
Heat-Stress-Mitigating Effects of a Protein-Hydrolysate-Based Biostimulant Are Linked to Changes in Protease, DHN, and HSP Gene Expression in Maize

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Experiment
2.2. Hydroponic Experiments
2.2.1. Biostimulant Effects on Germination and Growth of Hydroponically Grown Maize Plants
2.2.2. Heat Shock Applied on Hydroponically Grown Plants Pre-Treated with Biostimulant
2.3. Determination of Photosynthetic Pigments, Oxidative Stress Markers, and Metabolites
2.4. Enzyme Assays
2.5. RT-PCR Analysis
2.6. Statistical Analyses
3. Results
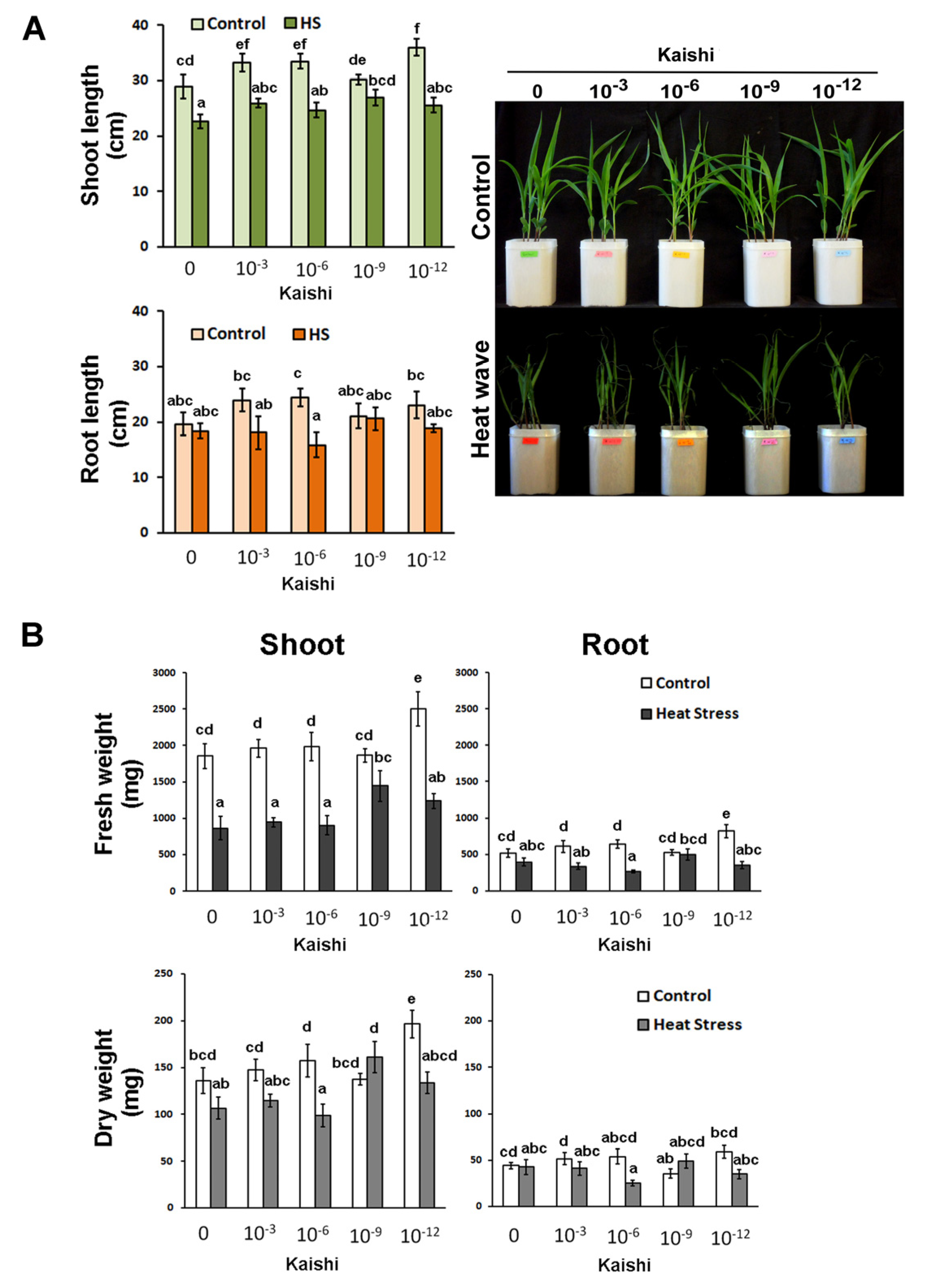
3.1. Effects of the Biostimulant on Soil-Grown Maize Plants Subjected to Simulated Heat Wave Treatment
3.1.1. Growth Parameters
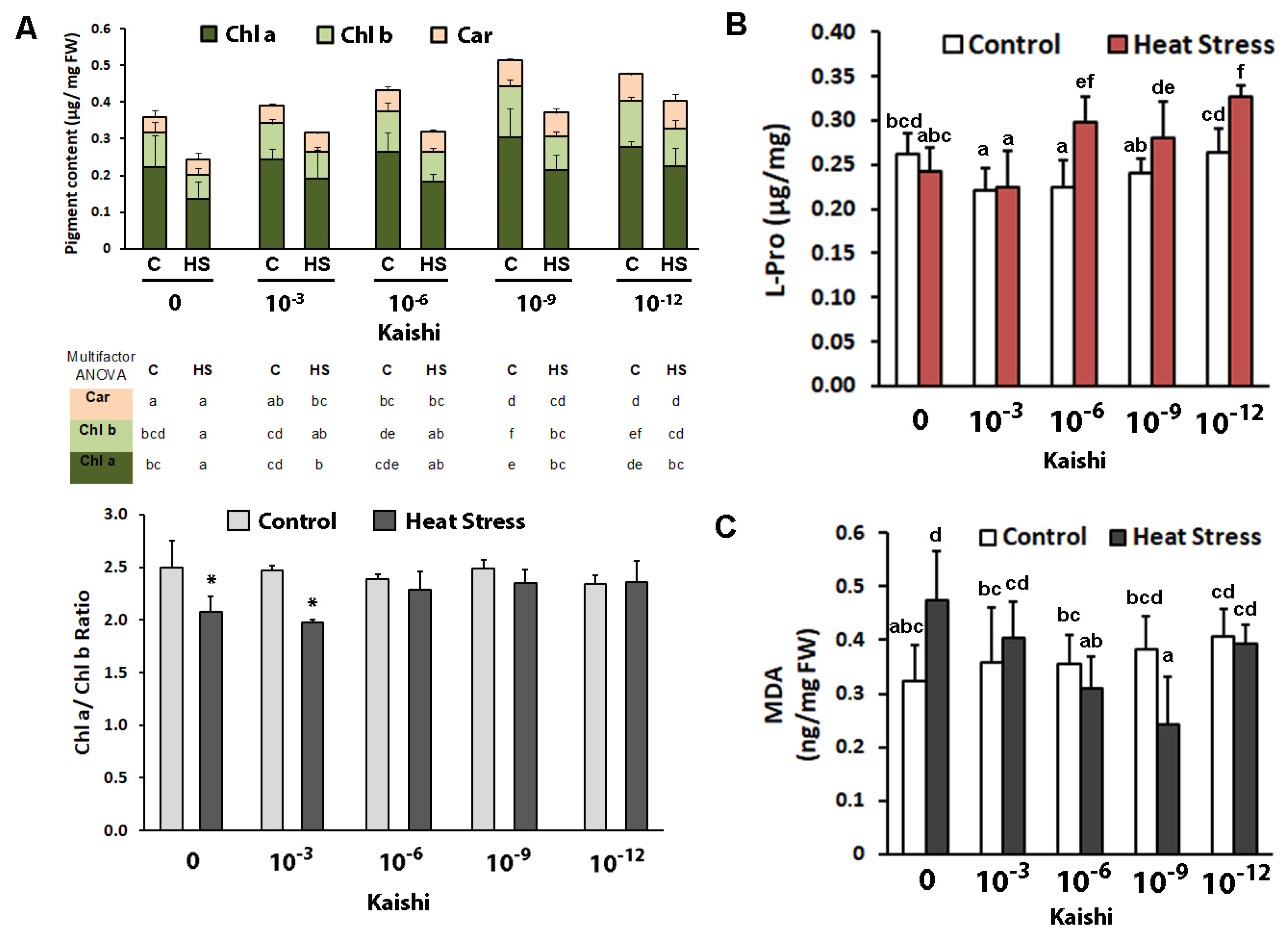
3.1.2. Biochemical Analyses
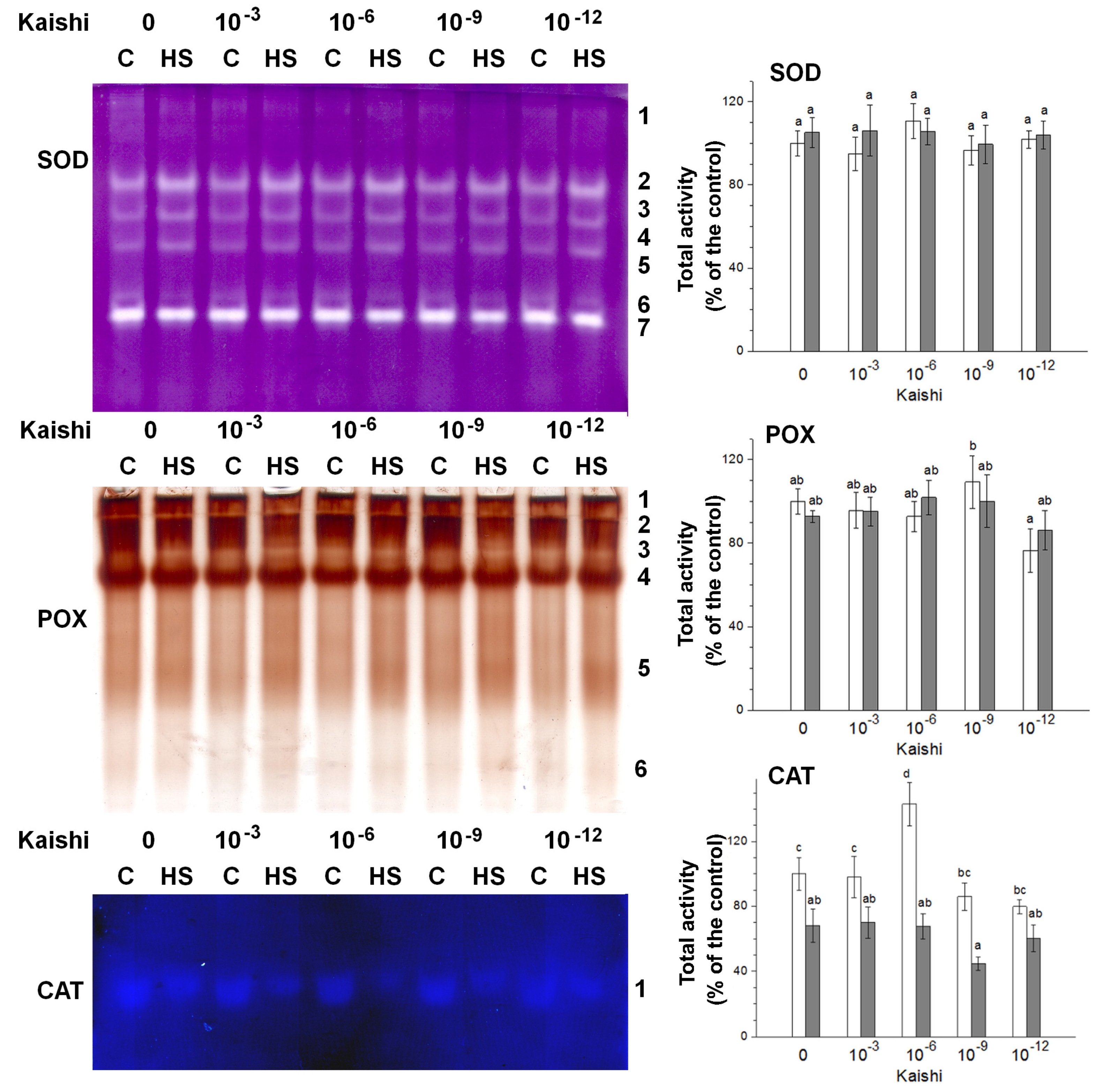
3.1.3. In-Gel Staining of Antioxidative Enzyme Activities
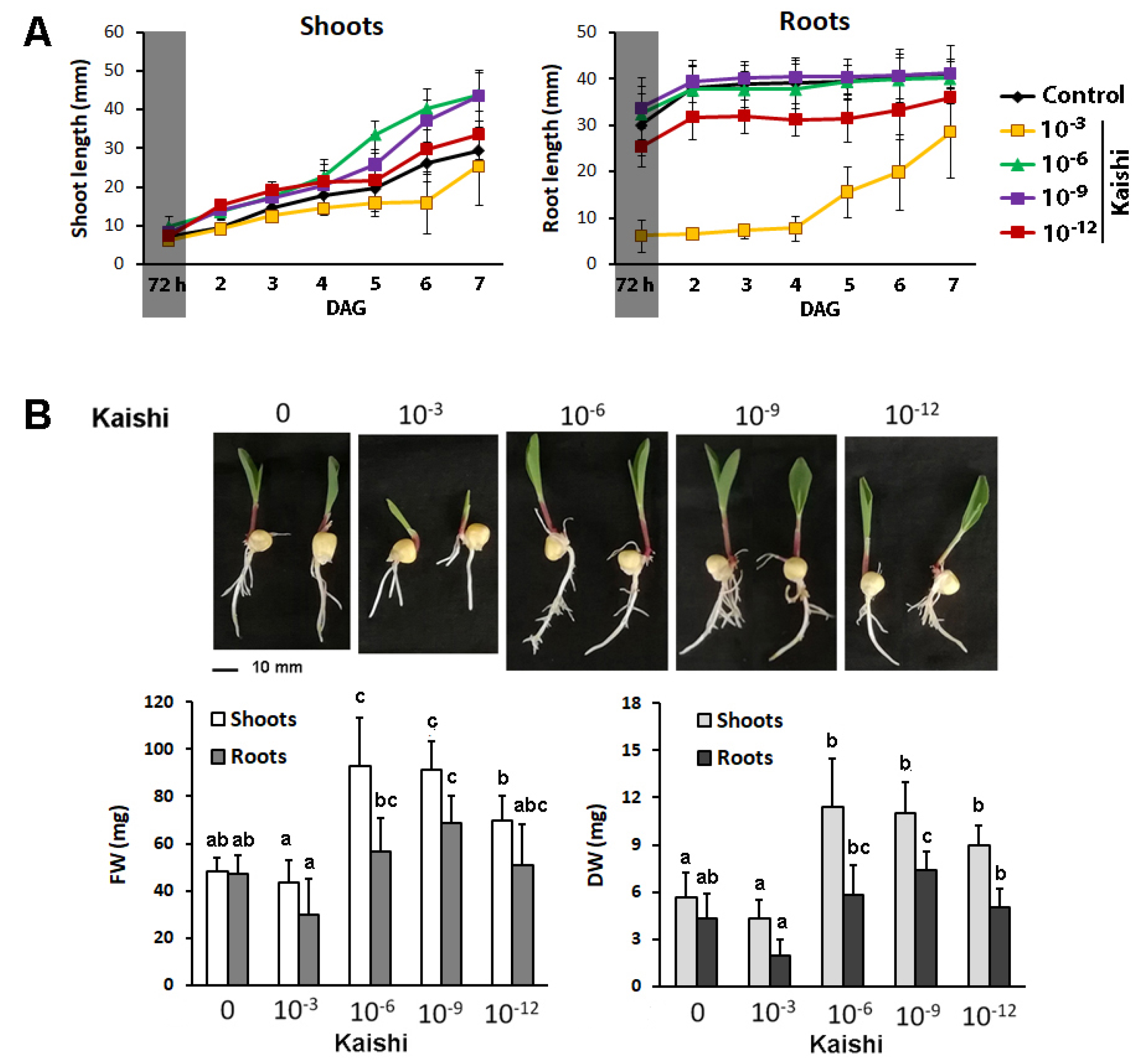
3.2. Growth-Stimulating Effects of the Biostimulant in Hydroponic Cultures
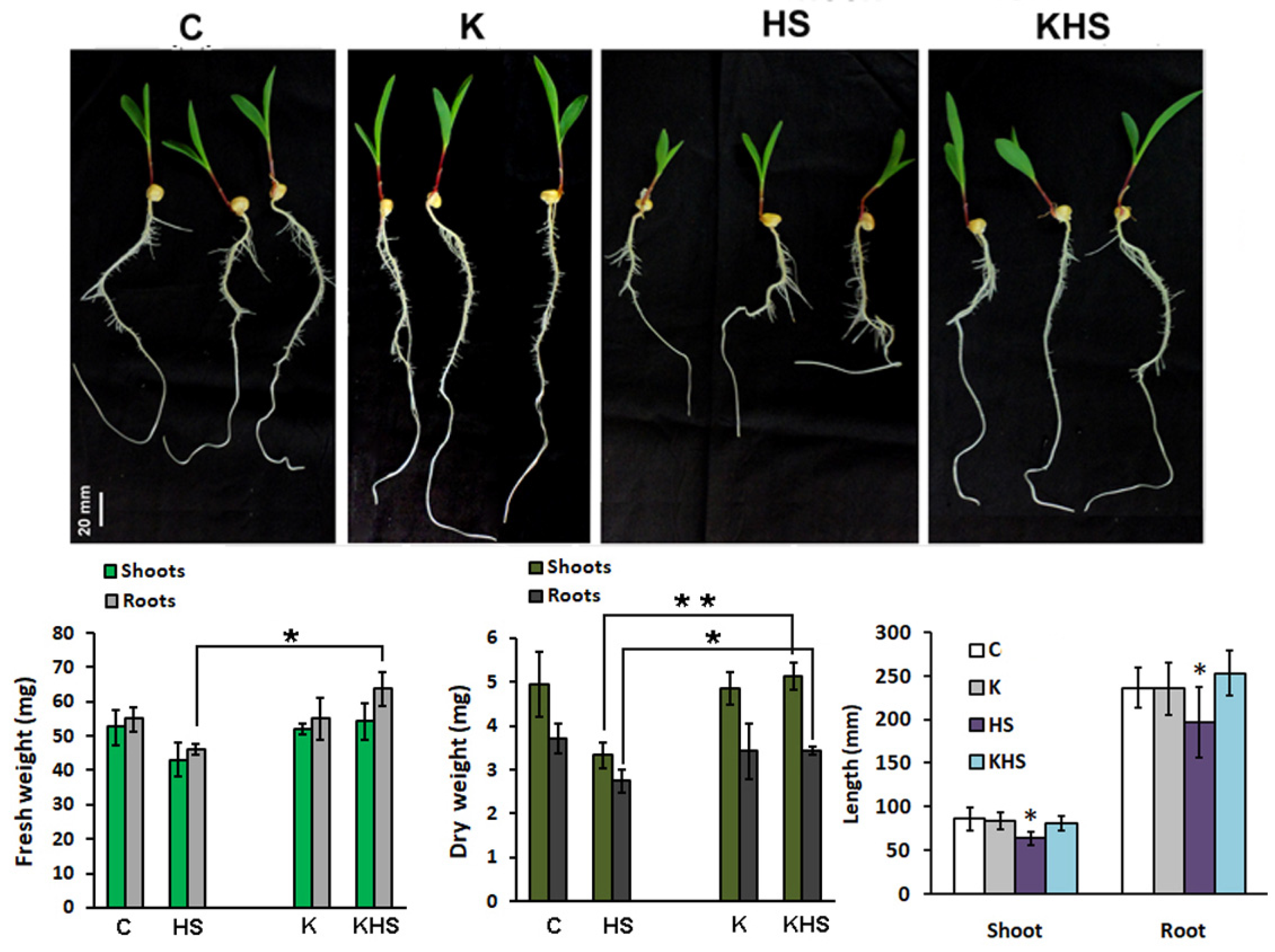
3.3. Pre-Treatment with Kaishi Mitigates the Negative Effect of Heat Shock in Hydroponically Grown Maize Plants
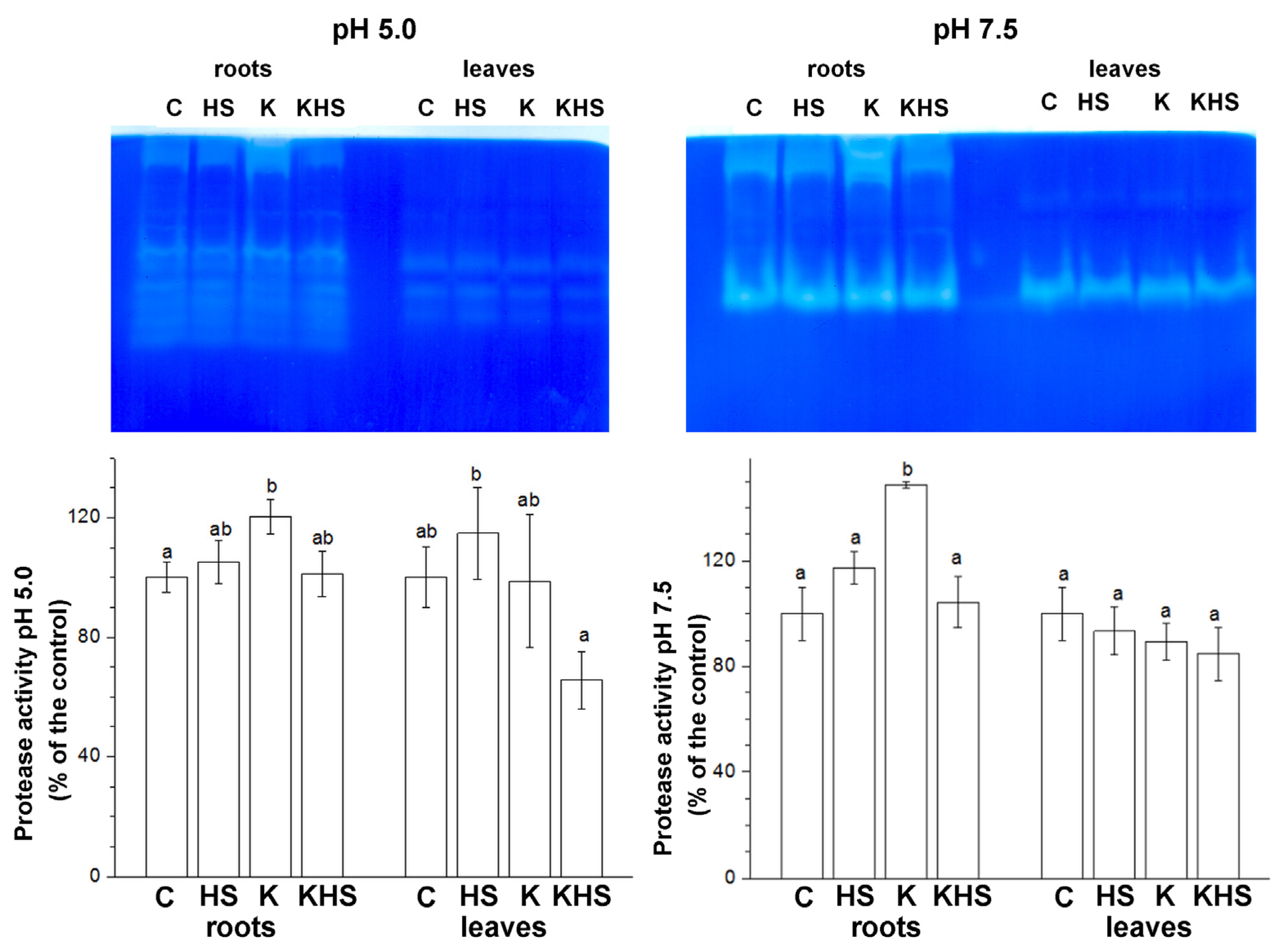
3.4. Protease Activity in Plants Treated with the Biostimulant and Subjected to Heat Shock
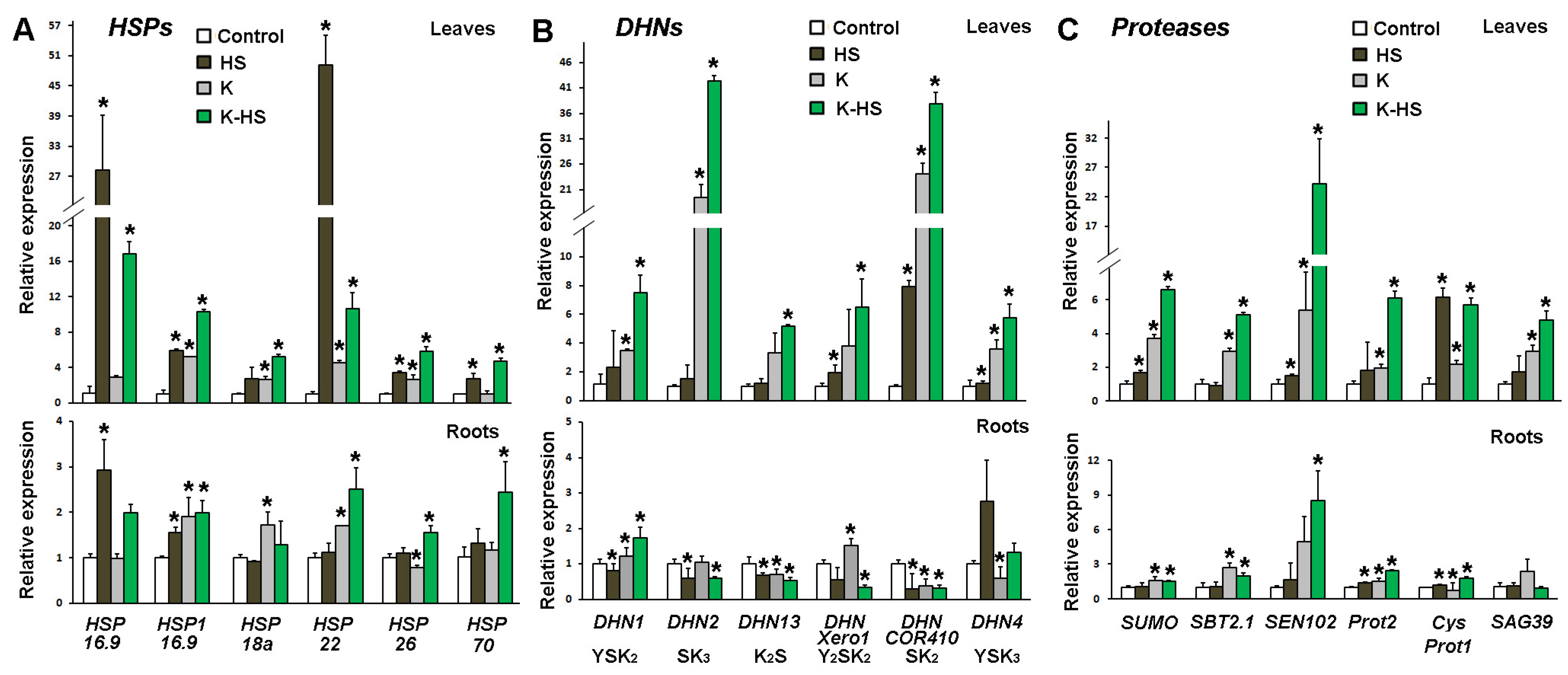
3.5. Transcript Profiling of HSP-, DHN-, and Protease-Coding Genes in Plants Treated with the Biostimulant and Subjected to Heat Shock
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Perkins-Kirkpatrick, S.E.; Gibson, P.B. Changes in regional heatwave characteristics as a function of increasing global temperature. Sci. Rep. 2017, 7, 12256. [Google Scholar] [CrossRef] [PubMed]
- Siebers, M.H.; Slattery, R.A.; Yendrek, C.R.; Locke, A.M.; Drag, D.; Ainsworth, E.A.; Bernacchi, C.J.; Ort, D.R. Simulated heat waves during maize reproductive stages alter reproductive growth but have no lasting effect when applied during vegetative stages. Agric. Ecosyst. Environ. 2017, 240, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Chukwudi, U.P.; Kutu, F.R.; Mavengahama, S. Influence of Heat Stress, Variations in Soil Type, and Soil Amendment on the Growth of Three Drought–Tolerant Maize Varieties. Agronomy 2021, 11, 1485. [Google Scholar] [CrossRef]
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G.E.S.J. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef]
- Lizaso, J.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.; Lorite, I.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crop. Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.A.; Wang, X.; Zafar, S.A.; Noor, M.A.; Hussain, H.A.; Azher Nawaz, M.; Farooq, M. Thermal Stresses in Maize: Effects and Management Strategies. Plants 2021, 10, 293. [Google Scholar] [CrossRef]
- Li, Z.; Howell, S.H. Heat Stress Responses and Thermotolerance in Maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef]
- Maiti, R.K.; Maiti, L.E.; Maiti, S.; Maiti, A.M.; Maiti, M.; Maiti, H. Genotypic variability in maize cultivars (Zea mays L.) for resistance to drought and salinity at the seedling stage. J. Plant Physiol. 1996, 148, 741–744. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfaalfa protein hydrolysate on the gene expression and activity of enzymes of TCA cycle and N metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- Ricci, M.; Tilbury, L.; Daridon, B.; Sukalac, K. General Principles to Justify Plant Biostimulant Claims. Front. Plant Sci. 2019, 10, 494. [Google Scholar] [CrossRef]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.B.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth and drought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- García-García, A.L.; García-Machado, F.J.; Borges, A.A.; Morales-Sierra, S.; Boto, A.; Jiménez-Arias, D. Pure Organic Active Compounds Against Abiotic Stress: A Biostimulant Overview. Front. Plant Sci. 2020, 11, 575829. [Google Scholar] [CrossRef] [PubMed]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Santi, C.; Zamboni, A.; Varanini, Z.; Pandolfini, T. Growth Stimulatory Effects and Genome-Wide Transcriptional Changes Produced by Protein Hydrolysates in Maize Seedlings. Front. Plant Sci. 2017, 8, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Wang, G.; Sui, J.; Liu, G.; Ma, F.; Bao, Z. Biostimulants alleviate temperature stress in tomato seedlings. Sci. Hortic. 2022, 293, 110712. [Google Scholar] [CrossRef]
- Botta, A. Enhancing plant tolerance to temperature stress with amino acids: An approach to their mode of action. Acta Hortic. 2013, 1009, 29–35. [Google Scholar] [CrossRef]
- Cholakova-Bimbalova, R.; Petrov, V.; Vassilev, A. Photosynthetic performance of young maize (Zea mays L.) plants exposed to chilling stress can be improved by the application of protein hydrolysates. Acta Agrobot. 2019, 72, 1769. [Google Scholar] [CrossRef]
- Cholakova-Bimbalova, R.; Koleva, L.; Vassilev, A. Effects of a biostimulant and a mineral fertilizer on the antioxidative defence system of chilling-exposed maize plants. Agric. Sci. 2018, 10, 33–40. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Grange, E.; Mannino, G.; van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance during Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11, 836. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Trevisan, S.; Manoli, A.; Quaggiotti, S. A novel biostimulant, belonging to protein hydrolysates, mitigates abiotic stress effects on maize seedlings grown in hydroponics. Agronomy 2019, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertani, A.; Nardi, S.; Francioso, O.; Sanchez-Cortes, S.; Foggia, M.D.; Schiavon, M. Effects of Two Protein Hydrolysates Obtained From Chickpea (Cicer arietinum L.) and Spirulina platensis on Zea mays (L.) Plants. Front. Plant Sci. 2019, 10, 954. [Google Scholar] [CrossRef]
- Vitale, E.; Velikova, V.; Tsonev, T.; Ferrandino, I.; Capriello, T.; Arena, C. The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill). Plants 2021, 10, 861. [Google Scholar] [CrossRef]
- Teixeira, W.F.; Fagan, E.B.; Soares, L.H.; Soares, J.N.; Reichardt, K.; Neto, D.D. Seed and Foliar Application of Amino Acids Improve Variables of Nitrogen Metabolism and Productivity in Soybean Crop. Front. Plant Sci. 2018, 9, 396. [Google Scholar] [CrossRef] [Green Version]
- Mola, I.D.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, L.; Rouphael, Y.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Gavelienė, V.; Pakalniškytė, L.; Novickienė, L.; Balčiauskas, L. Effect of biostimulants on cold resistance and productivity formation in winter rapeseed and winter wheat. Ir. J. Agric. Food Res. 2018, 57, 71–83. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Zhang, L. Structural and functional dynamics of dehydrins: A plant protector proteins under abiotic stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Ye, X.Y. Transcriptome response of maize (Zea mays L.) seedlings to heat stress. Protoplasma 2021, 259, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Xu, C. Identification and characterization of proteins associated with plant tolerance to heat stress. J. Integr. Plant Biol. 2008, 50, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; Koskull-Doring, P.; Vierling, E.; Scharf, K. Complexity of the heat stress response in plants. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Rashed, M.A.-S.; Abou-Deif, M.H.; Khalil, K.M.; Mahmoud, F.E.-S. Expression levels of heat shock proteins through western blot and real-time polymerase chain reaction in maize. Jordan J. Biol. Sci. 2021, 14, 671–676. [Google Scholar]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiol. Plant. 1997, 100, 291–296. [Google Scholar] [CrossRef]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P.; Chakrabartya, D. Dehydrin in the past four decades: From chaperones to transcription co-regulators in regulating abiotic stress response. Curr. Res. Biotechnol. 2021, 3, 249–259. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef]
- Vierstra, R.D. Proteolysis in plants: Mechanisms and functions. Plant Mol. Biol. 1996, 32, 275–302. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, L.; Wu, X.; Faleri, C.; Tai, F.; Zhang, M.; Liu, H.; Wang, W.; Cai, G. Genome-Wide Identification and Comparison of Cysteine Proteases in the Pollen Coat and Other Tissues in Maize. Front. Plant Sci. 2021, 12, 709534. [Google Scholar] [CrossRef]
- López-Hidalgo, C.; Meijón, M.; Lamelas, L.; Valledor, L. The rainbow protocol: A sequential method for quantifying pigments, sugars, free amino acids, phenolics, flavonoids and MDA from a small amount of sample. Plant Cell Environ. 2021, 44, 1977–1986. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gonzalez, A.; Steffen, K.L.; Lynch, J.P. Light and excess manganese. Implications for oxidative stress in common bean. Plant Physiol. 1998, 118, 493–504. [Google Scholar] [PubMed] [Green Version]
- Ornstein, L. Enzyme Bulletin; Canalco Industrial Corporation: Rockville, MD, USA, 1964; Volume 12. [Google Scholar]
- Woodbury, W.; Spencer, A.K.; Stahmann, M.A. An improved procedure using ferricyanide for detecting catalase isozymes. Anal. Biochem. 1971, 44, 301–305. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol. Biochem. 2010, 48, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Blaszczak, A.G.; Smith, R.; Gutierrez, A.; Galbraith, D.W.; Janda, J.; Vanier, C.; Wozniak, E.M. Molecular mechanism of action for the novel biostimulant CYT31 in plants exposed to drought stress. In II World Congress on the Use of Biostimulants in Agriculture; Brown, P., Muhammad, S., Eds.; ISHS: Leuven, Belgium, 2016. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, R.; Del Buono, D. Use of a Biostimulant to Mitigate Salt Stress in Maize Plants. Agronomy 2021, 11, 1755. [Google Scholar] [CrossRef]
- Staykov, N.S.; Angelov, M.; Petrov, V.; Minkov, P.; Kanojia, A.; Guinan, K.J.; Alseekh, S.; Fernie, A.R.; Sujeeth, N.; Gechev, T.S. An Ascophyllum nodosum-Derived Biostimulant Protects Model and Crop Plants from Oxidative Stress. Metabolites 2021, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Goher, M.; Iqbal, N. Heat stress-induced cell death, changes in antioxidants, lipid peroxidation, and protease activity in wheat leaves. J. Plant Growth Regul. 2012, 31, 283–291. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Carra, S.; Alberti, S.; Benesch, J.L.; Boelens, W.; Buchner, J.; Carver, J.A.; Cecconi, C.; Ecroyd, H.; Gusev, N.; Hightower, L.E.; et al. Small heat shock proteins: Multifaceted proteins with important implications for life. Cell Stress Chaperones 2019, 24, 295–308. [Google Scholar] [CrossRef]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Lund, A.A.; Blum, P.H.; Bhattramakki, D.; Elthon, T.E. Heat-stress response of maize mitochondria. Plant Physiol. 1998, 116, 1097–1110. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Yang, Y.; Gong, F.; Zhang, D.; Zhang, L.; Wu, L.; Li, C.; Wang, W. Protein sHSP26 improves chloroplast performance under heat stress by interacting with specific chloroplast proteins in maize (Zea mays). J. Prot. 2015, 115, 81–92. [Google Scholar] [CrossRef]
- Hara, M. The multifunctionality of dehydrins: An overview. Plant Signal. Behav. 2010, 5, 503–508. [Google Scholar] [CrossRef]
- Nagaraju, M.; Reddy, P.S.; Kumar, S.A.; Kumar, A.; Suravajhala, P.; Ali, A.; Srivastava, R.K.; Kavi Kishora, P.B.; Rao, D.M. Genome-wide in silico analysis of dehydrins in Sorghum bicolor, Setaria italica and Zea mays and quantitative analysis of dehydrin gene expressions under abiotic stresses in Sorghum bicolor. Plant Gene 2018, 13, 64–75. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Song, Q.; Zhang, T.; Li, D.; Yang, X. The maize late embryogenesis abundant protein ZmDHN13 positively regulates copper tolerance in transgenic yeast and tobacco. Crop J. 2019, 7, 403–410. [Google Scholar] [CrossRef]
- Schaffer, M.A.; Fischer, R.L. Transcriptional activation by heat and cold of a thiol protease gene in tomato. Plant Physiol. 1990, 93, 1486–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jespersen, D.; Belanger, F.C.; Huang, B. Candidate genes and molecular markers associated with heat tolerance in colonial Bentgrass. PLoS ONE 2017, 12, e0171183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustine, R.C.; York, S.L.; Rytz, T.C.; Vierstra, R.D. Defining the SUMO system in maize: SUMOylation is up-regulated during endosperm development and rapidly induced by stress. Plant Physiol. 2016, 171, 2191–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrell, R.; Sadanandom, A. Dealing with Stress: A Review of Plant SUMO Proteases. Front. Plant Sci. 2019, 10, 1122. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus | Forward PRIMER (5′–3′) | Reverse primer (5′–3′) |
|---|---|---|---|
| ZmDHN1 | LOC542373 | agggacagggacagtttcct | ccactcgcaagtgctgtacta |
| ZmDHN2 | LOC542251 | cgatcagaagcgttgcgttg | ggtctttaaagcacacgggc |
| ZmDHN13 | LOC100285266 | cacaaggaaggcatcgtgga | gctgcgacaccagatctcag |
| ZmDHNXero1 | LOC100279027 | cgtgcatattgctgtgctcc | agccagagccaaacctacac |
| ZmDHNCOR410 | LOC100281087 | gaaggtagctagcgttggca | accacgtcctacacaagcag |
| ZmDHN4 | LOC103635599 | cgccacaggcatatctggaa | tccttcaggcccttctttcg |
| ZmHSP1 | LOC100286044 | cggagaacaccaaggtggat | accttccacgtccaatcgtc |
| ZmHSP16.9 | LOC100280576 | cccaacccaatcccaatcca | cacggagaatgggtcgaaca |
| ZmHSP18a | LOC542293 | agttcatgcgcaagttcgtg | gacaacggtctcccctcag |
| ZmHSP22 | LOC100283239 | cgaagaagagtattggcggc | gatcgcacactttctctgcc |
| ZmHSP26 | LOC542576 | ccaagtagcgaaatggcagc | gtcgacactgttgtccctgt |
| ZmHSP70 | LOC103635762 | agccgatgatcgtggttagc | ttgaaataggcaggcacggt |
| ZmSUMO | LOC100280713 | tgcaggagaatggatgggac | gtcctcctgcggaagtagtg |
| ZmSBT2.1 | LOC100381627 | tctttgggtgttctcgcctc | cggcaaattaatggcgaggg |
| ZmSEN102 | LOC100280695 | gagaatggctacgtgcggat | acaccgtctcgttgagttgt |
| Zmprot2 | LOC100281516 | cgtcatgtccgatgtcaagc | gatacgggacgcctacagtg |
| ZmCysprot1 | LOC100283826 | gcatgaggacctcgatctgg | tacagcggattcatgggacg |
| ZmSAG39 | LOC103641507 | tagtggactgcgacgtgaac | tcctcgtagcccttgatgga |
| Zm ACT1 | LOC100282267 | cttcgaagaaaatgcggcgg | attctgctcgaagaggtggc |
| Zm TUB5 | LOC542248 | cctgcccaaggcaagagaaa | gaggaatcactgggcatggt |
| Zm αEF | LOC542581 | tgttctcactctcagacaccag | cccatggctgaaggaaaatgt |
| Priming | Treatment | Soluble Sugars (mg g−1 FW) | Starch (mg g−1 FW) | Total Phenolics (mg g−1 FW) | Flavonoids (mg g−1 FW) |
|---|---|---|---|---|---|
| K0 | C | 39.08 ± 2.32 e | 36.58 ± 3.44 f | 12.15 ± 0.26 ab | 4.47 ± 0.28 c |
| HS | 35.49 ± 2.19 cd | 10.68 ± 0.38 a | 12.9 ± 0.82 bc | 4.01 ± 0.12 a | |
| K10−3 | C | 31.46 ± 0.88 ab | 51.96 ± 4.59 h | 11.26 ± 0.44 a | 4.16 ± 0.15 ab |
| HS | 33.87 ± 1.76 bcd | 33.81 ± 1.88 def | 13.55 ± 0.21 c | 4.49 ± 0.29 c | |
| K10−6 | C | 35.48 ± 2.32 cd | 41.40 ± 2.45 g | 13.09 ± 0.15 bc | 4.29 ± 0.19 bc |
| HS | 35.87 ± 3.59 cde | 33.05 ± 1.69 cde | 13.55 ± 0.59 c | 4.14 ± 0.22 ab | |
| K10−9 | C | 32.97 ± 3.05 abc | 30.33 ± 1.57 bcd | 13.38 ± 0.40 c | 4.73 ± 0.18 d |
| HS | 33.92 ± 1.64 bcd | 34.23 ± 1.17 bc | 12.18 ± 0.22 a | 4.28 ± 0.24 bc | |
| K10−12 | C | 29.78 ± 2.39 a | 28.33 ± 1.18 ef | 13.08 ± 0.49 bc | 4.73 ± 0.11 d |
| HS | 37.15 ± 2.5 de | 30.87 ± 0.45 b | 14.83 ± 0.73 d | 4.10 ± 0.09 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaseva, I.I.; Simova-Stoilova, L.; Kostadinova, A.; Yuperlieva-Mateeva, B.; Karakicheva, T.; Vassileva, V. Heat-Stress-Mitigating Effects of a Protein-Hydrolysate-Based Biostimulant Are Linked to Changes in Protease, DHN, and HSP Gene Expression in Maize. Agronomy 2022, 12, 1127. https://doi.org/10.3390/agronomy12051127
Vaseva II, Simova-Stoilova L, Kostadinova A, Yuperlieva-Mateeva B, Karakicheva T, Vassileva V. Heat-Stress-Mitigating Effects of a Protein-Hydrolysate-Based Biostimulant Are Linked to Changes in Protease, DHN, and HSP Gene Expression in Maize. Agronomy. 2022; 12(5):1127. https://doi.org/10.3390/agronomy12051127
Chicago/Turabian StyleVaseva, Irina I., Lyudmila Simova-Stoilova, Anelia Kostadinova, Bistra Yuperlieva-Mateeva, Tania Karakicheva, and Valya Vassileva. 2022. "Heat-Stress-Mitigating Effects of a Protein-Hydrolysate-Based Biostimulant Are Linked to Changes in Protease, DHN, and HSP Gene Expression in Maize" Agronomy 12, no. 5: 1127. https://doi.org/10.3390/agronomy12051127
APA StyleVaseva, I. I., Simova-Stoilova, L., Kostadinova, A., Yuperlieva-Mateeva, B., Karakicheva, T., & Vassileva, V. (2022). Heat-Stress-Mitigating Effects of a Protein-Hydrolysate-Based Biostimulant Are Linked to Changes in Protease, DHN, and HSP Gene Expression in Maize. Agronomy, 12(5), 1127. https://doi.org/10.3390/agronomy12051127