
Comparative Analysis of Canopy Cooling in Wheat under High Temperature and Drought Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
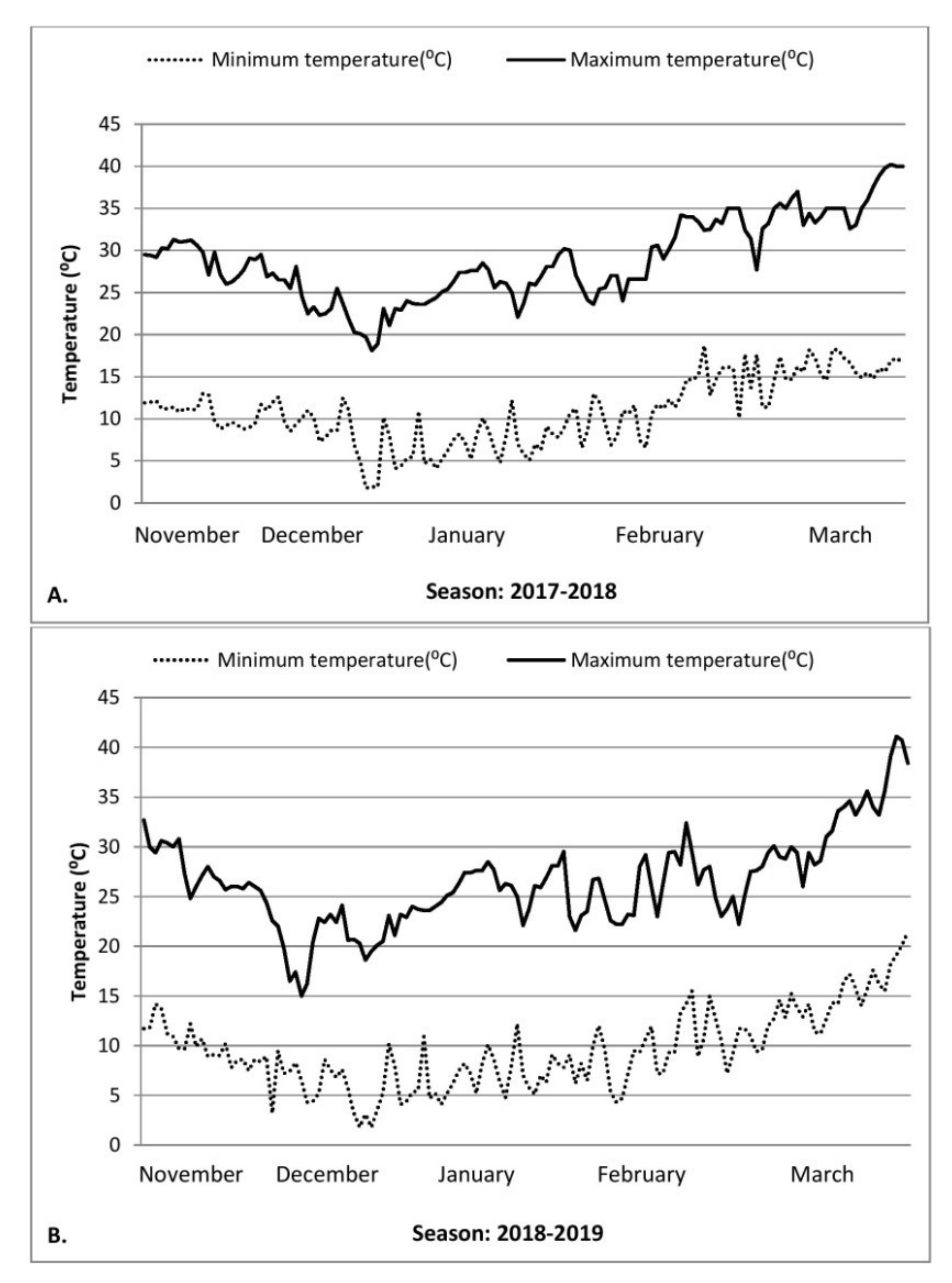
2.2. Environment Monitoring and Plant Measurements
2.3. Statistical Analysis
3. Results
3.1. Effect of Drought and High Temperature
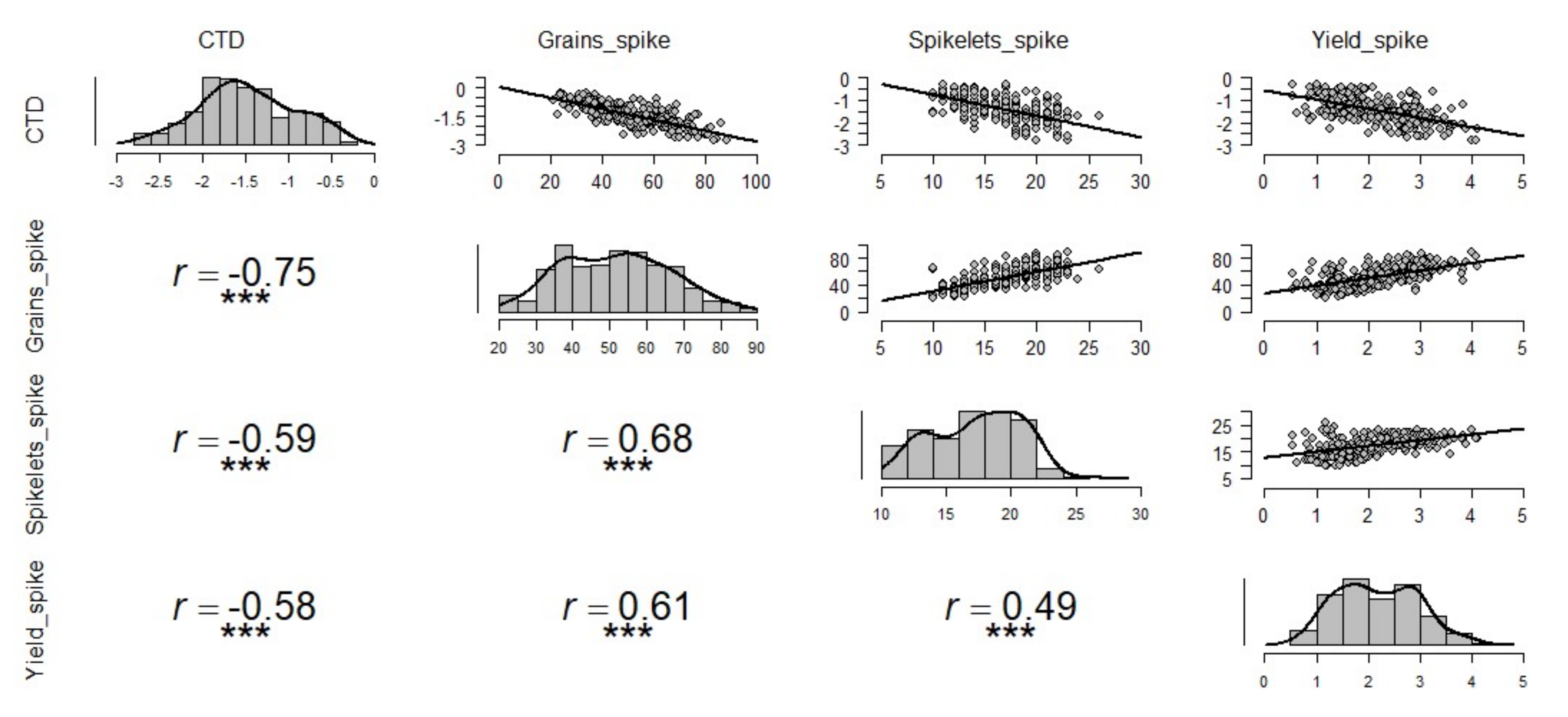
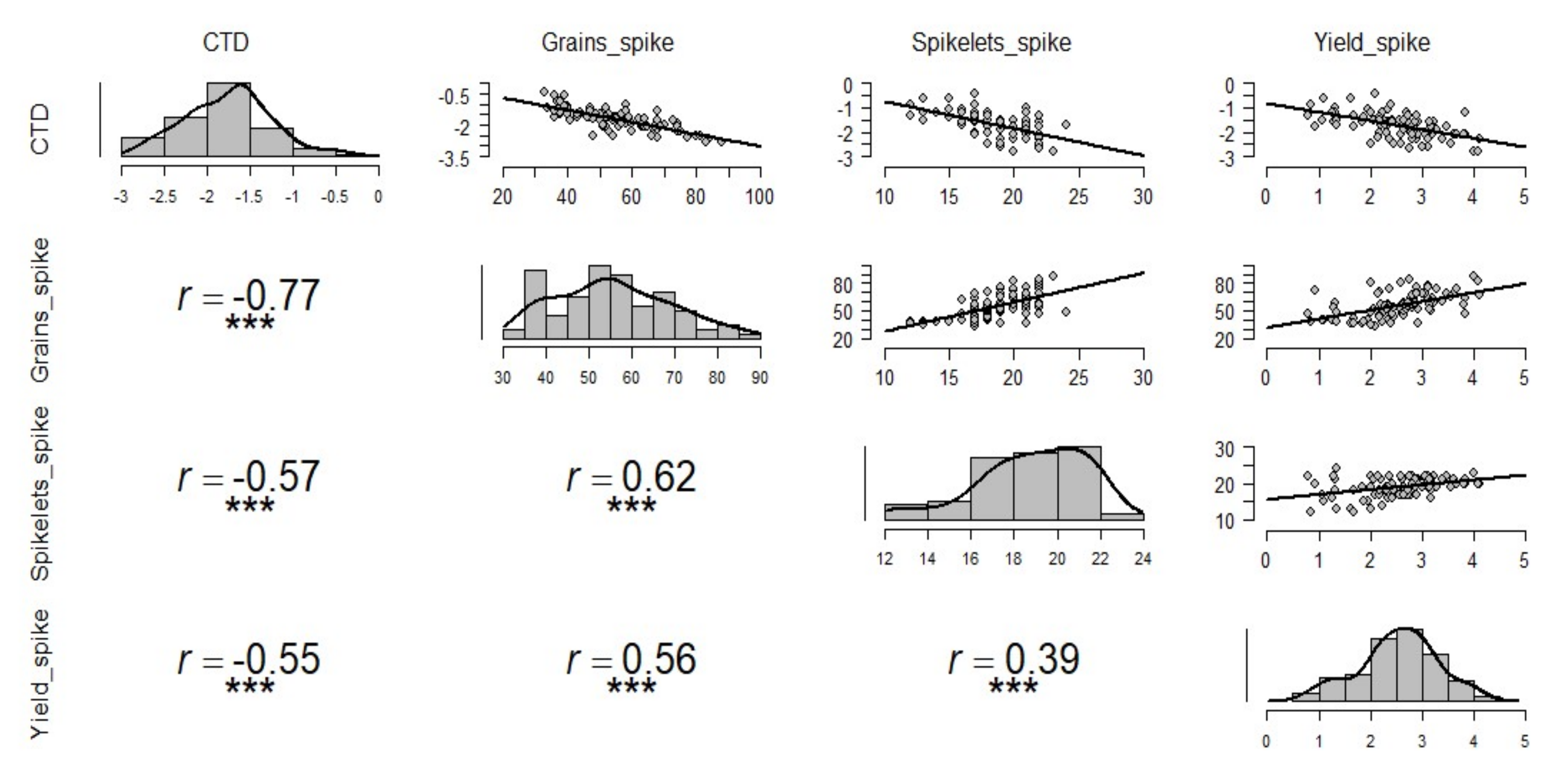
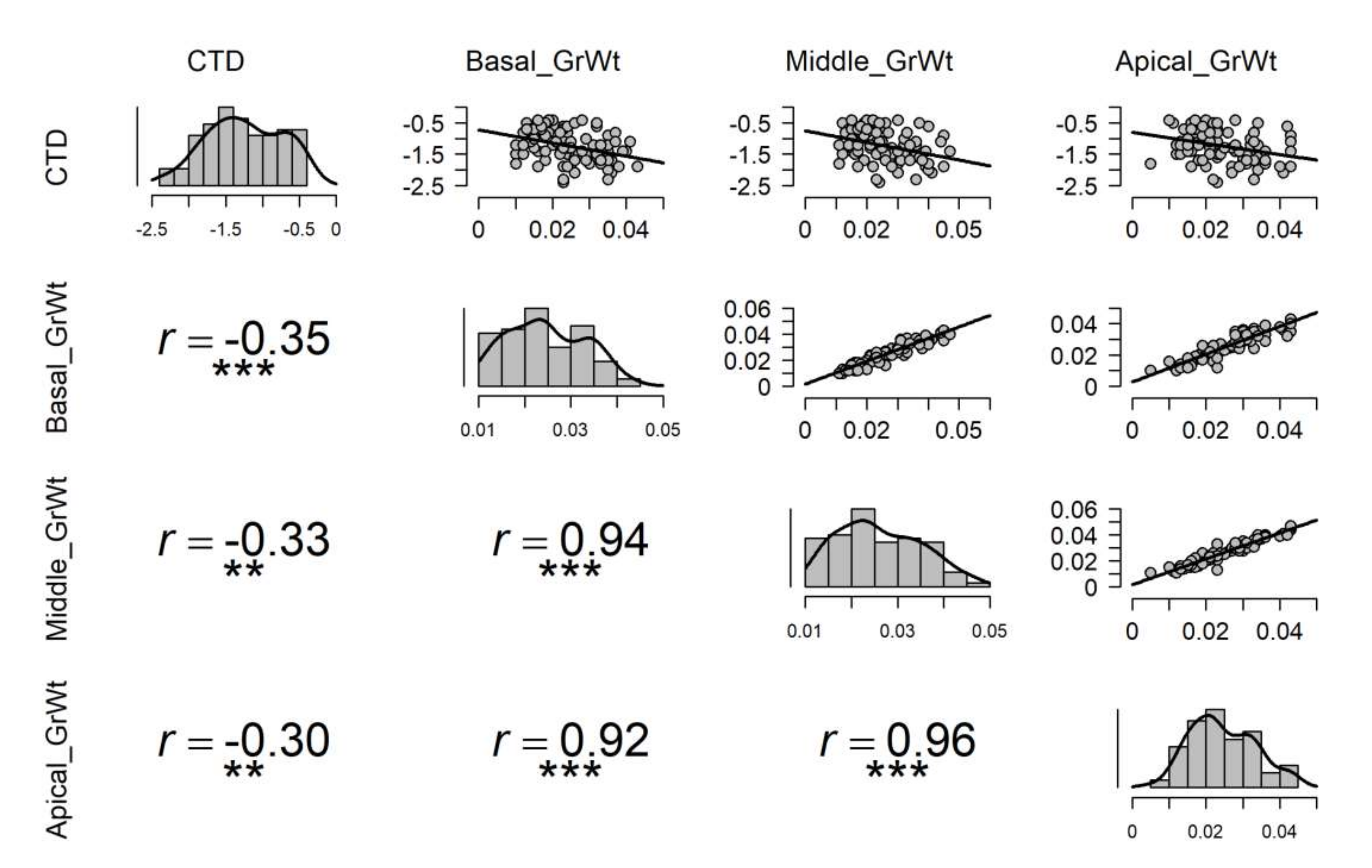
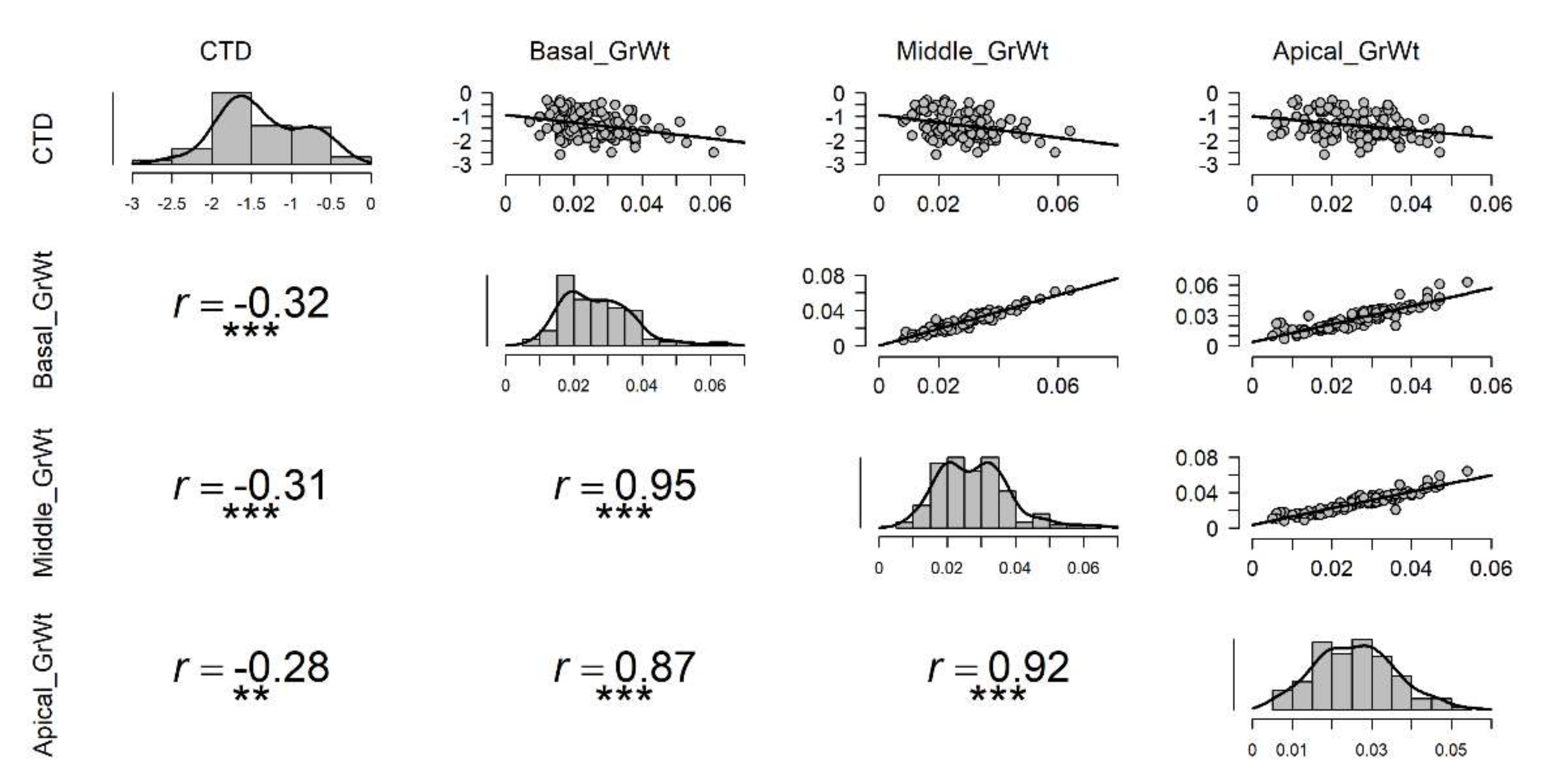
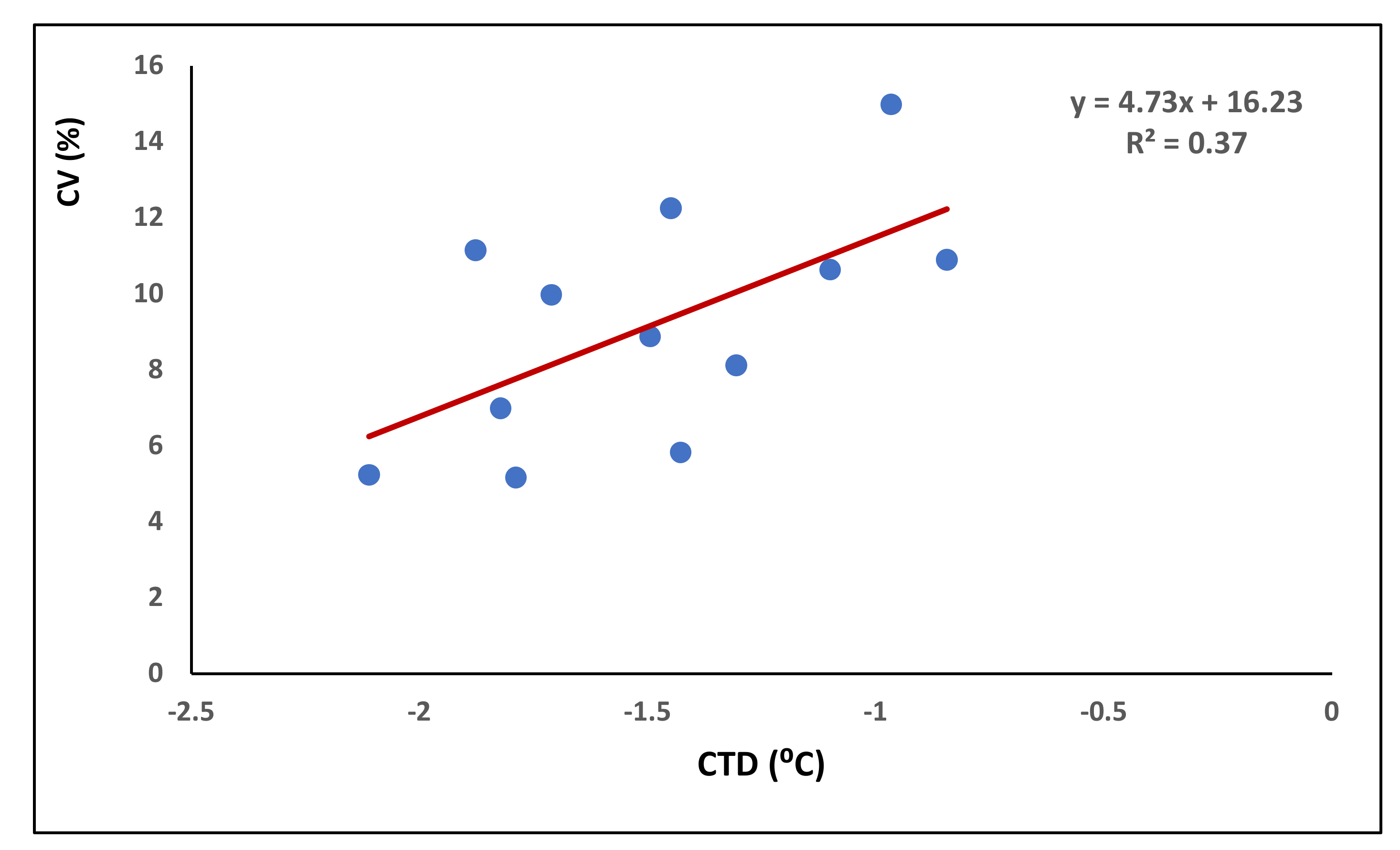
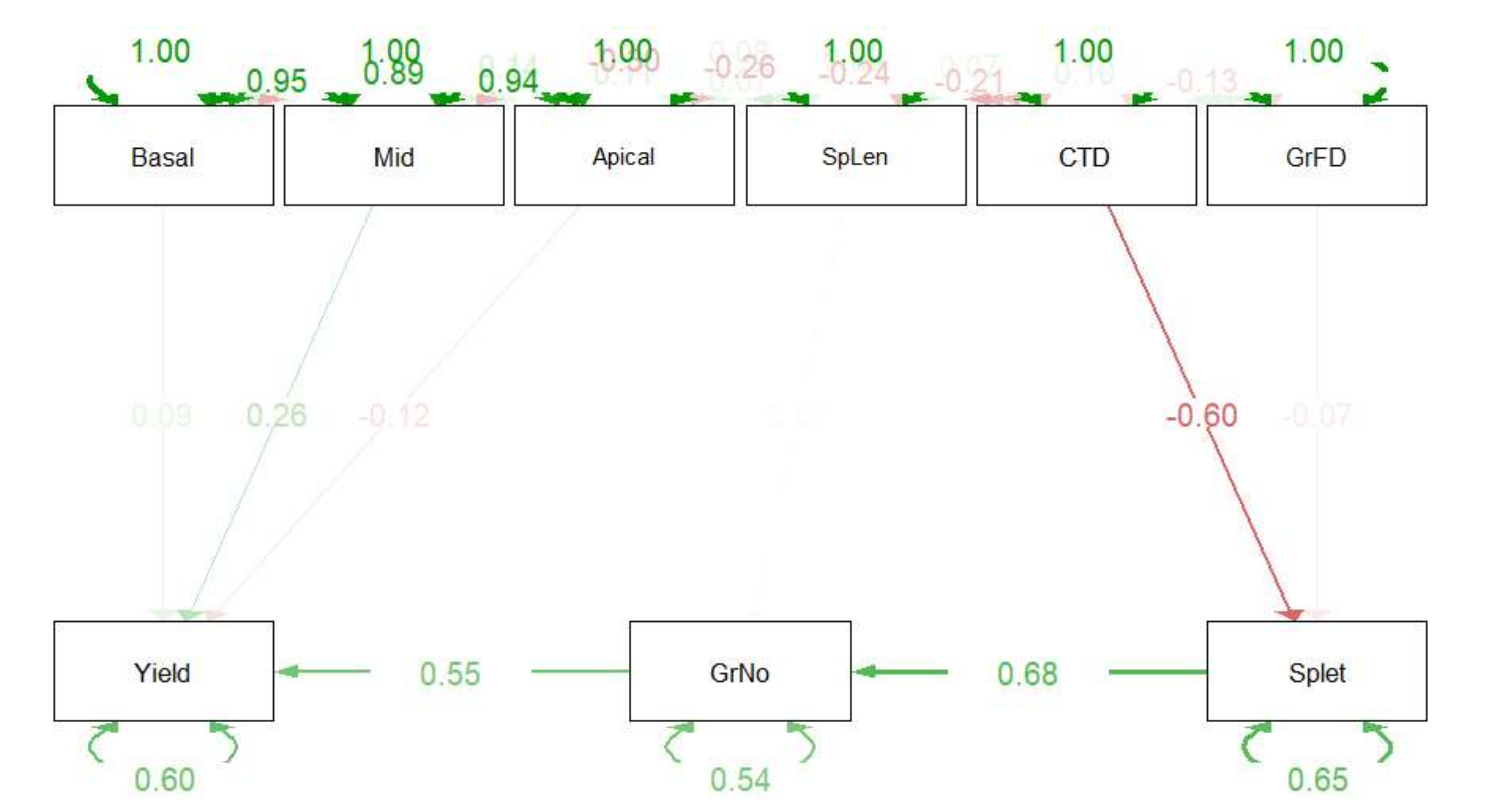
3.2. The Influence of CTD on Grain Yield and Yield Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grote, U.; Fasse, A.; Nguyen, T.; Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 317. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought Stress in Wheat during Flowering and Grain-filling Periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Sofield, I.; Evans, L.; Cook, M.; Wardlaw, I. Factors Influencing the Rate and Duration of Grain Filling in Wheat. Funct. Plant Biol. 1977, 4, 785. [Google Scholar] [CrossRef]
- Stone, P.; Nicolas, M. Effect of Timing of Heat Stress during Grain Filling on Two Wheat Varieties Differing in Heat Tolerance. I. Grain Growth. Funct. Plant Biol. 1995, 22, 927. [Google Scholar] [CrossRef]
- Dorofeev, V.F.; Rudenko, M.I.; Filatenko, A.A.; Baras, J.; Segnalova, J.; Lemann, H. (Compiler) Тhе Intеrnаtionаl Сomесon List of Dеsсriрtors for thе Gеnus Тritiсum L.; VIR: Russia, Leningrad, 1984. (In Russian)
- Konopatskaia, I.; Vavilova, V.; Blinov, A.; Goncharov, N.P. Spike Morphology Genes in Wheat Species (Triticum L.). Proc. Latv. Acad. Sci. Sect. B. Nat. Exact Appl. Sci. 2016, 70, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Baillot, N.; Girousse, C.; Allard, V.; Piquet-Pissaloux, A.; Le Gouis, J. Different grain-filling rates explain grain-weight differences along the wheat ear. PLoS ONE 2018, 13, e0209597. [Google Scholar] [CrossRef]
- Hughes, N.; Askew, K.; Scotson, C.P.; Williams, K.; Sauze, C.; Corke, F.; Doonan, J.H.; Nibau, C. Non-destructive, high-content analysis of wheat grain traits using X-ray micro computed tomography. Plant Methods 2017, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Calderini, D.F.; Ortiz-Monasterio, I. Grain position affects grain macronutrient and micronutrient concentrations in wheat. Crop Sci. 2003, 43, 141–151. [Google Scholar] [CrossRef]
- Liu, H.Z.; Wang, Y.H.; Wang, E.X.; Zhang, P.G.; Chen, D.P.; Liu, J.D. Genotypic and spike positional difference in grain phytase activity, phytate, inorganicphosphorus, iron, and zinc contents in wheat (Triticum aestivum L.). J. Cereal Sci. 2006, 44, 212–219. [Google Scholar] [CrossRef]
- Lizana, C.X.; Riegel, R.; Gomez, D.L.; Herrera, J.; Isla, A.; Mcqueen-Mason, J.S.; Calderini, F.D. Expansin expression is associated with grain size dynamics in wheat (Triticum aestivum L.). J. Exp. Bot. 2010, 61, 1147–1157. [Google Scholar] [CrossRef]
- Xie, Q.; Mayes, S.; Sparkes, D. Carpel size, grain filling, and morphology determine individual grain weight in wheat. J. Exp. Bot. 2015, 66, 6715–6730. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.P.; Balota, M.; Delgado, M.I.B.; Amani, I.; Fischer, R.A. Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions. Aust. J. Plant Physiol. 1994, 21, 717–730. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Larque Saavedra, A. Wheat yield progress is associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Ninanya, J.; Ramírez, D.A.; Rinza, J.; Silva-Díaz, C.; Cervantes, M.; García, J.; Quiroz, R. Canopy Temperature as a Key Physiological Trait to Improve Yield Prediction under Water Restrictions in Potato. Agronomy 2021, 11, 1436. [Google Scholar] [CrossRef]
- Gutierrez-Rodriguez, M.; Reynolds, M.P.; Escalante-Estrada, J.A.; Rodriguez-Gonzalez, M.T. Association between canopy reflectance indices and yield and physiological traits in bread wheat under drought and well-irrigated conditions. Aust. J. Agric. Res. 2004, 55, 1139–1147. [Google Scholar] [CrossRef]
- Rajala, A.; Hakala, K.; Mäkelä, P.; Peltonen-Sainio, P. Drought Effect on Grain Number and Grain Weight at Spike and Spikelet Level in Six-Row Spring Barley. J. Agron. Crop Sci. 2010, 197, 103–112. [Google Scholar] [CrossRef]
- Rehman, H.U.; Tariq, A.; Ashraf, I.; Ahmed, M.; Muscolo, A.; Basra, S.M.A.; Reynolds, M. Evaluation of Physiological and Morphological Traits for Improving Spring Wheat Adaptation to Terminal Heat Stress. Plants 2021, 10, 455. [Google Scholar] [CrossRef]
- Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R.; Hakim, M.A.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S.; et al. Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices. Agronomy 2021, 11, 1792. [Google Scholar] [CrossRef]
- Ullah, S.; Trethowan, R.; Bramley, H. The Physiological Basis of Improved Heat Tolerance in Selected Emmer-Derived Hexaploid Wheat Genotypes. Front. Plant Sci. 2021, 12, 739246. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of Bread Wheat Genotypes for Drought Tolerance Using Phenotypic and Proline Analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [Green Version]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of diverse Wheat (Triticum aestivum L.) Genotypes Varying in Sensitivity to Heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichthardt, C.; Chen, T.W.; Stahl, A.; Stutzel, H. Co-evolution of sink and source in the recent breeding history of winter wheat in Germany. Front. Plant Sci. 2020, 10, 1771. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, J.L.; Mandal, D.K.; Mandal, C. Agro-Ecological Sub-Regions Map of India (1:4.4 m); National Bureau of Soil Survey and Land Use Planning, Indian Council of Agricultural Research (ICAR): Nagpur, India, 1996.
- Arora, A.; Kundu, S.; Dilbaghi, N.; Sharma, I.; Tiwari, R. Population structure and genetic diversity among Indian wheat varieties using microsatellite (SSR) markers. Aust. J. Crop Sci. 2014, 8, 1281–1289. [Google Scholar]
- Sharma, P.; Sharma, R.B. Molecular characterization of wheat germplasm for leaf and stem rust resistance genes using linked SSR markers. Can. J. Plant Breed. 2014, 2, 15–27. [Google Scholar]
- Malik, B.S.; Sethi, A.P.; Tewari, V. Cultivar development program. Ann. Wheat Newslet. 2003, 49, 51. [Google Scholar]
- Malik, R.; Tiwari, R.; Arora, A.; Kumar, P.; Sheoran, S.; Sharma, S.; Singh, R.; Tiwari, V.; Sharma, I. Genotypic characterization of elite Indian wheat genotypes using molecular markers and their pedigree analysis. Aust. J. Crop Sci. 2013, 7, 561–567. [Google Scholar] [CrossRef]
- Singh, T.; Dhaliwal, I.; Khosla, G. Breeder seed production of wheat at PAU: A way to quality seed availability. Agric. Res. J. 2020, 57, 284–296. [Google Scholar] [CrossRef]
- Agrawal, R.K. Development of Improved Varieties. Twenty Five Years of Co-Ordinated Wheat Research 1961–1986; Tandon, J.P., Sethi, A.P., Eds.; Wheat Project Directorate, IARI: New Delhi, India, 1986; pp. 34–39. [Google Scholar]
- Jain, K.B.L. Personal Communication; Indian Agricultural Research Institute: New Delhi, India, 1994. [Google Scholar]
- Ram, S.; Jain, N.; Dawar, V.; Singh, R.P.; Shoran, J. Acid-PAGE gliadin pattern of Indian wheats (Triticum aestivum L.) representing different environments and periods. Crop Sci. 2005, 45, 1256–1263. [Google Scholar] [CrossRef]
- The Australian Winter Cereals Collection Pedigree Database; Agricultural Research Centre: Tamworth, Australia, 1994.
- Kundu, S.; Singh, C.; Shoran, J.; Singh, S.S. An update on released wheat varieties and registered genetic stocks (Triticum L.). In Technical bulletin No.13; Directorate of Wheat Research: Karnal, India, 2010; p. 34. [Google Scholar]
- Pietragalla, J. Chapter 1. Canopy temperature. In Physiological Breeding II: A Field Guide to Wheat Phenotyping; Pask, A.J.D., Pietragalla, J., Mullan, D.M., Reynolds, M.P., Eds.; CIMMYT: Méx, Mexico, 2012. [Google Scholar]
- Fischer, K.S.; Maurer, R. Drought resistance in spring wheat genotypes. I. Grain yield responses. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- R team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2013, p. 201. Available online: https://www.R-project.org/ (accessed on 3 September 2021).
- Fox, J. Structural Equation Modeling With the sem Package in R. Struct. Equ. Modeling. 2006, 13, 465–486. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.K.; Ibrahim, A.M.H.; Rudd, J.; Hays, D.; Malla, S.; Baker, J. Increasing hard winter wheat yield potential via synthetic hexaploid wheat: II. Heritability and combining ability of yield and its components. Crop Sci. 2013, 53, 67–73. [Google Scholar] [CrossRef]
- Mahpara, S.; Hussain, S.T.; Iqbal, J.; Noorka, I.R.; Salman, S. 12. Analysis of generation means for some metric plant traits in two wheat (Triticum aestivum L.) hybrids. Pure Appl. Biol. 2018, 7, 93–102. [Google Scholar] [CrossRef]
- Mahpara, S.; Rehmani, M.I.A.; Hussain, S.; Iqbal, J.; Qureshi, M.K.; Shehzad, M.A.; Dar, J.S. Heterosis for some physio-morphological plant traits in spring wheat crosses. Pure Appl. Biol. 2017, 6, 1103–1110. [Google Scholar] [CrossRef]
- Ferrante, A.; Savin, R.; Slafer, G.A. Floret development and grain setting differences between modern durum wheats under contrasting nitrogen availability. J. Exp. Bot. 2013, 64, 169–184. [Google Scholar] [CrossRef]
- Guo, Z.; Schnurbusch, T. Variation of floret fertility in hexaploid wheat revealed by tiller removal. J. Exp. Bot. 2015, 66, 5945–5958. [Google Scholar] [CrossRef] [Green Version]
- Gautam, A.; Sai Prasad, S.V.; Jajoo, A.; Ambati, D. Canopy Temperature as a Selection Parameter for Grain Yield and Its Components in Durum Wheat Under Terminal Heat Stress in Late Sown Conditions. Agric. Res. 2015, 4, 238–244. [Google Scholar] [CrossRef]
- Rutkoski, J.; Poland, J.; Mondal, S.; Autrique, E.; Pérez, L.G.; Crossa, J.; Reynolds, M.; Singh, R. Canopy Temperature and Vegetation Indices from High-Throughput Phenotyping Improve Accuracy of Pedigree and Genomic Selection for Grain Yield in Wheat. G3 Genes Genom. Genet. 2016, 6, 2799–2808. [Google Scholar] [CrossRef] [Green Version]
- Thapa, S.; Jessup, K.E.; Pradhan, G.P.; Rudd, J.C.; Liu, S.; Mahan, J.R.; Devkota, R.N.; Baker, J.A.; Xue, Q. Canopy temperature depression at grain filling correlates to winter wheat yield in the U.S. Southern High Plains. Field Crops Res. 2018, 217, 11–19. [Google Scholar] [CrossRef]
- Blum, A. Plant Breeding for Stress Environments; CRC Press: Boca Raton, FL, USA, 1988; p. 223. [Google Scholar]
- Sharma, D.L.; Anderson, W.K. Small grain screenings in wheat: Interactions of genotypes with season, site, and management practices. Aust. J. Agric. Res. 2004, 55, 797. [Google Scholar] [CrossRef]
- Nuttall, J.G.; O’Leary, G.J.; Panozzo, J.F.; Walker, C.K.; Barlow, K.M.; Fitzgerald, G. Models of grain quality in wheat—A review. Field Crops Res. 2017, 202, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Philipp, N.; Weichert, H.; Bohra, U.; Weschke, W.; Schulthess, A.W.; Weber, H. Grain numberand grain yield distribution along the spike remainstable despite breeding for high yield in winter wheat. PLoS ONE 2018, 13, e0205452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, M.; Foulkes, M.J.; Slafer, G.A.; Berry, P.; Parry, M.A.; Snape, J.W.; Angus, W.J. Raising yield potential in wheat. J. Exp. Bot. 2009, 60, 1899–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, P.; Rawson, H. The Weights of Individual Grains of the Wheat Ear in Relation to Their Growth Potential, the Supply of Assimilate and Interaction between Grains. Funct. Plant Biol. 1978, 5, 61–72. [Google Scholar] [CrossRef]
- Wardlaw, I.F. Temperature control of translocation. In Mechanism of Regulation of Plant Growth; Bielske, R.L., Ferguson, A.R., Cresswell, M.M., Eds.; Bulletin of Royal Society of New Zealand: Wellington, New Zealand, 1974; pp. 533–538. [Google Scholar]
- Tashiro, T.; Wardlaw, I.F. The Effect of High Temperature at Different Stages of Ripening on GrainSet, Grain Weight and Grain Dimensions in the Semi-dwarf Wheat “Banks”. Ann. Bot. 1990, 65, 51–61. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, D.; Alqudah, A.M.; Röder, M.S.; Ganal, M.W.; Schnurbusch, T. Genome-wide association analyses of 54 traits identified multiple loci for the determination of floret fertility in wheat. New Phytol. 2017, 214, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Freeman, K.W.; Raun, W.R.; Johnson, G.V.; Mullen, R.W.; Stone, M.L.; Solie, J.B. Late-season prediction of wheat grain yield and grain protein. Commun. Soil Sci. Plant Anal. 2007, 34, 1837–1852. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, M.; Shah, M.K.N. Performance of wheat genotypes for morpho-physiological traits using multivariate analysis under terminal heat stress. Pak. J. Bot. 2019, 52, 1981–1988. [Google Scholar] [CrossRef]
- Ozturk, I. Effect of Canopy Temperature at Different Growth Stage on Yield Component in Bread Wheat (Triticum aestivum L.) Genotypes under Rainfed Condition. Int. J. Innov. Approaches Agric. Res. 2020, 4, 136–146. [Google Scholar] [CrossRef]
- Parihar, R.; Agrawal, A.P.; Burman, M.; Minz, M.G. Relationship between grain yield and other yield attributing characters in wheat under terminal heat stress. J. Pharmacog. Phytochem. 2018, 7, 2114–2117. [Google Scholar]
- Zečević, V.; Knežević, D.; Bošković, J.; Mićanović, D.; Dimitrijević, B. Genetic and phenotypic variability of number of spikelets per Spike in winter wheat. Kragujev. J. Sci. 2009, 31, 85–90. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Genotypes | Pedigree | Source of Genotype * | Spike Shape |
|---|---|---|---|---|
| 1 | AKAW 4627 | WH-147/SUNSTAR * 6/C-80-1 [25] | PDKV, Akola | Compact |
| 2 | HALNA | HD-1982/K-816 [26] | CSUAT, Kanpur | Normal |
| 3 | HD 2643 | VEE S/HD 2407/HD 2329 [27] | IARI, New Delhi | Spelt |
| 4 | HD 2967 | ALD/COC//URES/HD2160m/HD 2278 [28] | IARI, New Delhi | Spelt |
| 5 | HD 2987 | HI1011/HD2348//MENDOS//IWP72/DL153-2 [25] | IARI, New Delhi | Normal |
| 6 | HPBW01 | T.DICOCCON, CI9309/AE.SQUARROSA (409)//MUTUS/3/2 * MUTUS [29] | PAU, Ludhiana | Normal |
| 7 | HUW 234 | HUW-12 * 2/CPAN-1666//HUW-12 [30] | BHU, Varanasi | Compact |
| 8 | MP 4010 | CAM50123-3M-Y-2M-1Y-2M-Y-2M-2Y-OM-OMR/S [25] | JNKVV Jabalpur | Spelt |
| 9 | PBW 175 | HD2160/4/JN/GAGE/JN/KAL/3/PV18/C273 [31] | PAU, Ludhiana | Normal |
| 10 | PBW 396 | CNO67/MFD//MON”S”/3/SERI [32] | PAU, Ludhiana | Spelt |
| 11 | Raj 3765 | HD-2402/VL-639; HP-1633/HP-1776 [32] | Durgapura, Jaipur | Normal |
| 12 | Raj 4037 | DL-788-2/RAJ-3717 [32] | Durgapura, Jaipur | Normal |
| 13 | Raj 4079 | UP2363/WH595 [32] | Durgapura, Jaipur | Normal |
| 14 | Sunstar | CONDOR, AUS/4/2 * WW-15/3/STEINWEDEL/YAROSLAV-EMMER//LA-PREVISION [33] | Australia | Compact |
| 15 | WH 730 | CPAN 2092/Improved Lok-1 [34] | HAU, Hisar | Normal |
| Source of Variation | Df | Grains Spike−1 | Yield Spike−1 (g) | Spikelets Spike−1 | GFD | CTD (°C) |
|---|---|---|---|---|---|---|
| Year | 1 | 69.51 | 0.074 | 18.148 ** | 2569.96 *** | 0.0623 |
| Treatment | 2 | 1150.36 *** | 9.6804 *** | 168.344 *** | 267.27 *** | 5.1925 *** |
| Genotype | 14 | 2395.64 *** | 4.9404 *** | 123.856 *** | 66.93 *** | 2.2761 *** |
| Year:Treatment | 2 | 96 | 0.0511 | 18.226 *** | 181.07 *** | 0.2663 |
| Year:Genotype | 14 | 28.1 | 0.0865 | 5.505 *** | 56.24 *** | 0.2592 |
| Treatment:Genotype | 28 | 329.39 *** | 1.045 *** | 18.829 *** | 8.68 * | 0.441 *** |
| Year:Treatment:Genotype | 28 | 29.1 | 0.0783 | 6.535 *** | 17.09 *** | 0.2035 |
| Residuals | 180 | 56.47 | 0.2363 | 1.685 | 5.18 | 0.1628 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, V.; Rane, J.; Nankar, A.N. Comparative Analysis of Canopy Cooling in Wheat under High Temperature and Drought Stress. Agronomy 2022, 12, 978. https://doi.org/10.3390/agronomy12040978
Thakur V, Rane J, Nankar AN. Comparative Analysis of Canopy Cooling in Wheat under High Temperature and Drought Stress. Agronomy. 2022; 12(4):978. https://doi.org/10.3390/agronomy12040978
Chicago/Turabian StyleThakur, Vidisha, Jagadish Rane, and Amol N. Nankar. 2022. "Comparative Analysis of Canopy Cooling in Wheat under High Temperature and Drought Stress" Agronomy 12, no. 4: 978. https://doi.org/10.3390/agronomy12040978
APA StyleThakur, V., Rane, J., & Nankar, A. N. (2022). Comparative Analysis of Canopy Cooling in Wheat under High Temperature and Drought Stress. Agronomy, 12(4), 978. https://doi.org/10.3390/agronomy12040978