
Glucose Increases the Abundance of Phosphate Solubilizing Bacterial Community for Better Apple Seedling Growth and Phosphate Uptake

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Sample Collection and Analysis
2.3. Soil DNA Isolation
2.4. PCR Amplifications and Illumina Library Generation
2.5. Statistical Analysis
3. Results
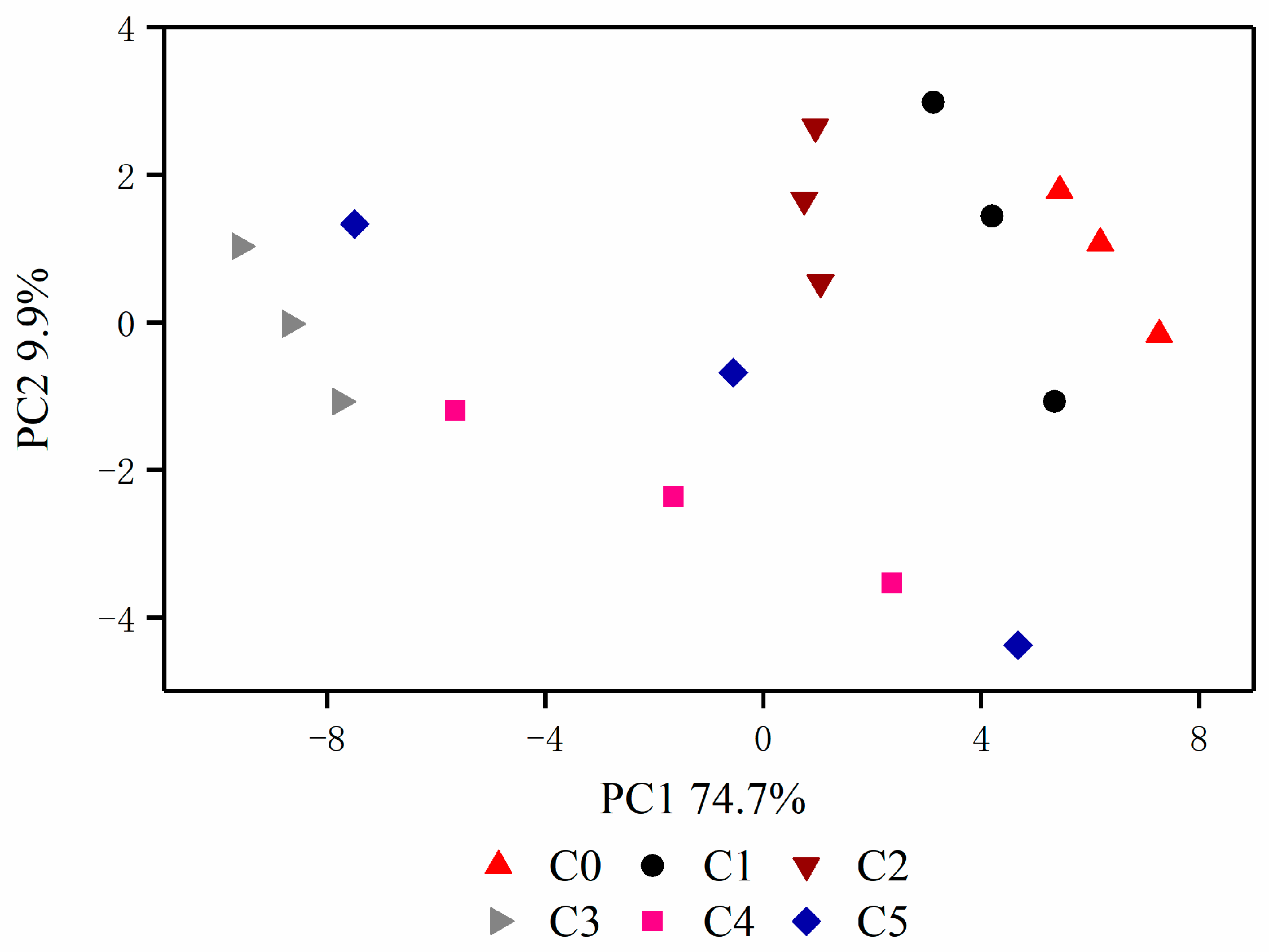
3.1. Classification and Sequencing of Bacterial Community
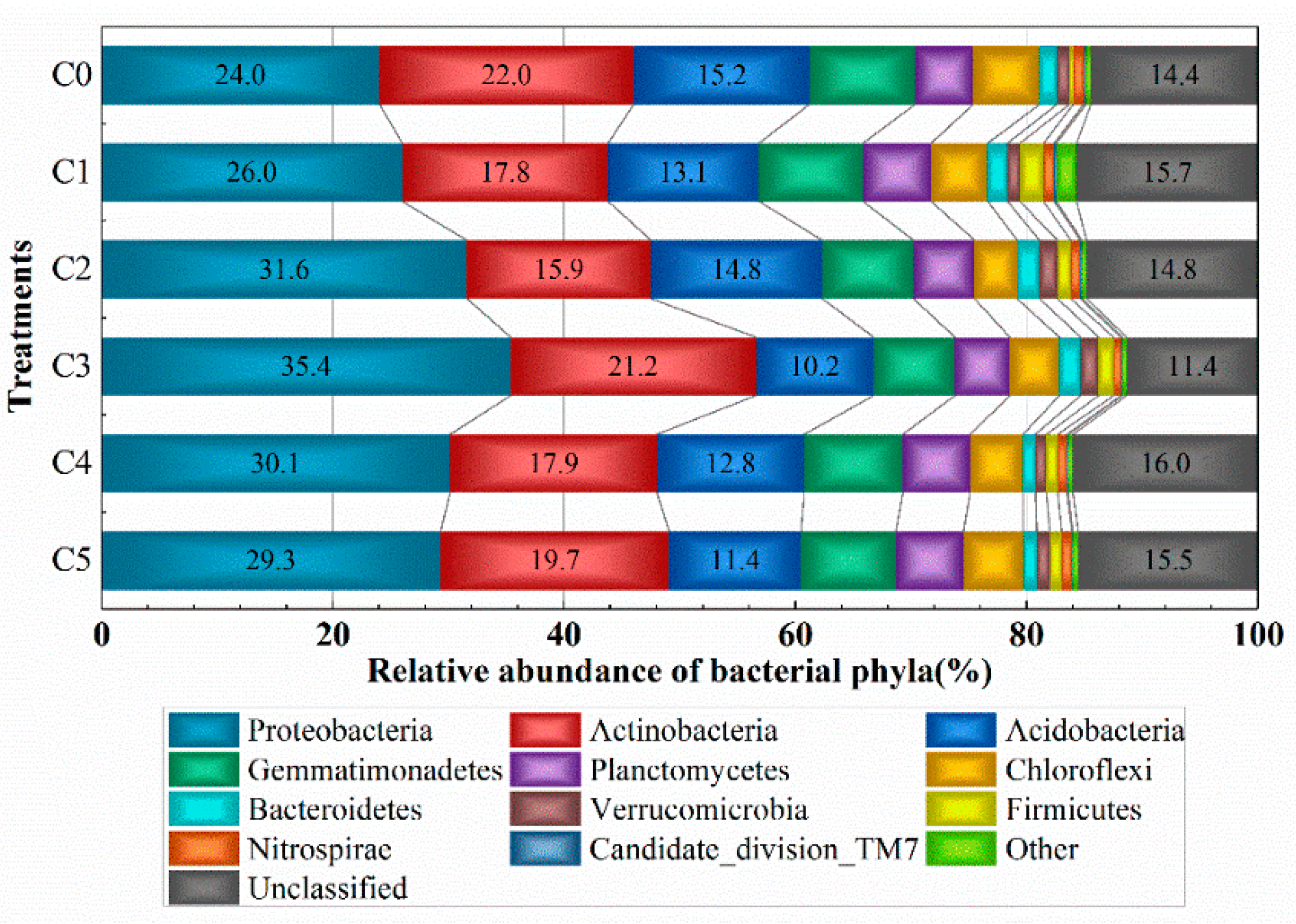
3.2. Classification and Relative Abundance of Different Taxa
3.3. Plant Growth and Phosphorus Uptake
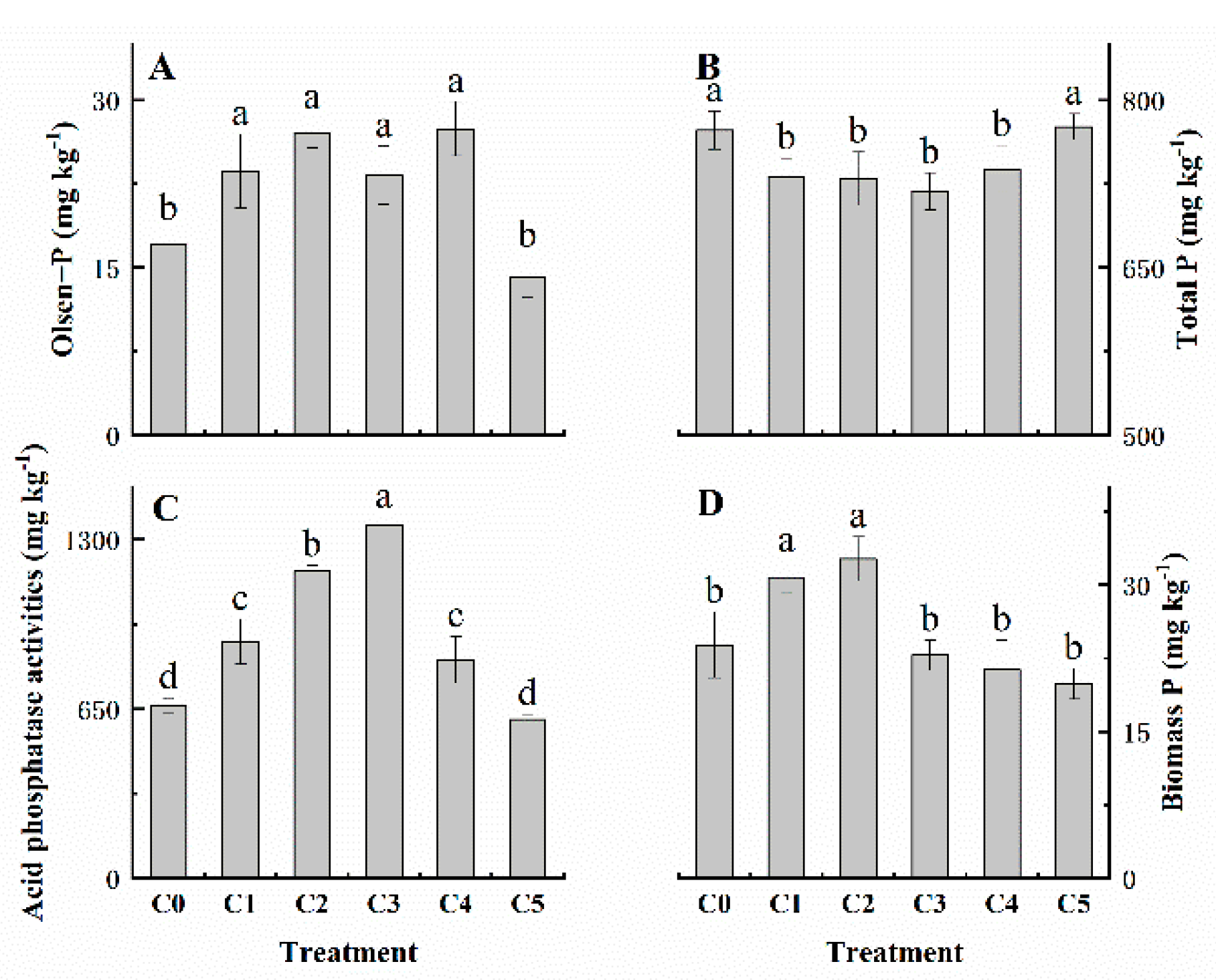
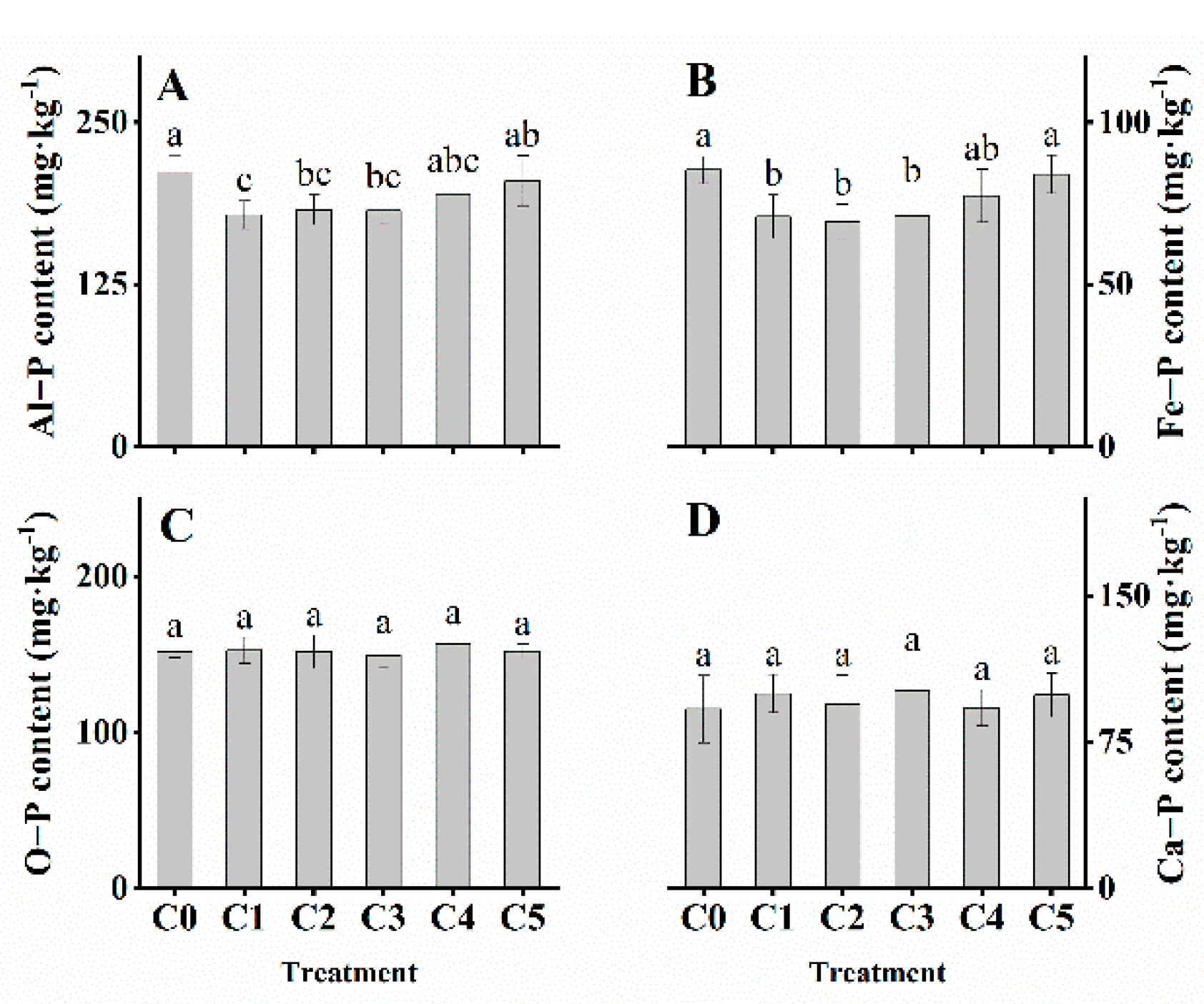
3.4. Soil Chemical Properties
3.5. Changes and Correlation Analysis of Phosphorus-Solubilizing Microorganisms
4. Discussion
4.1. Changes in Bacterial Communities
4.2. Changes in Soil Phosphorus Availability
4.3. Changes in Plant Phosphorous
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Balemi, T.; Negisho, K. Management of soil phosphorus and plant adaptation mechanisms to phosphorus stress for sustainable crop production: A review. J. Soil Sci. Plant Nutr. 2012, 12, 547–562. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Z.; Cheng, Y.; Shi, Y. Phosphorus availability in cropland soils of China and related affecting factors. J. Appl. Ecol. 2013, 24, 260–268. [Google Scholar]
- Zhu, Z.L.; Xia, Y.; Liu, J.J.; Ge, S.F.; Jiang, Y.M. Analysis of Soil Phosphorus Input and Phosphorus Environment Load Risk in Major Apple Production Regions of Shandong Province. Acta Hortic. Sin. 2017, 44, 97–105. [Google Scholar]
- Zhu, J.; Li, M.; Whelan, M. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of Phosphate Solubilization and Physiological Functions of Phosphate-Solubilizing Microorganisms; Springer International Publishing: Cham, Switzerland, 2014; pp. 31–62. [Google Scholar]
- Jenkinson, D.S. Cycles of Soils. Carbon, Nitrogen Phosphorus, Sulfur, Micronutrients; Wiley: New York, NY, USA, 1986. Available online: https://www.vgls.vic.gov.au/client/en_AU/VGLS-public/search/detailnonmodal/ent:$002f$002fSD_ILS$002f0$002fSD_ILS:28092/ada?qu=Soil+dynamics.&d=ent%3A%2F%2FSD_ILS%2F0%2FSD_ILS%3A28092%7EILS%7E44&ps=300&h=8 (accessed on 1 April 2022).
- Yadav, B.K.; Tarafdar, J.C.; Panwar, J. Microbial dynamics in rhizosphere of fruit plants during summer and monsoon in arid environment. Indian J. Hortic. 2011, 68, 161–169. [Google Scholar]
- Chen, C.R.; Condron, L.M.; Davis, M.R.; Sherlock, R.R. Seasonal changes in soil phosphorus and associated microbial properties under adjacent grassland and forest in New Zealand. For. Ecol. Manag. 2003, 177, 539–557. [Google Scholar] [CrossRef]
- Agbenin, J.O.; Adeniyi, T. The microbial biomass properties of a savanna soil under improved grass and legume pastures in northern Nigeria—ScienceDirect. Agric. Ecosyst. Environ. 2005, 109, 245–254. [Google Scholar] [CrossRef]
- Liu, S.Z.; Zhang, Q.; Zhao, C.J.; Bi, N.N.; Wang, X.W. Effect of Soil Organic Matter on Soil Characteristics and Fruit Quality in Orchard. J. Anhui Agric. Sci. 2010, 38, 21104–21106. [Google Scholar]
- Shen, F.; Wu, J.; Fan, H.; Liu, W.; Guo, X.; Duan, H.; Hu, L.; Lei, X.; Wei, X. Soil N/P and C/P ratio regulate the responses of soil microbial community composition and enzyme activities in a long-term nitrogen loaded Chinese fir forest. Plant Soil 2019, 436, 1–17. [Google Scholar] [CrossRef]
- Spohn, M.; Ermak, A.; Kuzyakov, Y. Microbial gross organic phosphorus mineralization can be stimulated by root exudates—A 33P isotopic dilution study. Soil Biol. Biochem. 2013, 65, 254–263. [Google Scholar] [CrossRef]
- Falchini, L.; Naumova, N.; Kuikman, P.J.; Bloem, J.; Nannipieri, P. CO2 evolution and denaturing gradient gel electrophoresis profiles of bacterial communities in soil following addition of low molecular weight substrates to simulate root exudation. Soil Biol. Biochem. 2003, 35, 775–782. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Wallace, H.M.; Xu, C.Y.; Van Zwieten, L.; Weng, Z.H.; Xu, Z.; Che, R.; Tahmasbian, I.; Hu, H.W.; Bai, S.H. The effects of short term, long term and reapplication of biochar on soil bacteria. Sci. Total Environ. 2018, 636, 142–151. [Google Scholar] [CrossRef]
- Yao, Q.; Liu, J.; Yu, Z.; Li, Y.; Jin, J.; Liu, X.; Wang, G. Changes of bacterial community compositions after three years of biochar application in a black soil of northeast China. Appl. Soil Ecol. 2017, 113, 11–21. [Google Scholar] [CrossRef]
- Cheng, C.G.; Zhao, D.Y.; Lv, D.G.; Jiang, M.; Du, G.D. Effects of plant-derived organic materials and humification driving forces on soil microbial community diversity in orchards. Plant Nutr. Fertil. Sci. 2014, 20, 913–922. [Google Scholar]
- Tian, W.; Wang, L.; Li, Y.; Zhuang, K.; Li, G.; Zhang, J.; Xiao, X.; Xi, Y. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agric. Ecosyst. Environ. 2015, 213, 219–227. [Google Scholar] [CrossRef]
- Jeffery, S.; Verheijen, F.G.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Abdallahi, M.M.; N’Dayegamiye, A. Effects of two incorporations of green manures on soil physical and biological properties and on wheat (Triticum aestivum L.) yields and N uptake. Can. J. Soil Sci. 2000, 80, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Bowden, C.L.; Evanylo, G.K.; Zhang, X.; Ervin, E.H.; Seiler, J.R. Soil Carbon and Physiological Responses of Corn and Soybean to Organic Amendments. Compost Sci. Util. 2010, 18, 162–173. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Liu, M.; Jiang, C.; Che, Y. Microbial community and functional diversity associated with different aggregate fractions of a paddy soil fertilized with organic manure and/or NPK fertilizer for 20 years. J. Soils Sediments 2015, 15, 292–301. [Google Scholar] [CrossRef]
- Young, I.; Crawford, J.; Nunan, N.; Otten, W.; Spiers, A. Microbial Distribution in Soils: Physics and Scaling. Adv. Agron. 2008, 100, 81–121. [Google Scholar]
- Zhang, L.; Ding, X.; Chen, S.; He, X.; Zhang, F.; Feng, G. Reducing carbon: Phosphorus ratio can enhance microbial phytin mineralization and lessen competition with maize for phosphorus. J. Plant Interact. 2014, 9, 850–856. [Google Scholar] [CrossRef]
- Ge, S.F.; Zhou, L.; Men, Y.G.; Li, H.N.; Wei, S.C.; Jiang, Y.M. Effect of Carbon Application on Nitrogen and Phosphorus Leaching in Apple Orchard Soil. J. Soil Water Conserv. 2013, 27, 31–35. [Google Scholar]
- Ding, X.D.; Zhang, S.R.; Lou, J.H.; Wang, Z.H.; Li, Y.; Zhang, L.; Wang, K.R. Effects of Combined Organic Manure and Phosphorus Fertilizer on the Phosphorus Leaching Risk in Coastal Saline Soil. Ecol. Environ. 2016, 25, 1169–1173. [Google Scholar]
- Song, Q.; Li, F.; Liu, H.; Wang, J.; Li, S. Effect of plastic film mulching on soil microbial biomass in spring wheat field in semi-arid loess area. J. Appl. Ecol. 2003, 9, 1512–1516. [Google Scholar]
- Fernando, P.S.; Jacobo, A.; Víctor, P. A novel conceptual approach to read-filtering in high-throughput amplicon sequencing studies. Nucleic Acids Res. 2015, 44, e40. [Google Scholar]
- Gu, Y.; Jiang, B. Determination of inorganic phosphorus classification in calcareous soil. Soils 1990, 101–102, 110. [Google Scholar]
- Anna, K.; Elmar, P.; Timmy, S.; Jrg, P.; Christian, Q.; Matthias, H.; Oliver, G.F. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar]
- Mago, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Reischke, S.; Rousk, J.; Bth, E. The effects of glucose loading rates on bacterial and fungal growth in soil. Soil Biol. Biochem. 2014, 70, 88–95. [Google Scholar] [CrossRef]
- Rinnan, R.; Baath, E. Differential Utilization of Carbon Substrates by Bacteria and Fungi in Tundra Soil. Appl. Environ. Microbiol. 2009, 75, 3611–3620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marianne, K.; Christina, K.; Lucia, F.; Barbara, K.; Angela, S.; Sophie, Z.B.; Andreas, R. Fungal and bacterial utilization of organic substrates depends on substrate complexity and N availability. FEMS Microbiol. Ecol. 2014, 87, 142–152. [Google Scholar]
- De Graaff, M.A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishore, N.; Pindi, P.K.; Reddy, S.R. Phosphate-Solubilizing Microorganisms: A Critical Review; Springer: New Delhi, India, 2015. [Google Scholar]
- Nobili, M.; Contin, M.; Mondini, C.; Brookes, P.C. Soil microbial biomass is triggered into activity by trace amounts of substrate. Soil Biol. Biochem. 2001, 33, 1163–1170. [Google Scholar] [CrossRef]
- el Zahar Haichar, F.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Nemergut, D.R.; Schmidt, S.K.; Townsend, A.R. Increases in soil respiration following labile carbon additions linked to rapid shifts in soil microbial community composition. Biogeochemistry 2007, 82, 229–240. [Google Scholar] [CrossRef]
- De Araújo, L.M.C.; Sampaio, E.V.D.S.B.; De Araújo, M.D.S.B. Phosphorus Desorption from Fe and Al Oxides Mediated by Soil Microorganisms. Commun. Soil Sci. Plant Anal. 2015, 46, 633–640. [Google Scholar] [CrossRef]
- Wu, J.; Min, H.; Xiao, H.A.; Su, Y.R.; Tong, C.L.; Huang, D.Y.; Syers, J.K. Dynamics in microbial immobilization and transformations of phosphorus in highly weathered subtropical soil following organic amendments. Plant Soil 2007, 290, 333–342. [Google Scholar] [CrossRef]
- Nahas, E. Phosphate Solubilizing Microorganisms: Effect of Carbon, Nitrogen, and Phosphorus Sources; Springer: Dordrecht, The Netherlands, 2007; pp. 111–115. [Google Scholar]
- Scervino, J.M.; Papinutti, V.L.; Godoy, M.S.; Rodriguez, M.A.; Monica, I.D.; Recchi, M.; Pettinari, M.J.; Godeas, A.M. Medium pH, carbon and nitrogen concentrations modulate the phosphate solubilization efficiency of Penicillium purpurogenum through organic acid production. J. Appl. Microbiol. 2011, 110, 1215–1223. [Google Scholar] [CrossRef]
- Sashidhar, B.; Podile, A.R. Mineral phosphate solubilization by rhizosphere bacteria and scope for manipulation of the direct oxidation pathway involving glucose dehydrogenase. J. Appl. Microbiol. 2010, 109, 1–12. [Google Scholar] [CrossRef]
- Spohn, M.; Kuzyakov, Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Biol. Biochem. 2013, 61, 69–75. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mccormack, M.; Li, L.; Hall, Q.; Xiang, C.; Sheen, J. Glucose-TOR signalling reprograms the transcriptome and activates meristems. Nature 2013, 496, 181. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Singh, M.; Laxmi, A. Interaction between Glucose and Brassinosteroid during the Regulation of Lateral Root Development in Arabidopsis. Plant Physiol. 2015, 168, 307–320. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| April | May | June | July | |
|---|---|---|---|---|
| Average rainfall (mm) | 27 | 43.6 | 90.6 | 209.9 |
| Average temperature (°C) | 13.7 | 19.1 | 24.4 | 26 |
| Treatment | Nseqsa | OTUs | Chao 1 | Ace | Shannon | Coverage |
|---|---|---|---|---|---|---|
| C0 | 22,305 ± 4862 ab | 2812 ± 189 ab | 3489 ± 188 a | 3533 ± 136 a | 6.986 ± 0.0111 a | 0.962 ± 0.0092 a |
| C1 | 22,993 ± 3231 a | 2881 ± 101 a | 3557 ± 221 a | 3610 ± 215 a | 6.985 ± 0.0068 a | 0.963 ± 0.0107 a |
| C2 | 17,870 ± 553 a | 2500 ± 220 cd | 3367 ± 131 ab | 3416 ± 141 ab | 6.827 ± 0.0513 b | 0.952 ± 0.0012 a |
| C3 | 20,793 ± 2805 a | 2608 ± 115 bc | 3372 ± 18 ab | 3451 ± 38 a | 6.490 ± 0.1890 d | 0.960 ± 0.0078 a |
| C4 | 22,099 ± 2325 a | 2675 ± 75 abc | 3494 ± 69 a | 3516 ± 52 ab | 6.608 ± 0.0596 c | 0.970 ± 0.0170 a |
| C5 | 17,883 ± 1193 a | 2362 ± 81 d | 3114 ± 337 b | 3148 ± 357 b | 6.56 ± 0.07630 cd | 0.956 ± 0.0114 a |
| Treatment | Total Biomass (g/Plant) | P Uptake (mg/Plant) | Total Root Length (cm) | Root Total Surface Area (cm2) | Number of Root Tips |
|---|---|---|---|---|---|
| C0 | 23.9 ± 3.6 c | 39.8 ± 5.8 b | 1818 ± 389 cd | 389 ± 26 c | 3507 ± 491 b |
| C1 | 28.8 ± 3.8 ab | 52.1 ± 3.3 a | 2451 ± 465 c | 608 ± 73 b | 5371 ± 249 a |
| C2 | 29.5 ± 2.6 ab | 52.5 ± 0.4 a | 4327 ± 575 a | 1020 ± 98 a | 4789 ± 602 a |
| C3 | 30.2 ± 2.4 a | 49.5 ± 2.7 a | 3487 ± 491 b | 1010 ± 128 a | 5126 ± 427 a |
| C4 | 25.3 ± 4.1 bc | 42.6 ± 2.2 b | 3630 ± 422 ab | 715 ± 67 b | 4880 ± 408 a |
| C5 | 20.9 ± 2.9 c | 30.1 ± 1.7 c | 1464 ± 312 d | 230 ± 32 d | 2046 ± 239 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Z.; Zhao, L.; Zhang, J.; Jiang, W.; Wei, M.; Xu, X.; Jiang, Y.; Ge, S. Glucose Increases the Abundance of Phosphate Solubilizing Bacterial Community for Better Apple Seedling Growth and Phosphate Uptake. Agronomy 2022, 12, 1181. https://doi.org/10.3390/agronomy12051181
Jia Z, Zhao L, Zhang J, Jiang W, Wei M, Xu X, Jiang Y, Ge S. Glucose Increases the Abundance of Phosphate Solubilizing Bacterial Community for Better Apple Seedling Growth and Phosphate Uptake. Agronomy. 2022; 12(5):1181. https://doi.org/10.3390/agronomy12051181
Chicago/Turabian StyleJia, Zhihang, Lin Zhao, Jia Zhang, Wei Jiang, Meng Wei, Xinxiang Xu, Yuanmao Jiang, and Shunfeng Ge. 2022. "Glucose Increases the Abundance of Phosphate Solubilizing Bacterial Community for Better Apple Seedling Growth and Phosphate Uptake" Agronomy 12, no. 5: 1181. https://doi.org/10.3390/agronomy12051181
APA StyleJia, Z., Zhao, L., Zhang, J., Jiang, W., Wei, M., Xu, X., Jiang, Y., & Ge, S. (2022). Glucose Increases the Abundance of Phosphate Solubilizing Bacterial Community for Better Apple Seedling Growth and Phosphate Uptake. Agronomy, 12(5), 1181. https://doi.org/10.3390/agronomy12051181