
Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedure
2.2. Data Collection and Statistical Analysis
3. Results
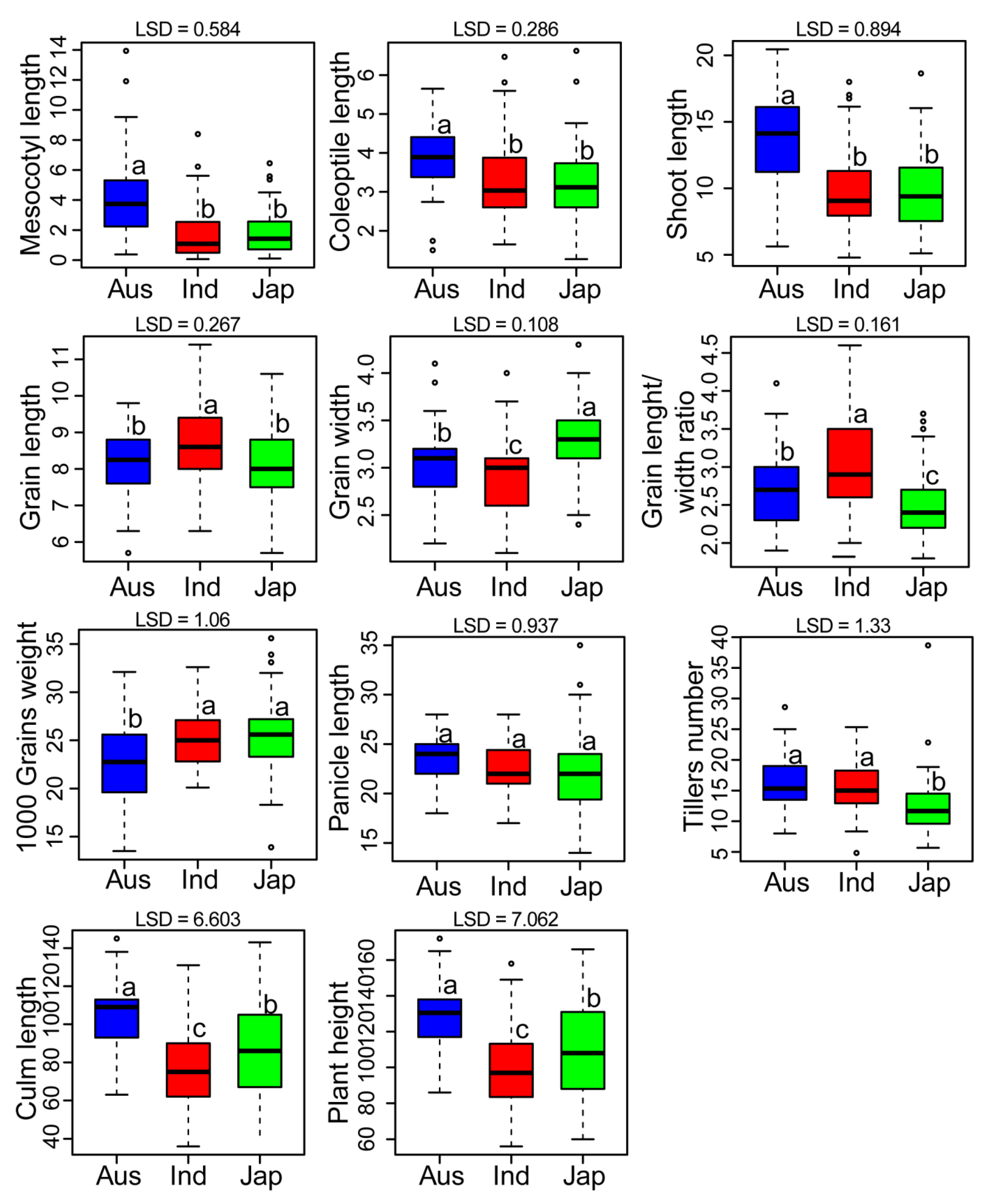
3.1. Mean Performance of Rice Groups and Analysis of Variance (ANOVA)
3.2. Estimation of Heritability in 190 Rice Cultivars
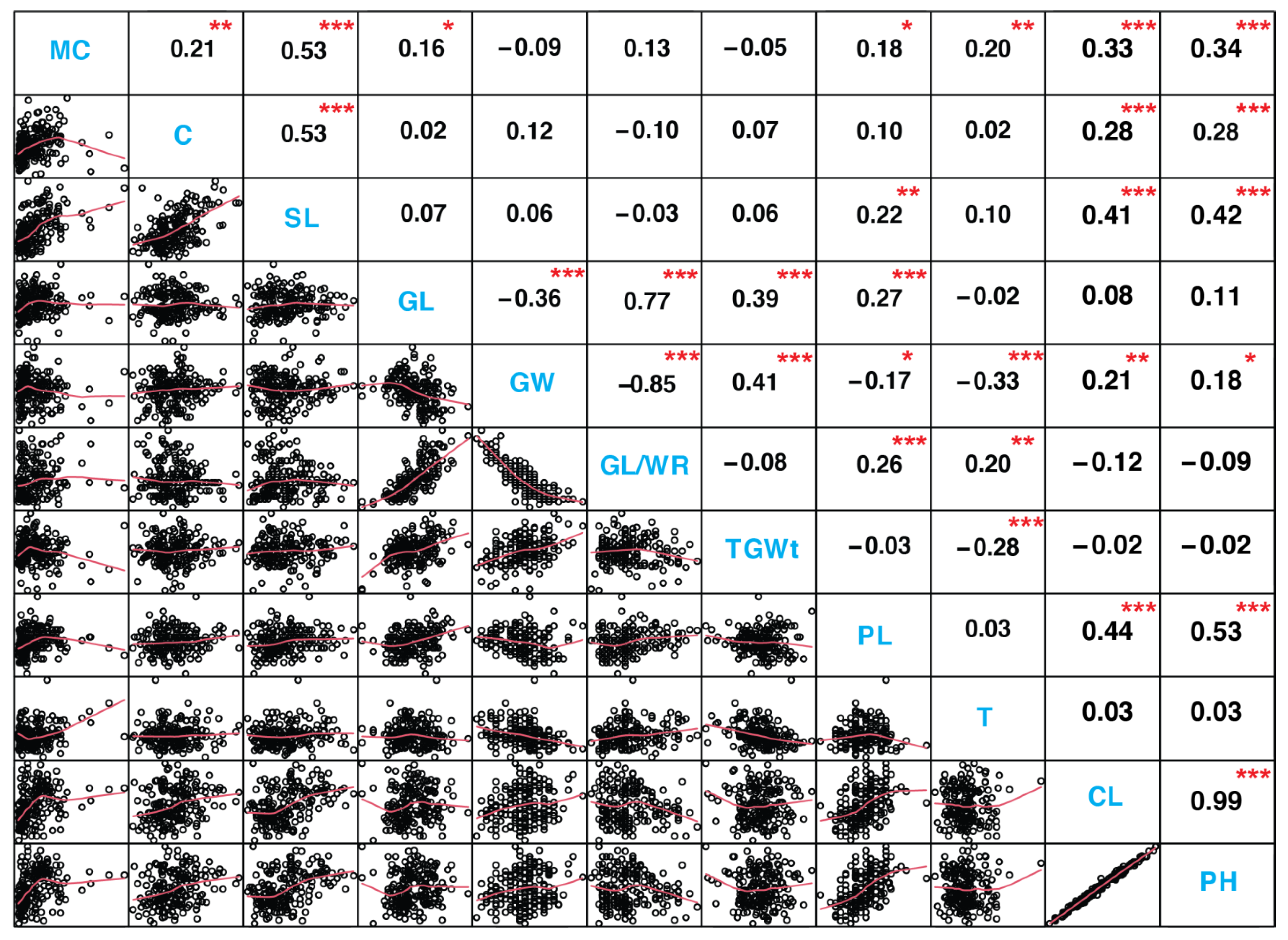
3.3. Relationship between and among the Seedlings and Maturity Traits
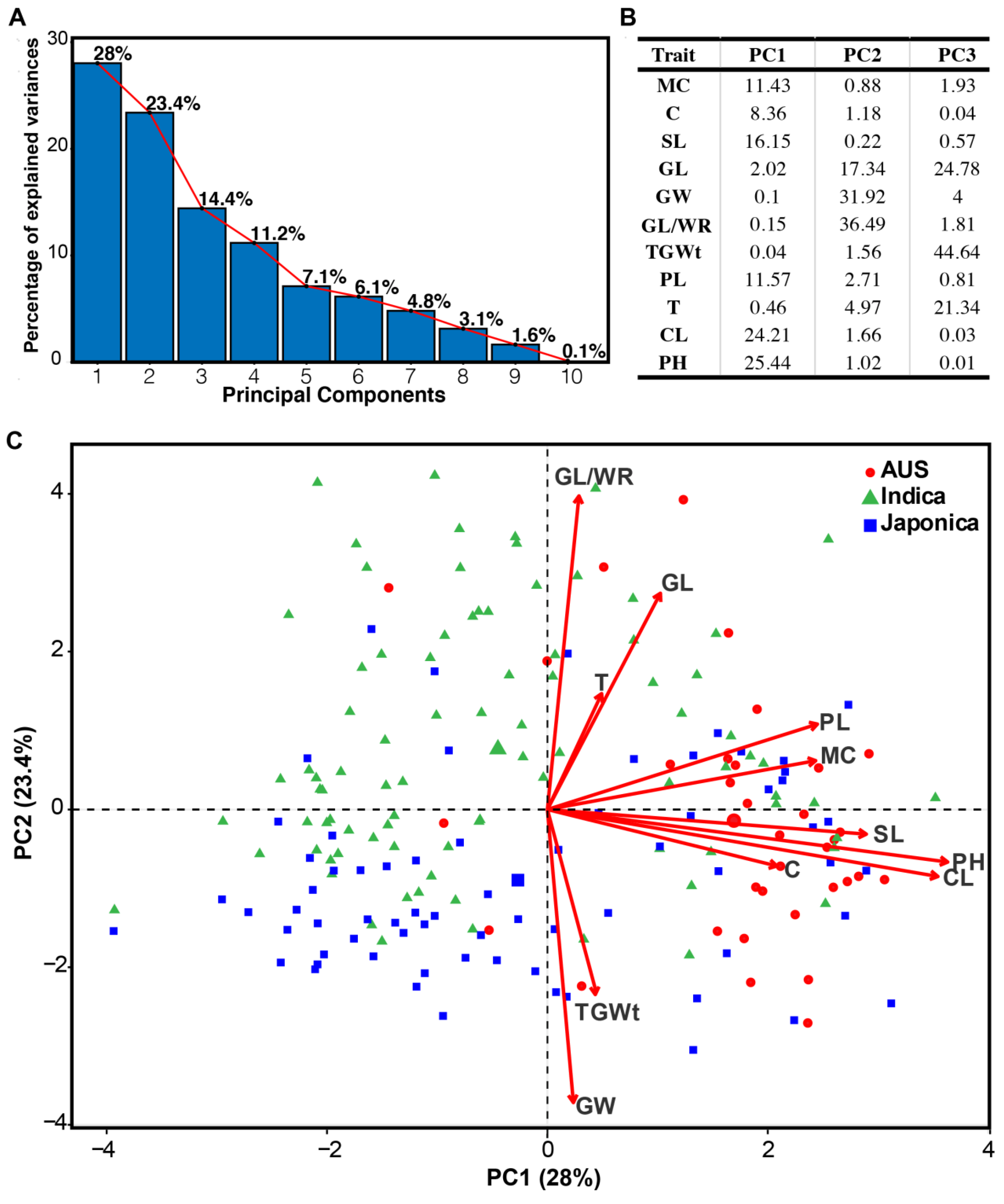
3.4. Principal Components Analysis (PCA)
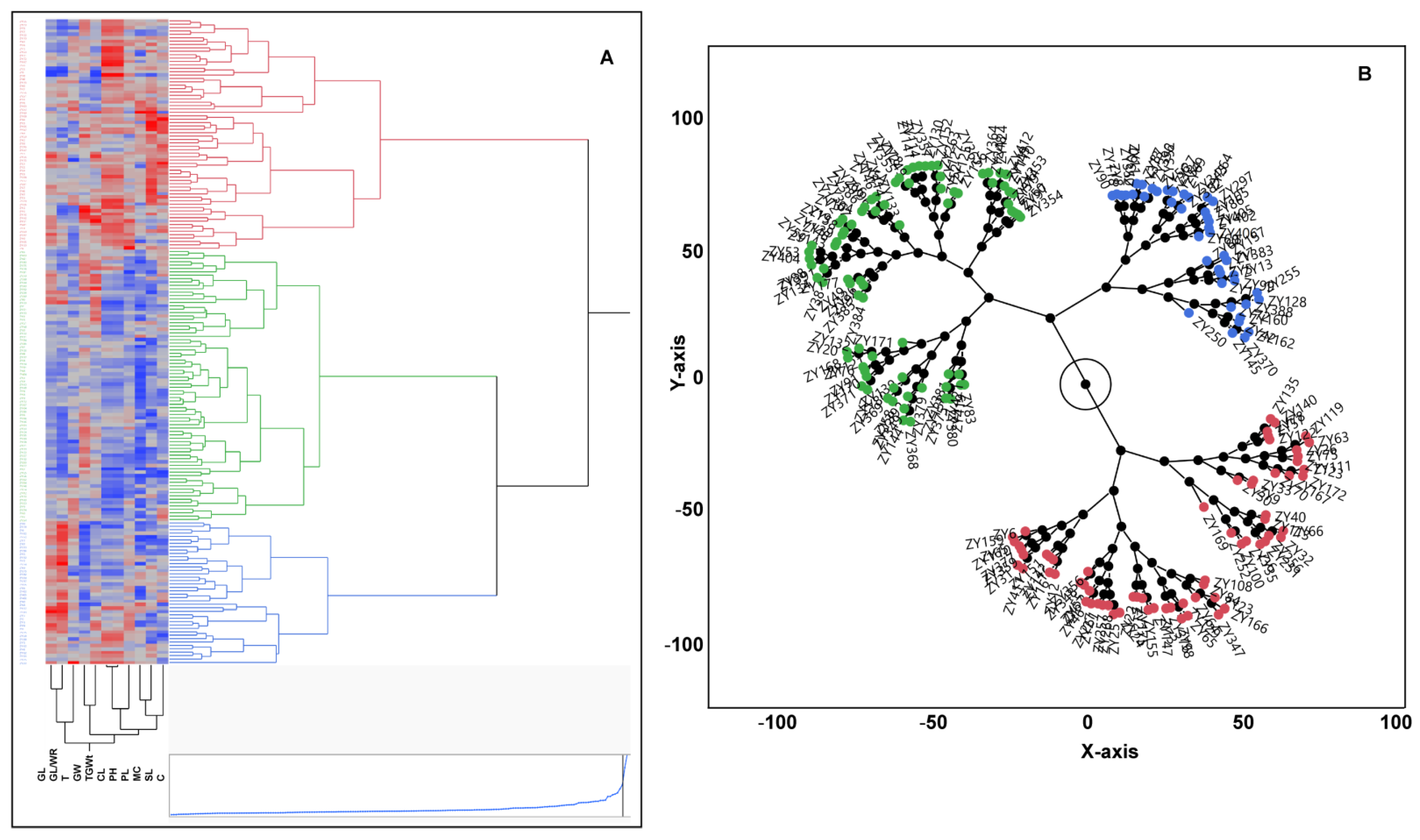
3.5. Estimation of Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, R.; Li, M.; Ashraf, U.; Liu, S.; Zhang, J. Exploring the Relationships between Yield and Yield-Related Traits for Rice Varieties Released in China from 1978 to 2017. Front. Plant Sci. 2019, 10, 00543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.R.; Cai, X.X.; Jin, X. Efficient indica and japonica rice identification based on the InDel molecular method: Its implication in rice breeding and evolutionary research. Prog. Nat. Sci.-Mater. 2009, 19, 1241–1252. [Google Scholar] [CrossRef]
- Vaughan, D.A.; Lu, B.R.; Tomooka, N. The evolving story of rice evolution. Plant Sci. 2008, 174, 394–408. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, K.; Xia, H.; Chen, L.; Chen, K. Comparative proteomic analysis of indica and japonica rice varieties. Genet. Mol. Biol. 2014, 37, 652–661. [Google Scholar] [CrossRef]
- Choi, J.Y.; Platts, A.E.; Fuller, D.Q.; Hsing, Y.I.; Wing, R.A.; Purugganan, M.D. The Rice Paradox: Multiple Origins but Single Domestication in Asian Rice. Mol. Biol. Evol. 2017, 34, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Civan, P.; Ali, S.; Batista-Navarro, R.; Drosou, K.; Ihejieto, C.; Chakraborty, D.; Ray, A.; Gladieux, P.; Brown, T.A. Origin of the Aromatic Group of Cultivated Rice (Oryza sativa L.) Traced to the Indian Subcontinent. Genome Biol. Evol. 2019, 11, 832–843. [Google Scholar] [CrossRef] [Green Version]
- McNally, K.L.; Childs, K.L.; Bohnert, R.; Davidson, R.M.; Zhao, K.; Ulat, V.J.; Zeller, G.; Clark, R.M.; Hoen, D.R.; Bureau, T.E. Genomewide SNP variation reveals relationships among landraces and modern varieties of rice. Proc. Natl. Acad. Sci. USA 2009, 106, 12273–12278. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lawas, L.M.; Malo, R.; Glaubitz, U.; Erban, A.; Mauleon, R.; Heuer, S.; Zuther, E.; Kopka, J.; Hincha, D.K. Metabolic and transcriptomic signatures of rice floral organs reveal sugar starvation as a factor in reproductive failure under heat and drought stress. Plant Cell Environ. 2015, 38, 2171–2192. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [Green Version]
- Henry, A.; Gowda, V.R.; Torres, R.O.; McNally, K.L.; Serraj, R. Variation in root system architecture and drought response in rice (Oryza sativa): Phenotyping of the OryzaSNP panel in rainfed lowland fields. Field Crops Res. 2011, 120, 205–214. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Wang, X.J.; Yuan, L.I.; Hai, X.U.; Chen, W.F. Analysis of Yield and Quality Traits and Their Relationship in Japonica Rice in Northern China. Acta Agron. Sin. 2015, 41, 910–918. [Google Scholar] [CrossRef]
- Wu, J.; Feng, F.; Lian, X.; Teng, X.; Wei, H.; Yu, H.; Xie, W.; Yan, M.; Fan, P.; Li, Y.; et al. Genome-wide Association Study (GWAS) of mesocotyl elongation based on re-sequencing approach in rice. BMC Plant Biol. 2015, 15, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Ladha, J.K. Direct Seeding of Rice: Recent Developments and Future Research Needs. Adv. Agron. 2011, 111, 297–413. [Google Scholar] [CrossRef]
- Wang, W.Q.; Peng, S.B.; Liu, H.Y.; Tao, Y.; Huang, J.L.; Cu, K.H.; Nie, L.X. The possibility of replacing puddled transplanted flooded rice with dry seeded rice in central China: A review. Field Crops Res. 2017, 214, 310–320. [Google Scholar] [CrossRef]
- Liu, H.; He, A.; Jiang, G.; Hussain, S.; Nie, L. Faster leaf senescence after flowering in wet direct-seeded rice was mainly regulated by decrease in cytokinin content as compared with transplanted-flooded rice. Food Energy Secur. 2020, 9, e232. [Google Scholar] [CrossRef]
- Tao, Y.; Chen, Q.; Peng, S.B.; Wang, W.Q.; Nie, L.X. Lower global warming potential and higher yield of wet direct-seeded rice in Central China. Agron. Sustain. Dev. 2016, 36, 24. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Tang, S.Q.; Hu, P.S.; Louis, A.; Jiao, G.A.; Tang, J. Analysis on Factors Affecting Seedling Establishment in Rice. Rice Sci. 2007, 14, 27–32. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhu, D.F.; Xiong, H.; Chen, H.Z.; Xiang, J.; Lin, X.P. Development and transition of rice planting in China. Agric. Sci. Technol. 2012, 13, 1270–1276. [Google Scholar]
- Ismail, A.M.; Johnson, D.E.; Ella, E.S.; Vergara, G.V.; Baltazar, A.M. Adaptation to flooding during emergence and seedling growth in rice and weeds, and implications for crop establishment. AoB Plants 2012, 2012, pls019. [Google Scholar] [CrossRef]
- Lee, H.S.; Sasaki, K.; Kang, J.W.; Sato, T.; Song, W.Y.; Ahn, S.N. Mesocotyl Elongation is Essential for Seedling Emergence Under Deep-Seeding Condition in Rice. Rice 2017, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Barsa, S.M.A.; Wahid, A. Priming of field-sown rice seed enhances germination, seedling establishment, allometry and yield. Plant Growth Regul. 2006, 49, 285–294. [Google Scholar] [CrossRef]
- Patil, S.; Zafar, S.A.; Uzair, M.; Zhao, J.F.; Fang, J.J.; Li, X.Y. An Improved Mesocotyl Elongation Assay for the Rapid Identification and Characterization of Strigolactone-Related Rice Mutants. Agronomy 2019, 9, 208. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.H.; Lu, X.; Liu, H.Y.; Zhao, Q.Z.; Ye, G.Y. Mesocotyl elongation, an essential trait for dry-seeded rice (Oryza sativa L.): A review of physiological and genetic basis. Planta 2020, 251, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.N.; Johnson, D.E.; Sivaprasad, B.; Ladha, J.K.; Mortimer, A.M. Weed management in direct-seeded rice. Advances in Agronomy 2007, 93, 153–255. [Google Scholar] [CrossRef]
- Dang, X.; Thi, T.G.T.; Dong, G.; Wang, H.; Edzesi, W.M.; Hong, D. Genetic diversity and association mapping of seed vigor in rice (Oryza sativa L.). Planta 2014, 239, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Sasaki, K.; Higashitani, A.; Ahn, S.N.; Sato, T. Mapping and characterization of quantitative trait loci for mesocotyl elongation in rice (Oryza sativa L.). Rice 2012, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Zhang, G.; Wu, M.; Cao, L.; Chen, S. Genetic analysis of mesocotyl elongation in rice (Oryza sativa L. subsp.japonica). Acta Agron. Sin. 2006, 32, 249–252. [Google Scholar]
- Panda, S.; Majhi, P.K.; Anandan, A.; Mahender, A.; Veludandi, S.; Bastia, D.; Guttala, S.B.; Singh, S.K.; Saha, S.; Ali, J. Proofing Direct-Seeded Rice with Better Root Plasticity and Architecture. Int. J. Mol. Sci. 2021, 22, 6058. [Google Scholar] [CrossRef]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajatasereekul, S.; O’toole, J.C. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowland: 3. Plant factors contributing to drought resistance. Field Crops Res. 2002, 73, 181–200. [Google Scholar] [CrossRef]
- Jiang, Q.; Wang, W.; Chen, Q.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Response of first flood irrigation timing after rice dry-direct-seeding: Productivity and greenhouse gas emissions in Central China. Agric. Water Manag. 2016, 177, 241–247. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Meng, Y.; Liu, H.; Liu, C.; Ye, G. Rapid identification of QTL for mesocotyl length in rice through combining QTL-seq and genome-wide association analysis. Front. Genet. 2021, 12, 713446. [Google Scholar] [CrossRef] [PubMed]
- Matusmoto, T.; Yamada, K.; Yoshizawa, Y.; Keimei, O.H. Comparison of Effect of Brassinosteroid and Gibberellin Biosynthesis Inhibitors on Growth of Rice Seedlings. Rice Sci. 2016, 23, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhao, W.; Jiang, C.; Wang, X.; Xiong, H.; Todorovska, E.G.; Yin, Z.; Chen, Y.; Wang, X.; Xie, J. Genetic architecture and candidate genes for deep-sowing tolerance in rice revealed by non-syn GWAS. Front. Plant Sci. 2018, 9, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Zhang, M.; Niu, X.; Wang, C.; Xu, Q.; Feng, Y.; Wang, S.; Yuan, X.; Yu, H.; Wang, Y. Uncovering novel loci for mesocotyl elongation and shoot length in indica rice through genome-wide association mapping. Planta 2016, 243, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Zhang, Q.; Wang, C.-C.; Liu, Z.-X.; Jiang, Y.-J.; Zhai, L.-Y.; Zheng, T.-Q.; Xu, J.-L.; Li, Z.-K. Genetic dissection of seedling vigour in a diverse panel from the 3000 Rice (Oryza sativa L.) Genome Project. Sci. Rep. 2019, 9, 4804. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Ren, M.; Huang, M.; Qiu, H.; Chun, Y.; Li, L.; Kumar, A.; Fang, J.; Zhao, J.; He, H.; Li, X. Genome-Wide Association Study of the Genetic Basis of Effective Tiller Number in Rice. Rice 2021, 14, 1–13. [Google Scholar] [CrossRef]
- Wang, C.C.; Yu, H.; Huang, J.; Wang, W.S.; Faruquee, M.; Zhang, F.; Zhao, X.Q.; Fu, B.Y.; Chen, K.; Zhang, H.L.; et al. Towards a deeper haplotype mining of complex traits in rice with RFGB v2.0. Plant Biotechnol. J. 2020, 18, 14–16. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.H.; Zhang, L.Y.; Wang, J.K. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Hajiaqatabar, A.; Kiani, G.; Kazemitabar, S.K.; Alavi, M. Correlation and Path Coefficient Analysis for Yield and Yield Components in F2 Segregating Populations of Rice. Jordan J. Agric. Sci. 2016, 12, 749–754. [Google Scholar] [CrossRef]
- Huang, R.; Jiang, L.; Zheng, J.; Wang, T.; Wang, H.; Huang, Y.; Hong, Z. Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci. 2013, 18, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.L.; Mei, H.W.; Liu, H.Y.; Yu, X.Q.; Zou, G.H.; Luo, L.J. Sensitivities of rice grain yield and other panicle characters to late-stage drought stress revealed by phenotypic correlation and QTL analysis. Mol. Breed. 2010, 25, 603–613. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J. The Plant Architecture of Rice (Oryza sativa). Plant Mol. Biol. 2005, 59, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Mun, H.H.; Chang, T.T.; Beachell, H.M. The inheritance of culm length, panicle length, duration to heading and bacterial leaf blight reaction in a rice cross: Sigadis × Taichung (Native)1. Jpn. J. Breed. 2008, 18, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Mgonja, M.A.; Ladeinde, T.A.O.; Aken’Ova, M.E. Genetic analysis of mesocotyl length and its relationship with other agronomic characters in rice (Oryza sativa L.). Euphytica 1993, 72, 189–195. [Google Scholar] [CrossRef]
- Redofia, E.D.; Mackill, D.J. Genetic Variation for Seedling Vigor Traits in Rice. Crop Sci. 1996, 36, 285–290. [Google Scholar]
- Hour, A.-l.; Hsieh, W.-H.; Chang, S.-H.; Wu, Y.-P.; Chin, H.-S.; Lin, Y.-R. Genetic diversity of landraces and improved varieties of rice (Oryza sativa L.) in Taiwan. Rice 2020, 13, 82. [Google Scholar] [CrossRef]
- Simon, A.; Yuri, S.; Hironobu, S.; Kenji, I. Genotypic variation in coleoptile or mesocotyl lengths of upland rice (Oryza sativa L.) and seedling emergence in deep sowing. Afr. J. Agric. Res. 2012, 7, 6239–6248. [Google Scholar] [CrossRef] [Green Version]
- Nam-Jin, C. Elongation habit of mesocotyls and coleoptiles in weedy rice with high emergence ability in direct-seeding on dry paddy fields. Crop Pasture Sci. 2010, 61, 911–917. [Google Scholar]
- Kim, S.L.; Chung, Y.S.; Silva, R.R.; Ji, H.; Lee, H.; Choi, I.; Kim, N.; Lee, E.; Baek, J.; Lee, G.S.; et al. The opening of phenome-assisted selection era in the early seedling stage. Sci. Rep. 2019, 9, 9948. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Carpel size, grain filling, and morphology determine individual grain weight in wheat. J. Exp. Bot. 2015, 66, 6715–6730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhan, J.; Li, J.; Lu, X.; Liu, J.; Wang, Y.; Zhao, Q.; Ye, G. Genome-wide association study (GWAS) for mesocotyl elongation in rice (Oryza sativa L.) under multiple culture conditions. Genes 2020, 11, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, B.; Khan, A.S.; Ali, Z. Genetic variation in wheat germplasm for salinity tolerance atseedling stage: Improved statistical inference. Turk. J. Agric. For. 2015, 39, 182–192. [Google Scholar] [CrossRef]
- Adjah, K.L.; Abe, A.; Adetimirin, V.O.; Asante, M.D. Genetic variability, heritability and correlations for milling and grain appearance qualities in some accessions of rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2020, 26, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Noorka, I.R.; Amarah, B.; Saeed, R.; Teixeira da Silva, J.; Ashraf, E. Estimation of heterosis in wheat (Triticum aestivum L.) under contrasting water regimes. Int. J. Plant Breed 2013, 7, 55–60. [Google Scholar]
- Dilday, R.H.; Mgonja, M.A.; Amonsilpa, S.A.; Collins, F.C.; Wells, B.R. Plant height vs mesocotyl and coleoptile elongation in rice: Linkage or pleiotropism? Crop Sci. 1990, 30, 815–818. [Google Scholar] [CrossRef]
- Zhang, J.; Tong, T.; Potcho, P.M.; Huang, S.; Ma, L.; Tang, X. Nitrogen Effects on Yield, Quality and Physiological Characteristics of Giant Rice. Agronomy 2020, 10, 1816. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: Preparing for the 21st century. Genome 1999, 42, 646–655. [Google Scholar] [CrossRef]
- Jun, M.; Wenbo, M.; Yanhua, T.; Jianchang, Y.; Kaida, Z.; Qingsen, Z. The culm lodging resistance of heavy panicle type of rice. Zuo Wu Xue Bao 2004, 30, 143–148. [Google Scholar]
- Islam, M.S.; Peng, S.; Visperas, R.M.; Ereful, N.; Bhuiya, M.S.U.; Julfiquar, A. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crops Res. 2007, 101, 240–248. [Google Scholar] [CrossRef]
- Yano, K.; Morinaka, Y.; Wang, F.; Huang, P.; Takehara, S.; Hirai, T.; Ito, A.; Koketsu, E.; Kawamura, M.; Kotake, K.; et al. GWAS with principal component analysis identifies a gene comprehensively controlling rice architecture. Proc. Natl. Acad. Sci. USA 2019, 116, 21262–21267. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Zhang, S.; Abdelghany, A.M.; Shaibu, A.S.; Feng, Y.; Li, Y.; Tian, Y.; Hong, H.; Li, B.; Sun, J. Seed isoflavone profiling of 1168 soybean accessions from major growing ecoregions in China. Food Res. Int. 2020, 130, 108957. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Zhang, X.; Luo, J.; Liu, H.; Xiao, Y. Metabolomic analysis reveals differences in evolution between maize and rice. Plant J. 2020, 103, 1710–1722. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L. Progress in super-hybrid rice breeding. Crop J. 2017, 5, 100–102. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, C.; Lin, J.; Liu, J.; Liu, B.; Wang, J.; Huang, A.; Li, H.; Zhao, T. OsMPH1 regulates plant height and improves grain yield in rice. PLoS ONE 2017, 12, e0180825. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Overall Population | Aus Group | Indica Group | Japonica Group | ANOVA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | CV | Mean | Range | CV | Mean | Range | CV | Mean | Range | CV | G | h2 | |
| C | 3.34 | 1.27–6.62 | 29.28 | 3.85 | 1.5–5.65 | 24.55 | 3.24 | 1.65–6.47 | 30.37 | 3.21 | 1.27–6.62 | 28.54 | *** | 99 |
| MC | 2.24 | 0.06–13.93 | 95.46 | 4.17 | 0.37–13.93 | 74.87 | 1.79 | 0.06–8.39 | 94.00 | 1.85 | 0.10–6.45 | 78.77 | *** | 97 |
| SL | 10.51 | 4.79–20.45 | 31.54 | 13.64 | 5.63–20.45 | 25.46 | 9.87 | 4.79–18 | 29.03 | 9.77 | 5.11–18.64 | 29.48 | *** | 98 |
| GL | 8.41 | 5.7–11.4 | 10.97 | 8.22 | 5.7–9.8 | 10.88 | 8.69 | 6.3–11.4 | 10.13 | 8.15 | 5.7–10.6 | 11.06 | NS | - |
| GW | 3.08 | 2.1–4.3 | 13.19 | 3.06 | 2.2–4.1 | 13.21 | 2.9 | 2.1–4.0 | 12.57 | 3.31 | 2.4–4.3 | 10.09 | *** | - |
| GL/WR | 2.8 | 1.8–4.6 | 21.31 | 2.74 | 1.9–4.1 | 19.84 | 3.07 | 2–4.6 | 19.94 | 2.49 | 1.8–3.7 | 17.00 | *** | - |
| TGWt | 24.8 | 13.5–35.6 | 14.86 | 22.53 | 13.5–32.1 | 19.34 | 25.10 | 20.1–32.6 | 12.02 | 25.53 | 13.9–35.6 | 14.55 | ** | - |
| PL | 22.66 | 14–35 | 13.84 | 23.44 | 18–28 | 10.77 | 22.52 | 17–28 | 11.48 | 22.46 | 14–35 | 17.49 | NS | - |
| T | 14.63 | 4.83–38.66 | 32.13 | 15.95 | 8–28.6 | 29.39 | 15.79 | 4.83–25.33 | 26.86 | 12.52 | 5.66–38.66 | 36.55 | *** | - |
| CL | 85.77 | 36–145 | 28.06 | 105.29 | 63–145 | 18.92 | 77.86 | 36–131 | 27.81 | 86.14 | 42–143 | 27.33 | NS | - |
| PH | 108.44 | 56–172 | 23.6 | 128.73 | 86–172 | 16.38 | 100.38 | 56–158 | 22.64 | 108.60 | 60–166 | 23.71 | NS | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uzair, M.; Patil, S.B.; Zhang, H.; Kumar, A.; Mkumbwa, H.; Zafar, S.A.; Chun, Y.; Fang, J.; Zhao, J.; Khan, M.R.; et al. Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice. Agronomy 2022, 12, 975. https://doi.org/10.3390/agronomy12040975
Uzair M, Patil SB, Zhang H, Kumar A, Mkumbwa H, Zafar SA, Chun Y, Fang J, Zhao J, Khan MR, et al. Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice. Agronomy. 2022; 12(4):975. https://doi.org/10.3390/agronomy12040975
Chicago/Turabian StyleUzair, Muhammad, Suyash B. Patil, Hongrui Zhang, Ashmit Kumar, Humphrey Mkumbwa, Syed Adeel Zafar, Yan Chun, Jingjing Fang, Jinfeng Zhao, Muhammad Ramzan Khan, and et al. 2022. "Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice" Agronomy 12, no. 4: 975. https://doi.org/10.3390/agronomy12040975
APA StyleUzair, M., Patil, S. B., Zhang, H., Kumar, A., Mkumbwa, H., Zafar, S. A., Chun, Y., Fang, J., Zhao, J., Khan, M. R., Yuan, S., & Li, X. (2022). Screening Direct Seeding-Related Traits by Using an Improved Mesocotyl Elongation Assay and Association between Seedling and Maturity Traits in Rice. Agronomy, 12(4), 975. https://doi.org/10.3390/agronomy12040975