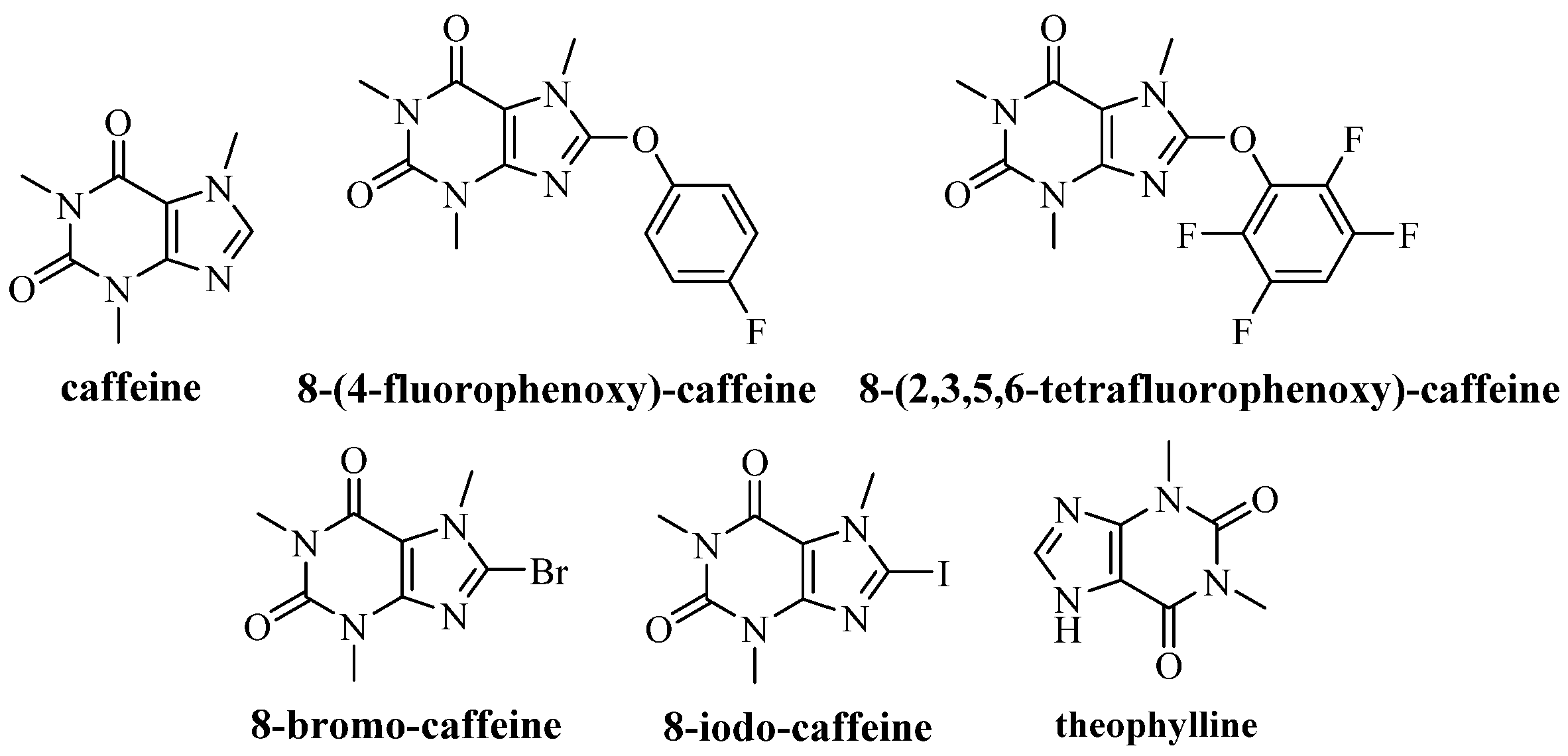
2.2. Synthesis of 8-(4-fluorophenoxy)-caffeine and 8-(2,3,5,6-tetrafluorophenoxy)-caffeine
2.3. Preparation of Conjugate Complexes
2.4. Characterization of Caffeine Derivatives by Nuclear Magnetic Resonance Spectroscopy
2.5. Physico-Chemical Characterization of Conjugated Complexes
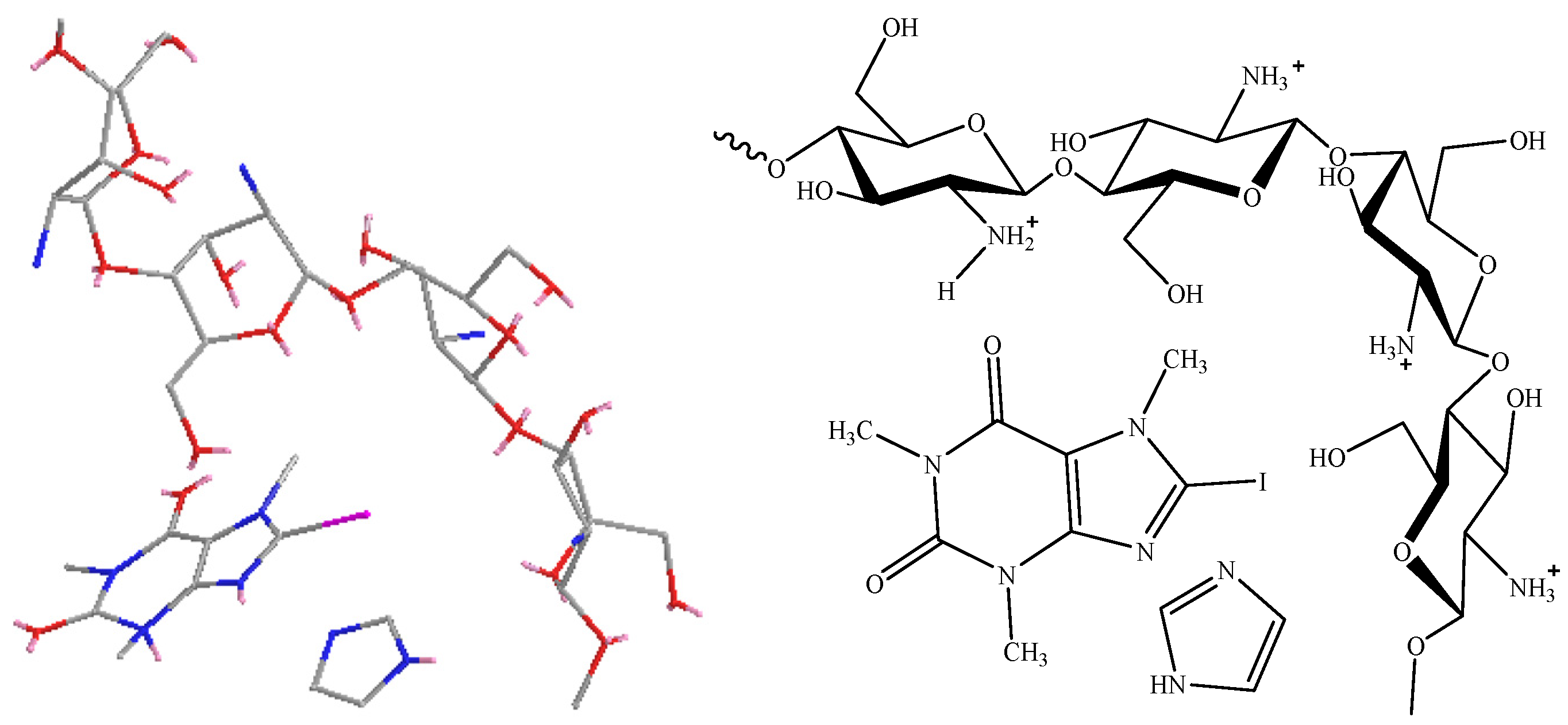
2.6. Energy Content and Molecular Structure of Imidazole Complexes with Caffeine Derivatives
2.7. Fungal Isolates
2.8. In Vitro Antimicrobial Activity Assessment
2.9. Statistical Analysis
3. Results
3.1. Characterization of Caffeine Derivatives by Nuclear Magnetic Resonance Spectroscopy
3.2. Vibrational Characterization
3.2.1. COS–Caffeine Derivative Conjugates
3.2.2. COS–Caffeine Derivative-Imidazole Complexes
3.3. Thermal Analysis of Conjugated Compounds
3.4. Energy Content and Molecular Structure of Imidazole Interaction with Caffeine Derivatives
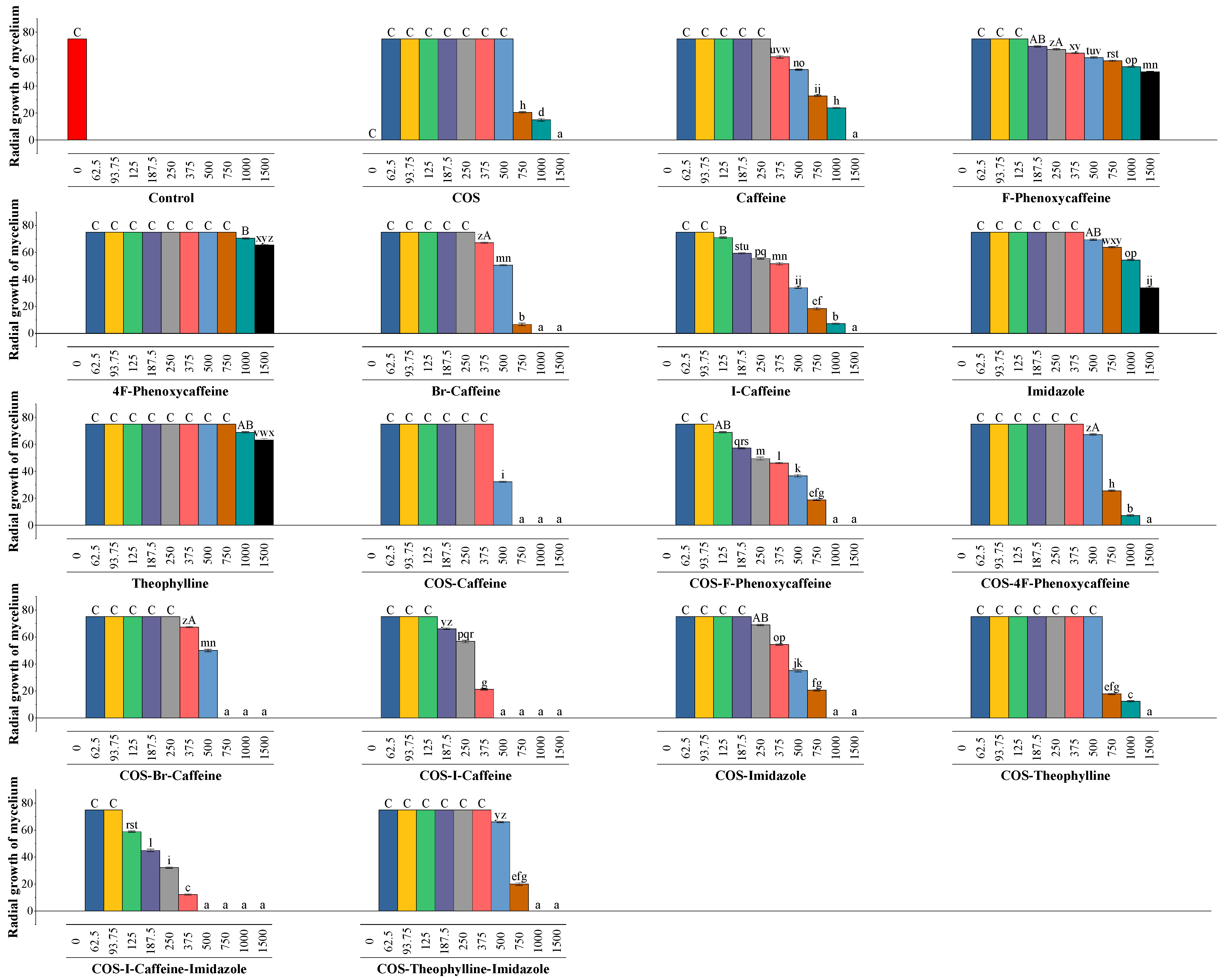
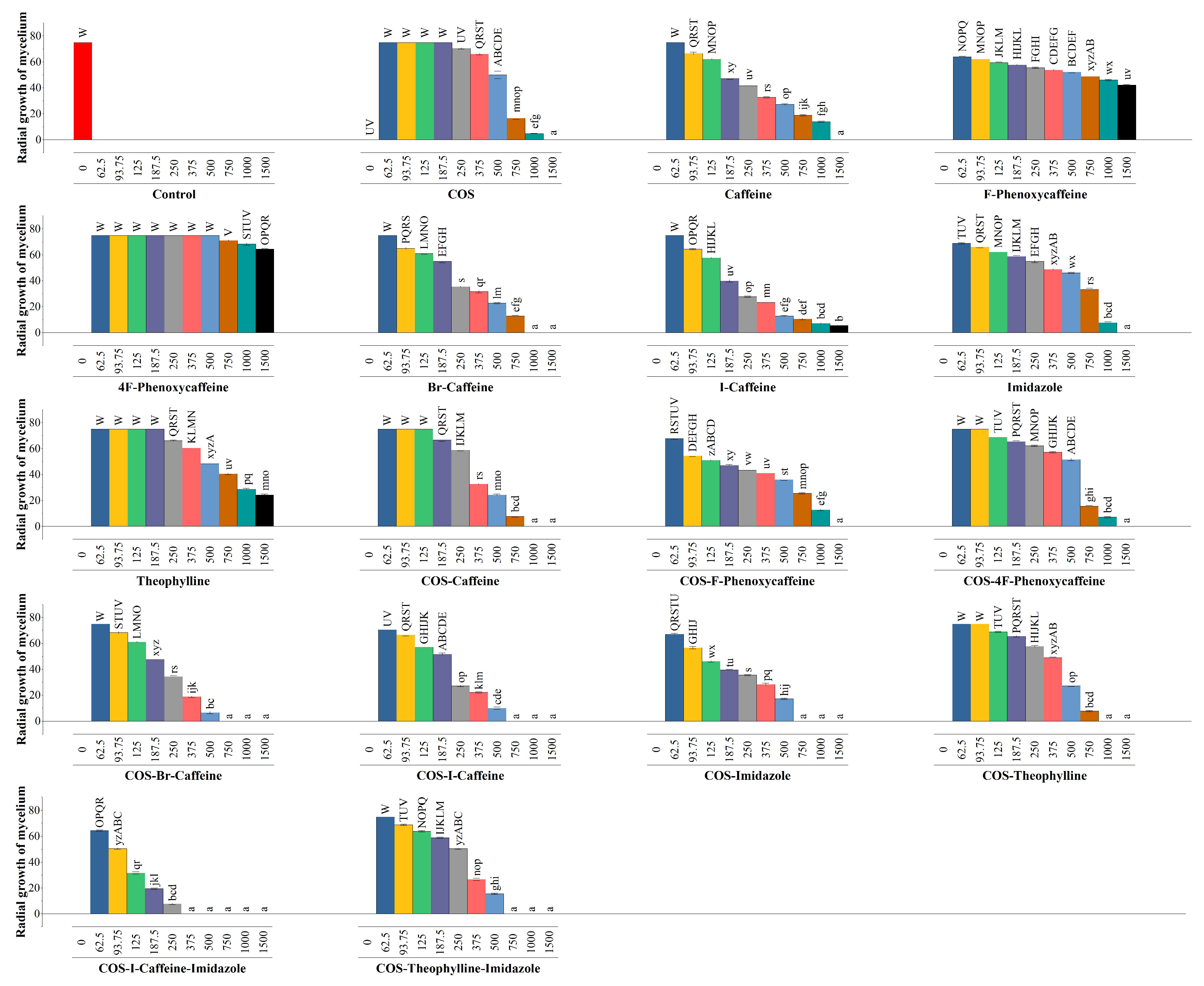
3.5. In Vitro Antimicrobial Activity
4. Discussion
4.1. On the Structure of the Complexes
4.2. On the Activity of the Complexes
4.2.1. Comparison with Imidazole as a Fungicide Control
4.2.2. Comparison with Reported Antifungal Activities of Methylxanthines against Wood-Degrading Fungi
4.3. On Possible Mechanisms of Action
4.4. Limitations of the Study and Future Research Lines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Liu, H.; Yang, N.; Xiong, L.; Wang, B. Synthesis and evaluation of novel xanthine-acrylamides and xanthine-acrylates as insecticidal agents. Pest Manag. Sci. 2022. Early view. [Google Scholar] [CrossRef]
- Singh, S.; Jain, L.; Pandey, M.B.; Singh, U.P.; Pandey, V.B. Antifungal activity of the alkaloids from Eschscholtzia californica. Folia Microbiol. 2009, 54, 204–206. [Google Scholar] [CrossRef]
- Tsirilakis, K.; Kim, C.; Vicencio, A.G.; Andrade, C.; Casadevall, A.; Goldman, D.L. Methylxanthine inhibit fungal chitinases and exhibit antifungal activity. Mycopathologia 2012, 173, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Pánek, M.; Borůvka, V.; Nábělková, J.; Šimůnková, K.; Zeidler, A.; Novák, D.; Černý, R.; Kobetičová, K. Efficacy of caffeine treatment for wood protection—influence of wood and fungi species. Polymers 2021, 13, 3758. [Google Scholar] [CrossRef]
- Kobetičová, K.; Nábělková, J.; Ďurišová, K.; Šimůnková, K.; Černý, R. Antifungal activity of methylxanthines based on their properties. BioResources 2020, 15, 8110–8120. [Google Scholar] [CrossRef]
- Šimůnková, K.; Reinprecht, L.; Nábělková, J.; Hýsek, Š.; Kindl, J.; Borůvka, V.; Lišková, T.; Šobotník, J.; Pánek, M. Caffeine—Perspective natural biocide for wood protection against decaying fungi and termites. J. Clean. Prod. 2021, 304, 127110. [Google Scholar] [CrossRef]
- Kobeticova, K.; Böhm, M.; Nábělková, J.; Černý, R. Influence of selected storage temperatures on wood properties and its biological resistance after the use of methylxanthines. BioResources 2021, 16, 13. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clement, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Xing, R.; Liu, S.; Li, P. Chitin and Chitosan Fragments Responsible for Plant Elicitor and Growth Stimulator. J. Agric. Food. Chem. 2020, 68, 12203–12211. [Google Scholar] [CrossRef]
- Abosabaa, S.A.; ElMeshad, A.N.; Arafa, M.G. Chitosan nanocarrier entrapping hydrophilic drugs as advanced polymeric system for dual pharmaceutical and cosmeceutical application: A comprehensive analysis using Box-Behnken design. Polymers 2021, 13, 677. [Google Scholar] [CrossRef]
- Seyedabadi, M.M.; Rostami, H.; Jafari, S.M.; Fathi, M. Development and characterization of chitosan-coated nanoliposomes for encapsulation of caffeine. Food Biosci. 2021, 40, 100857. [Google Scholar] [CrossRef]
- Popa, N.; Novac, O.; Profire, L.; Hritcu, D.; Popa, M.I. Inclusion and release of theophylline from chitosan based microparticles. Turk. J. Chem. 2010, 34, 255–262. [Google Scholar]
- Samanta, H.S.; Ray, S.K. Controlled release of tinidazole and theophylline from chitosan based composite hydrogels. Carbohydr. Polym. 2014, 106, 109–120. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Fernandez-Arrojo, L.; Mengibar, M.; Belmonte-Reche, E.; Peñalver, P.; Acosta, F.N.; Ballesteros, A.O.; Morales, J.C.; Kidibule, P.; Fernandez-Lobato, M.; et al. Enzymatic production of fully deacetylated chitooligosaccharides and their neuroprotective and anti-inflammatory properties. Biocatal. Biotransform. 2017, 36, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Ruano-Rosa, D.; Sánchez-Hernández, E.; Baquero-Foz, R.; Martín-Ramos, P.; Martín-Gil, J.; Torres-Sánchez, S.; Casanova-Gascón, J. Chitosan-based bioactive formulations for the control of powdery mildew in viticulture. Agronomy 2022, 12, 495. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [Green Version]
- Levy, Y.; Benderly, M.; Cohen, Y.; Gisi, U.; Bassand, D. The joint action of fungicides in mixtures: Comparison of two methods for synergy calculation. EPPO Bull. 1986, 16, 651–657. [Google Scholar] [CrossRef]
- Parab, R.H.; Dixit, B.; Desai, D.J. Synthesis, characterization and antimicrobial activity of imidazole derivatives. Asian J. Chem. 2011, 23, 2725–2728. [Google Scholar]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Arora, D.S.; Ohlan, D. In vitro studies on antifungal activity of tea (Camellia sinensis) and coffee (Coffea arabica) against wood-rotting fungi. J. Basic Microbiol. 1997, 37, 159–165. [Google Scholar] [CrossRef]
- Kwaśniewska-Sip, P.; Cofta, G.; Nowak, P.B. Resistance of fungal growth on Scots pine treated with caffeine. Int. Biodeterior. Biodegrad. 2018, 132, 178–184. [Google Scholar] [CrossRef]
- Sugiyama, A.; Sano, C.M.; Yazaki, K.; Sano, H. Caffeine fostering of mycoparasitic fungi against phytopathogens. Plant Signal. Behav. 2016, 11, e1113362. [Google Scholar] [CrossRef] [Green Version]
- Reinke, A.; Chen, J.C.Y.; Aronova, S.; Powers, T. Caffeine targets TOR complex I and provides evidence for a regulatory link between the FRB and kinase domains of Tor1p. J. Biol. Chem. 2006, 281, 31616–31626. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Qian, W.-J.; Li, N.-N.; Hao, X.-Y.; Wang, L.; Xiao, B.; Wang, X.-C.; Yang, Y.-J. Metabolic changes of caffeine in tea plant (Camellia sinensis (L.) O. Kuntze) as defense response to Colletotrichum fructicola. J. Agric. Food. Chem. 2016, 64, 6685–6693. [Google Scholar] [CrossRef]
- Singh, S.; Fatima, Z.; Ahmad, K.; Hameed, S. Repurposing of respiratory drug theophylline against Candida albicans: Mechanistic insights unveil alterations in membrane properties and metabolic fitness. J. Appl. Microbiol. 2020, 129, 860–875. [Google Scholar] [CrossRef]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro- and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Davidse, L.C. Benzimidazole fungicides: Mechanism of action and biological impact. Annu. Rev. Phytopathol. 1986, 24, 43–65. [Google Scholar] [CrossRef]
- Nasrollahi, Z.; Yadegari, M.H. Antifungal activity of caffeine in combination with fluconazole against Candida albicans. Infect. Epidemiol. Med. 2016, 2, 18–21. [Google Scholar] [CrossRef]
- Tayel, A.A.; Moussa, S.H.; Salem, M.F.; Mazrou, K.E.; El-Tras, W.F. Control of citrus molds using bioactive coatings incorporated with fungal chitosan/plant extracts composite. J. Sci. Food Agric. 2016, 96, 1306–1312. [Google Scholar] [CrossRef]
- Sabaa, M.W.; Mohamed, N.A.; Mohamed, R.R.; Khalil, N.M.; Abd El Latif, S.M. Synthesis, characterization and antimicrobial activity of poly (N-vinyl imidazole) grafted carboxymethyl chitosan. Carbohydr. Polym. 2010, 79, 998–1005. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Langa-Lomba, N.; González-García, V.; Casanova-Gascón, J.; Martín-Gil, J.; Santiago-Aliste, A.; Torres-Sánchez, S.; Martín-Ramos, P. Lignin–chitosan nanocarriers for the delivery of bioactive natural products against wood-decay phytopathogens. Agronomy 2022, 12, 461. [Google Scholar] [CrossRef]
- Dias, M.C. Phytotoxicity: An overview of the physiological responses of plants exposed to fungicides. J. Bot. 2012, 2012, 135479. [Google Scholar] [CrossRef] [Green Version]
- Buzón-Durán, L.; Langa-Lomba, N.; González-García, V.; Casanova-Gascón, J.; Sánchez-Hernández, E.; Martín-Gil, J.; Martín-Ramos, P. Rutin-stevioside and related conjugates for potential control of grapevine trunk diseases. Phytopathol. Mediterr. 2022, 61, 65–77. [Google Scholar] [CrossRef]
- Chou, C.H.; Waller, G.R. Possible allelopathic constituents of Coffea arabica. J. Chem. Ecol. 1980, 6, 643–654. [Google Scholar] [CrossRef]
- Czajkowski, R.; de Boer, W.J.; van der Wolf, J.M. Chemical disinfectants can reduce potato blackleg caused by Dickeya solani. Eur. J. Plant Pathol. 2013, 136, 419–432. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caffeine | COS–Caffeine | COS–Fluorophenoxy-Caffeine | COS–Tetrafluorophenoxy-Caffeine | COS–Bromo-Caffeine | COS–Iodo-Caffeine | Assignment |
|---|---|---|---|---|---|---|
| 1693 | 1708 | 1701 | 1706 | 1701 | 1697 | stretching vibration of conjugated C=O(2) and C=O(6) carbonyl groups |
| 1644 | 1652 | 1646 | 1659 | 1656 | 1659 | C=N and C=C stretching in imidazoles and NH2 bending |
| 1598 | 1598 | 1614 | 1605 | C=N vibration and NH2 bending | ||
| 1546 | 1548 1520 | 1557 1525 | 1556 1519 | 1562 | 1557 1538 | C=N and C=C vibrations |
| 1480 | 1481 | 1504 | 1492 | C–N vibration | ||
| 1455 | 1456 | 1473 1441 | 1455 | 1449 | 1441 | associated with methyl groups |
| 1429 | 1429 1398 | | 1403 | 1404 | C=C and C–H stretching and C=N deformation | |
| 1358 | 1360 | 1380 | 1380 | C–H deformation | ||
1326 | 1335 | 1341 | 1342 1321 | C−N stretching | ||
| 1285 | 1285 1272 | 1286 | 1284 1273 | 1283 | 1276 | C=C and C=O stretching |
| 1239 | 1238 | 1245 1200 | 1215 | 1252 1215 | 1256 1214 | stretching of the >C=O ketonic carbonyl group and C−N bonds |
| 1189 1133 | 1185 1137 | 1152 | 1185 1151 1136 | 1185 1151 1136 | 1185 1151 | C–N vibration in imidazoles and C–O stretching of the alcoholic hydroxyl group in chitosan |
| 1067 | 1067 | 1069 | 1068 | 1067 | 1067 | CH out-of-plane deformation |
| 1024 | 1027 | 1024 | 1031 | 1033 | 1031 | C–C stretching; C–N stretching; >C=O (ketonic) group |
| 972 | 977 946 | 970 | 964 946 | 972 | 974 946 | combination of C–CH3 vibrations and C–H deformation |
846 | 894 807 | 864 847 | 800 | 895 836 | C–C stretching and/or out-of-plane hydrogen deformation of the NH group |
| COS– Caffeine | COS– Caffeine- Imidazole | COS– F-Phenoxy Caffeine | COS– F-Phenoxy Caffeine- Imidazole | COS– 4F-Phenoxycaffeine | COS– 4F-Phenoxycaffeine- Imidazole | COS– Bromo-Caffeine | COS– Bromo-Caffeine- Imidazole | COS– Iodo-Caffeine | COS– Iodo-Caffeine-Imidazole |
|---|---|---|---|---|---|---|---|---|---|
| 1695 | 1698 | 1699 | 1701 | 1710 | 1712 | 1702 | 1705 | 1697 | 1698 |
| 1652 | 1659 | 1646 | 1648 | 1659 | 1663 | 1656 | 1668 | 1659 | 1661 |
| Conjugate Complex | 1st Stage | Weight Loss | 2nd Stage | Weight Loss | 3rd Stage | Weight Loss |
|---|---|---|---|---|---|---|
| COS–caffeine | 230, 340 | 70% | 465, 525 | 28% | >560 | 2% |
| COS–Fluorophenoxy-caffeine | 248, 345 | 54% | 480, 558 | 29% | >560 | 16% |
| COS -Tetrafluorofluorophenoxy-caffeine | 248, 337 | 63% | 500, 560 | 18% | >560 | 19% |
| COS–Bromo-caffeine | 205, 330 | 49% | 455, 533 | 42% | >560 | 8% |
| COS–Iodo-caffeine | 225 | 10% | 497, 515 | 53% | >560 | 37% |
| Conjugate Complex | Energy (kcal mol−1) |
|---|---|
| Caffeine-imidazole | 45.72 |
| Caffeine-imidazole (parallel rings) | 41.24 |
| Fluorophenoxy-caffeine-imidazole | 47.78 |
| Tetrafluorophenoxy-caffeine-imidazole | 55.95 |
| Bromo-caffeine-imidazole | 43.88 |
| Iodo-caffeine-imidazole | 56.06 |
| COS–imidazole | 132.43 |
| COS–caffeine-imidazole | 197.46 |
| Treatment | Effective Concentration (µg mL−1) | D. seriata | D. viticola | N. parvum |
|---|---|---|---|---|
| COS | EC50 | 744 | 554 | 680 |
| EC90 | 1180 | 1139 | 1327 | |
| Caffeine | EC50 | 678 | 303 | 479 |
| EC90 | 1341 | 1352 | 1360 | |
| Fluorophenoxy-caffeine | EC50 | 684 | 2548 | 147 |
| EC90 | 7106 * | 67,913 * | 1863 | |
| Tetrafluorophenoxy-caffeine | EC50 | 11,786 * | 24,746 * | 362 |
| EC90 | 108,037 * | 561,164 * | 1985 | |
| Bromo-caffeine | EC50 | 554 | 292 | 769 |
| EC90 | 767 | 829 | 1379 | |
| Iodo-caffeine | EC50 | 492 | 237 | 1044 |
| EC90 | 968 | 753 | 2792 | |
| Imidazole | EC50 | 1407 | 611 | 1241 |
| EC90 | 2793 | 1190 | 2392 | |
| Theophylline | EC50 | 6947 | 820 | 2771 |
| EC90 | 42,213 * | 2689 | 11,454 * |
| Treatment | Effective Concentration (µg mL−1) | D. seriata | D. viticola | N. parvum |
|---|---|---|---|---|
| COS–caffeine | EC50 | 483 | 331 | 598 |
| EC90 | 727 | 789 | 1267 | |
| COS–Fluorophenoxy-caffeine | EC50 | 472 | 494 | 121 |
| EC90 | 900 | 1205 | 998 | |
| COS–Tetrafluorofluorophenoxy-caffeine | EC50 | 681 | 565 | 517 |
| EC90 | 943 | 960 | 1045 | |
| COS–Bromo-caffeine | EC50 | 539 | 238 | 571 |
| EC90 | 688 | 490 | 1192 | |
| COS–Iodo-caffeine | EC50 | 310 | 240 | 599 |
| EC90 | 471 | 640 | 935 | |
| COS–Imidazole | EC50 | 494 | 206 | 419 |
| EC90 | 910 | 695 | 1052 | |
| COS–Theophylline | EC50 | 729 | 439 | 593 |
| EC90 | 1155 | 804 | 917 | |
| COS–Iodo-caffeine-imidazole | EC50 | 215 | 116 | 163 |
| EC90 | 425 | 267 | 509 | |
| COS–Theophylline-imidazole | EC50 | 659 | 329 | 752 |
| EC90 | 880 | 705 | 1250 |
| Pathogen | COS– Caffeine | COS– Br-caffeine | COS– I-caffeine | COS– F-phenoxy-caffeine | COS– 4F-phenoxy-caffeine | COS– Imidazole | COS– Theophylline |
|---|---|---|---|---|---|---|---|
| D. seriata | 1.73 | 1.35 | 2.26 | 2.25 | 2.48 | 1.82 | 1.99 |
| D. viticola | 1.56 | 1.96 | 1.42 | 1.86 | 2.37 | 1.68 | 1.99 |
| N. parvum | 1.05 | 1.13 | 1.92 | 1.55 | 1.52 | 1.62 | 2.59 |
| D. seriata | D. viticola | N. parvum |
|---|---|---|
| COS–1500 | COS–1500 | COS–1500 |
| COS–F-Phenoxycaffeine–1000 | COS–F-Phenoxycaffeine–1500 | COS–F-Phenoxycaffeine–1500 |
| COS–4F-Phenoxycaffeine–1500 | COS–4F-Phenoxycaffeine–1500 | COS–4F-Phenoxycaffeine–1500 |
| COS–Br-Caffeine–750 | COS–Br-Caffeine–750 | COS–Br-Caffeine–1500 |
| COS–I-Caffeine–500 | COS–I-Caffeine–750 | COS–I-Caffeine–1000 |
| COS–Imidazole–1000 | COS–Imidazole–750 | COS–Imidazole–1500 |
| COS–I-Caffeine-Imidazole–500 | COS–I-Caffeine-Imidazole–375 | COS–I-Caffeine-Imidazole–750 |
| COS–Theophylline–1500 | COS–Theophylline–1000 | COS–Theophylline–1500 |
| COS–Theophylline-Imidazole–1000 | COS–Theophylline-Imidazole–750 | COS–Theophylline-Imidazole–1500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Hernández, E.; Andrés-Juan, C.; Buzón-Durán, L.; Correa-Guimaraes, A.; Martín-Gil, J.; Martín-Ramos, P. Antifungal Activity of Methylxanthines against Grapevine Trunk Diseases. Agronomy 2022, 12, 885. https://doi.org/10.3390/agronomy12040885
Sánchez-Hernández E, Andrés-Juan C, Buzón-Durán L, Correa-Guimaraes A, Martín-Gil J, Martín-Ramos P. Antifungal Activity of Methylxanthines against Grapevine Trunk Diseases. Agronomy. 2022; 12(4):885. https://doi.org/10.3390/agronomy12040885
Chicago/Turabian StyleSánchez-Hernández, Eva, Celia Andrés-Juan, Laura Buzón-Durán, Adriana Correa-Guimaraes, Jesús Martín-Gil, and Pablo Martín-Ramos. 2022. "Antifungal Activity of Methylxanthines against Grapevine Trunk Diseases" Agronomy 12, no. 4: 885. https://doi.org/10.3390/agronomy12040885
APA StyleSánchez-Hernández, E., Andrés-Juan, C., Buzón-Durán, L., Correa-Guimaraes, A., Martín-Gil, J., & Martín-Ramos, P. (2022). Antifungal Activity of Methylxanthines against Grapevine Trunk Diseases. Agronomy, 12(4), 885. https://doi.org/10.3390/agronomy12040885