
Endophytic Bacteria Can Replace the Need for Synthetic Auxin during In Vitro Rooting of Pyrus communis

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and In Vitro Growth
2.2. Isolation and Identification of Endophytic Bacteria in Pyrus communis
2.3. Selection of IAA-Promoting Bacteria
2.4. 16S DNA Gene Sequencing of Isolated Microorganisms with Higher IAA Production
2.5. In Vitro Inoculation
2.6. Acclimatization
2.7. Ultrastructural Analysis by Scanning Electron Microscopy
2.8. Experimental Design and Statistical Analyses
2.9. Experiment Workflow
3. Results
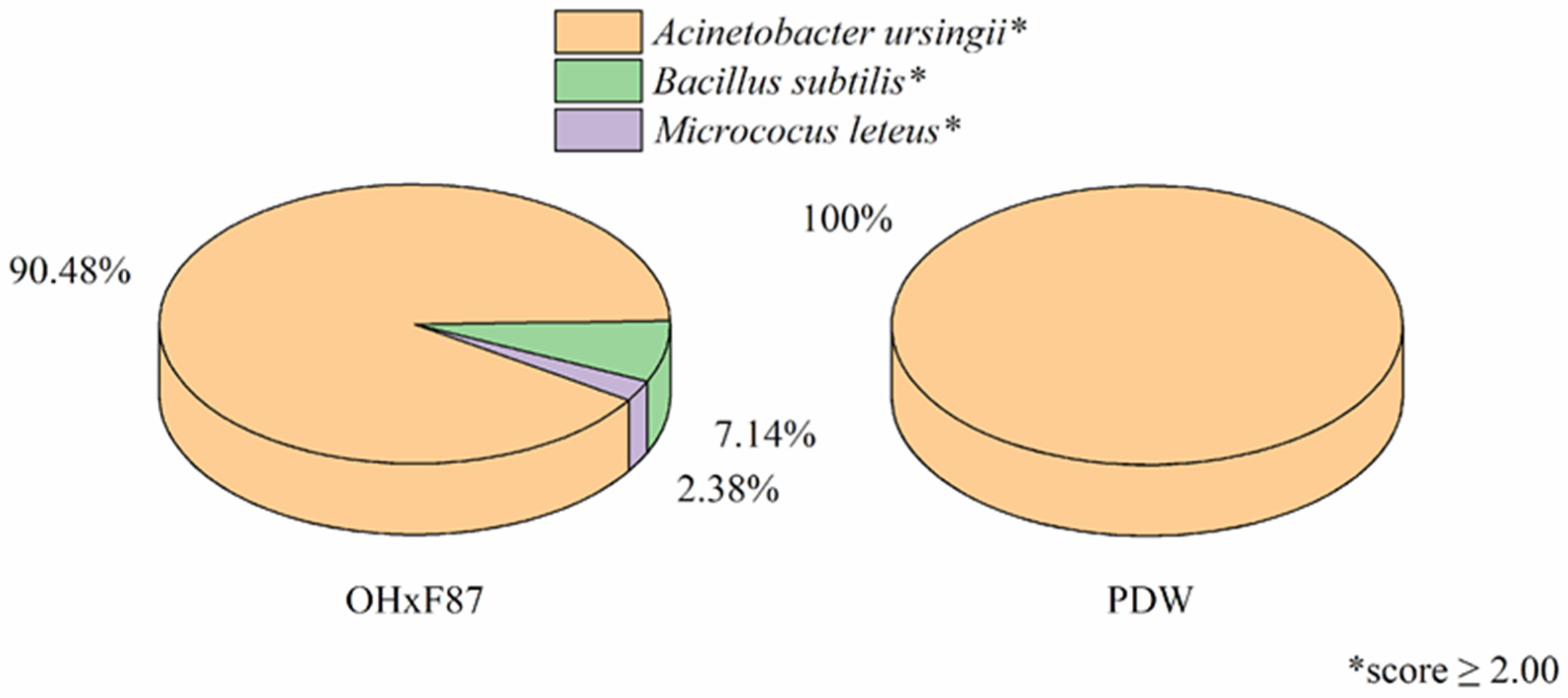
3.1. Isolation and Identification Using MALDI-TOF MS of Endophytic Bacteria in Pyrus communis Rootstocks
3.2. Selection of IAA-Promoting Bacteria
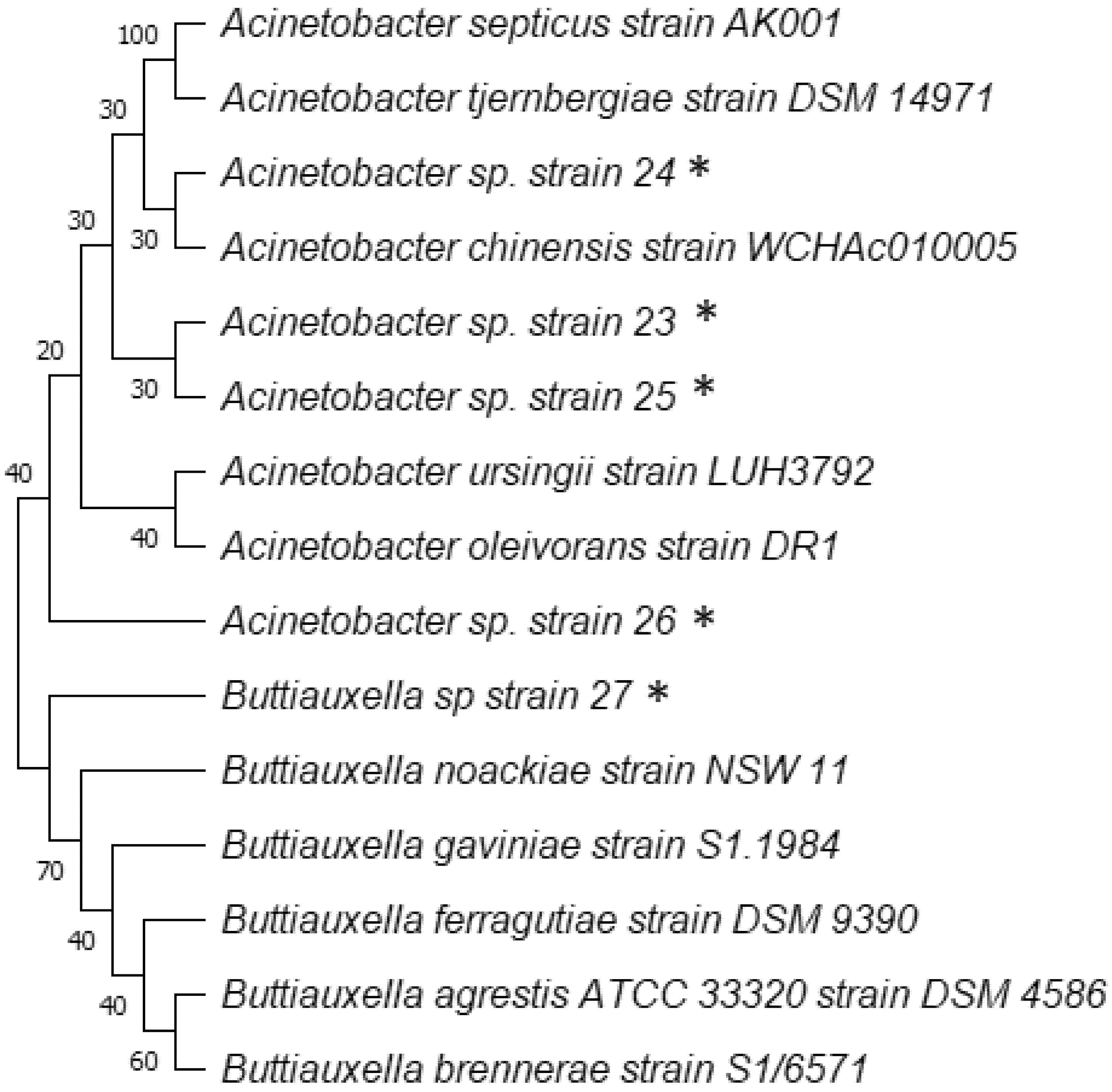
3.3. 16S DNA Gene Sequencing of Isolated Microorganisms with Higher AIA Production
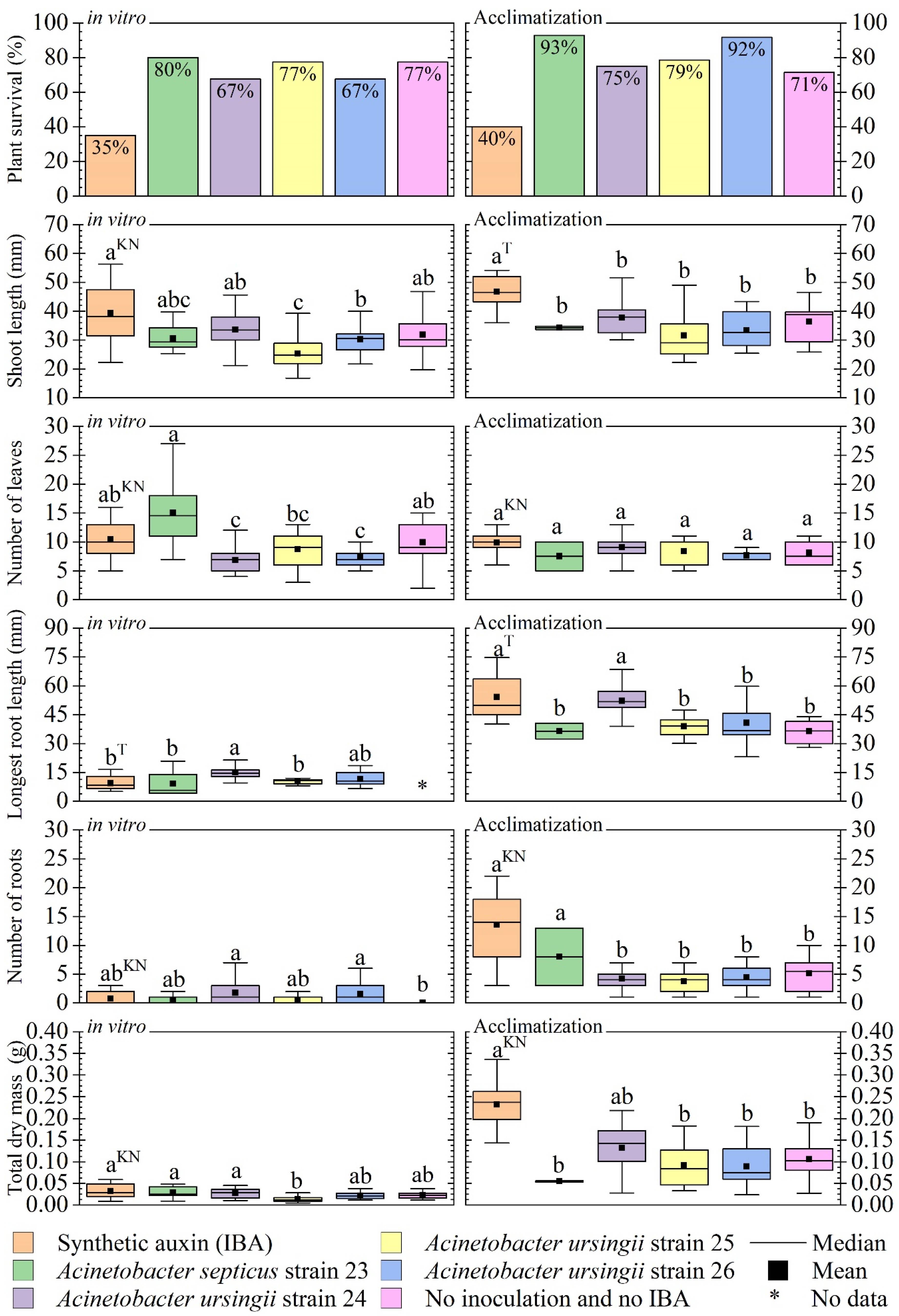
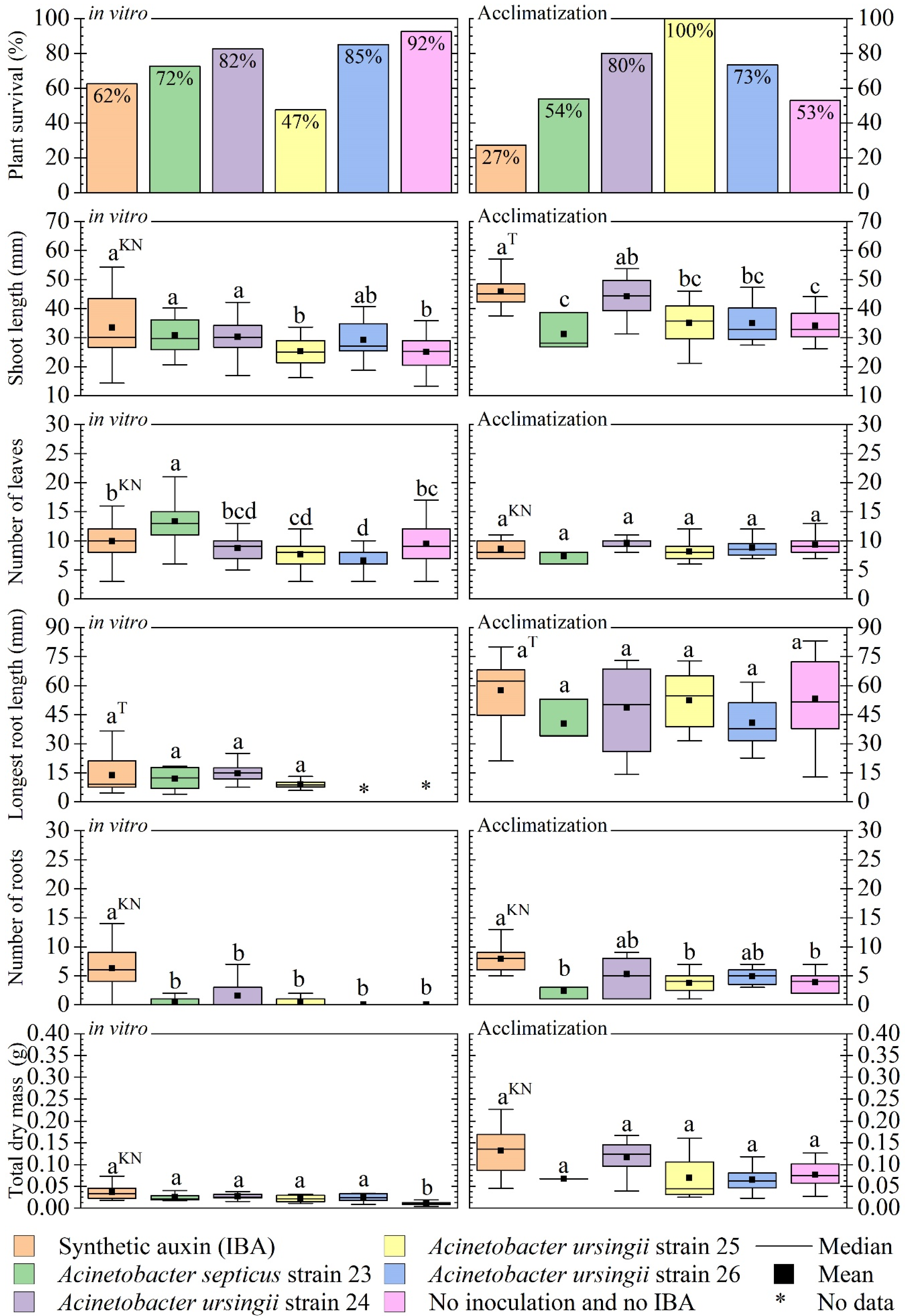
3.4. Evaluation of In Vitro Inoculation: In Vitro Development and Acclimatization
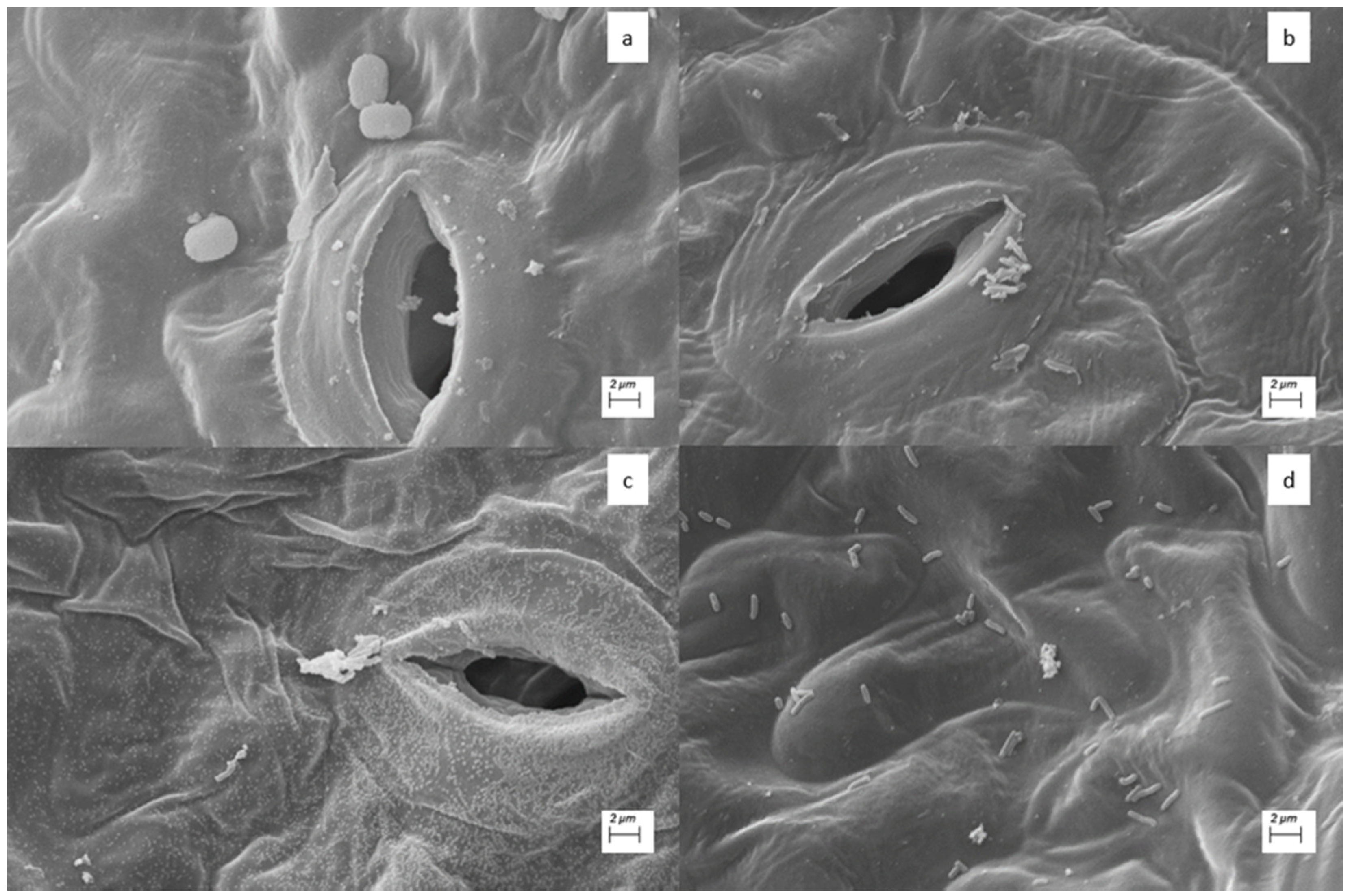
3.5. Ultrastructural Analysis by Scanning Electron Microscopy
4. Discussion
4.1. Isolation and Identification of Endophytic Bacteria in Pyrus communis Rootstocks
4.2. Selection of IAA-Promoting Bacteria
4.3. Evaluation of In Vitro Inoculation: In Vitro Development and Acclimatization
4.4. Ultrastructural Analysis by Scanning Electron Microscopy
4.5. General Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rufato, L.; Marcon Filho, J.L.; Marodin, G.A.B.; Kretzschmar, A.A.; Miqueluti, D.J. Intensidade e Épocas de Poda Verde Em Pereira “Abate Fetel” Sobre Dois Porta-Enxertos. Rev. Bras. Frutic. 2012, 34, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Hawerroth, F.J.; Herter, F.G.; Prezotto, M.E.; Haas, L.B.; Pretto, A. Aumento Da Produção de Pereira Asiática Pelo Uso de Fitorreguladores. Ciência Rural. St. Maria 2011, 41, 1750–1754. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies Junior, F.T.; Geneve, R.L. Plant Propagation: Principles and Practices, 7th ed.; Prentice Hall: Hoboken, NJ, USA, 2002. [Google Scholar]
- Park, S. Plant Tissue Culture; Elsevier: Amsterdam, The Netherlands, 2021; ISBN 978-0-12-821120-5. [Google Scholar]
- Ercisli, S.; Esitken, A.; Orhan, E.; Ozdemir, O. Rootstocks Used for Temperate Fruit Trees in Turkey: An Overview. Sodinink. Daržinink. 2006, 25, 27–33. [Google Scholar]
- Dimitrova, N.; Nacheva, L. An Optimized Micropropagation Protocol by Ex Vitro Rooting of Pear Rootstock OHF 333 (Pyrus communis L.). Acta Agrobot. 2021, 74, 744. [Google Scholar] [CrossRef]
- Apple & Pear Australia LTD (APAL) Rootstocks. Available online: https://apal.org.au/industry-info/intensive-pear-production/rootstocks/#pr (accessed on 15 July 2019).
- Washington State University (WSU) Rootstocks for Pear. Available online: http://treefruit.wsu.edu/web-article/pear-rootstocks/ (accessed on 15 July 2019).
- Grimaldi, F.; Grohskopf, M.A.; Muniz, A.W.; Frederico, A. Enraizamento in Vitro de Frutíferas Da Família Rosaceae. Rev. Ciências Agroveterinárias 2008, 7, 160–168. [Google Scholar]
- Aygun, A.; Dumanoglu, H. In Vitro Shoot Proliferation and In Vitro and Ex Vitro Root Formation of Pyrus Elaeagrifolia Pallas. Front. Plant Sci. 2015, 6, 225. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, D.; Wang, C.; Wang, X.; Li, J.; Wang, R. Construction of High Efficiency Regeneration and Transformation Systems of Pyrus Ussuriensis Maxim. Plant Cell Tissue Organ Cult. 2017, 131, 139–150. [Google Scholar] [CrossRef]
- Norton, M.E. The Role of Auxin and Cytokinin in the Control of Morphogenesis In Vitro in the Plant Families “Rosaceae” and “Ericaceae”; University of St. Andrews: St. Andrews, UK, 1984. [Google Scholar]
- Orlikowska, T.; Nowak, K.; Reed, B. Bacteria in the Plant Tissue Culture Environment. Plant Cell Tissue Organ Cult. 2017, 128, 487–508. [Google Scholar] [CrossRef]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.-S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The Use of Microbial Inoculants for Biological Control, Plant Growth Promotion, and Sustainable Agriculture: A Review. Eur. J. Plant Pathol. 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Souza, J.A.; Bettoni, J.C.; Costa, M.D.; Baldissera, T.C.; dos Passos, J.F.M.; Primieri, S. In Vitro Rooting and Acclimatization of ‘Marubakaido’ Apple Rootstock Using Indole-3-Acetic Acid from Rhizobacteria. Commun. Plant Sci. 2022, 12, 16–23. [Google Scholar] [CrossRef]
- Erturk, Y.; Ercisli, S.; Haznedar, A.; Cakmakci, R. Effects of Plant Growth Promoting Rhizobacteria (PGPR) on Rooting and Root Growth of Kiwifruit (Actinidia deliciosa) Stem Cuttings. Biol. Res. 2010, 43, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Montero-Calasanz, M.C.; Santamaría, C.; Albareda, M.; Daza, A.; Duan, J.; Glick, B.R.; Camacho, M. Alternative Rooting Induction of Semi-Hardwood Olive Cuttings by Several Auxin-Producing Bacteria for Organic Agriculture Systems. Span. J. Agric. Res. 2013, 11, 146–154. [Google Scholar] [CrossRef]
- Pretty, J. Agricultural Sustainability: Concepts, Principles and Evidence. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 447–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Keswani, C.; Singh, S.P.; Cueto, L.; García-Estrada, C.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Singh, S.P.; Blázquez, M.A.; Sansinenea, E. Auxins of Microbial Origin and Their Use in Agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 8549–8565. [Google Scholar] [CrossRef]
- Ali, M.; Boonerjee, S.; Islam, M.N.; Saha, M.L.; Hoque, M.I.; Sarker, R.H. Endogenous Bacterial Contamination of Plant Tissue Culture Materials: Identification and Control Strategy. Plant Tissue Cult. Biotechnol. 2018, 28, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Örgeç, M.Ö.; Pehlivan Karakaş, F.; Şahin, G.; Ağıl, F.; Zencirci, N. Einkorn (Triticum monococcum ssp. monococcum) in vitro propagation sterilization protocol. Int. J. Second. Metab. 2018, 5, 67–74. [Google Scholar] [CrossRef]
- Quambusch, M.; Winkelmann, T. Bacterial Endophytes in Plant Tissue Culture: Mode of Action, Detection, and Control. In Plant Cell Culture Protocols, Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1815, pp. 69–88. ISBN 9781493985944. [Google Scholar]
- Quambusch, M.; Brümmer, J.; Haller, K.; Winkelmann, T.; Bartsch, M. Dynamics of endophytic bacteria in plant in vitro culture: Quantification of three bacterial strains in Prunus avium in different plant organs and in vitro culture phases. Plant Cell Tissue Organ Cult. 2016, 126, 305–317. [Google Scholar] [CrossRef]
- Quambusch, M.; Pirttila, A.M.; Tejesvi, M.V.; Winkelmann, T.; Bartsch, M. Endophytic Bacteria in Plant Tissue Culture: Differences between Easy- and Difficult-to-Propagate Prunus Avium Genotypes. Tree Physiol. 2014, 34, 524–533. [Google Scholar] [CrossRef]
- Pirttilä, A.M.; Podolich, O.; Koskimäki, J.J.; Hohtola, E.; Hohtola, A. Role of Origin and Endophyte Infection in Browning of Bud-Derived Tissue Cultures of Scots Pine (Pinus sylvestris L.). Plant Cell Tissue Organ Cult. 2008, 95, 47–55. [Google Scholar] [CrossRef]
- Pohjanen, J.; Koskimaki, J.J.; Sutela, S.; Ardanov, P.; Suorsa, M.; Niemi, K.; Sarjala, T.; Haggman, H.; Pirttila, A.M. Interaction with ectomycorrhizal fungi and endophytic Methylobacterium affects nutrient uptake and growth of pine seedlings in vitro. Tree Physiol. 2014, 34, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Microbial Producers of Plant Growth Stimulators and Their Practical Use: A Review. Appl. Biochem. Microbiol. 2006, 42, 117–126. [Google Scholar] [CrossRef]
- Naik, K.; Mishra, S.; Srichandan, H.; Singh, P.K.; Sarangi, P.K. Plant Growth Promoting Microbes: Potential Link to Sustainable Agriculture and Environment. Biocatal. Agric. Biotechnol. 2019, 21, 101326. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of Action of Plant Growth Promoting Bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic Microbes: Biodiversity, Plant Growth-Promoting Mechanisms and Potential Applications for Agricultural Sustainability. Antonie Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and Plant-Microbe Interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [Green Version]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-Acetic Acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, M.; Srivastava, S. Stress-Responsive Indole-3-Acetic Acid Biosynthesis by Azospirillum brasilense SM and Its Ability to Modulate Plant Growth. Eur. J. Soil Biol. 2009, 45, 73–80. [Google Scholar] [CrossRef]
- Ahmed, A.; Hasnain, S. Auxin-producing Bacillus sp.: Auxin quantification and effect on the growth of Solanum tuberosum. Pure Appl. Chem. 2010, 82, 313–319. [Google Scholar] [CrossRef]
- Hossain, M.T.; Khan, A.; Harun-Or-Rashid, M.; Chung, Y.R. A volatile producing endophytic Bacillus siamensis YC7012 promotes root development independent on auxin or ethylene/jasmonic acid pathway. Plant Soil 2019, 439, 309–324. [Google Scholar] [CrossRef]
- López-Bucio, J.; Campos-Cuevas, J.C.; Hernández-Calderón, E.; Velásquez-Becerra, C.; Farías-Rodríguez, R.; Macías-Rodríguez, L.I.; Valencia-Cantero, E. Bacillus Megaterium Rhizobacteria Promote Growth and Alter Root-System Architecture through an Auxin- and Ethylene-Independent Signaling Mechanism in Arabidopsis Thaliana. Mol. Plant-Microbe Interact. 2007, 20, 207–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, D.; Mora, V.; Lopez, G.; Rosas, S.; Spaepen, S.; Vanderleyden, J.; Cassan, F. New insights into indole-3-acetic acid metabolism in Azospirillum brasilense. J. Appl. Microbiol. 2018, 125, 1774–1785. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Castro, R.; Campos-García, J.; López-Bucio, J. Pseudomonas putida and Pseudomonas fluorescens Influence Arabidopsis Root System Architecture through an Auxin Response Mediated by Bioactive Cyclodipeptides. J. Plant Growth Regul. 2019, 39, 254–265. [Google Scholar] [CrossRef]
- Karadeniz, A.; Topcuoğlu, Ş.F.; Inan, S. Auxin, Gibberellin, Cytokinin and Abscisic Acid Production in Some Bacteria. World J. Microbiol. Biotechnol. 2006, 22, 1061–1064. [Google Scholar] [CrossRef]
- Lin, H.-R.; Shu, H.-Y.; Lin, G.-H. Biological roles of indole-3-acetic acid in Acinetobacter baumannii. Microbiol. Res. 2018, 216, 30–39. [Google Scholar] [CrossRef]
- Foo, E.; Plett, J.M.; Lopez-Raez, J.A.; Reid, D. Editorial: The Role of Plant Hormones in Plant-Microbe Symbioses. Front. Plant Sci. 2019, 10, 1391. [Google Scholar] [CrossRef]
- Larraburu, E.E.; Carletti, S.M.; Cáceres, E.A.R.; Llorente, B.E. Micropropagation of photinia employing rhizobacteria to promote root development. Plant Cell Rep. 2007, 26, 711–717. [Google Scholar] [CrossRef]
- Peñafiel-Jaramillo, M.; Torres-Navarrete, E.D.; Barrera-Álvarez, A.E.; Prieto-Encalada, H.; Morante Carriel, J.; Canchignia Martínez, H.F. Producción de Ácido Indol-3-Acético Por Pseudomonas Veronii R4 y Formación de Raíces En Hojas de Vid “Thompson Seedless” in vitro. Cienc. Tecnol. 2016, 9, 31. [Google Scholar] [CrossRef]
- Russo, A.; Vettori, L.; Felici, C.; Fiaschi, G.; Morini, S.; Toffanin, A. Enhanced micropropagation response and biocontrol effect of Azospirillum brasilense Sp245 on Prunus cerasifera L. clone Mr.S 2/5 plants. J. Biotechnol. 2008, 134, 312–319. [Google Scholar] [CrossRef]
- Vettori, L.; Russo, A.; Felici, C.; Fiaschi, G.; Morini, S.; Toffanin, A. Improving micropropagation: Effect of Azospirillum brasilense Sp245 on acclimatization of rootstocks of fruit tree. J. Plant Interact. 2010, 5, 249–259. [Google Scholar] [CrossRef]
- Leblay, C.; Chevreau, E.; Raboin, L.M. Adventitious shoot regeneration from in vitro leaves of several pear cultivars (Pyrus communis L.). Plant Cell Tissue Organ Cult. 1991, 25, 99–105. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Carvalho, B.; Ávila, C.; Bernardes, T.; Pereira, M.N.; Santos, C.; Schwan, R. Fermentation profile and identification of lactic acid bacteria and yeasts of rehydrated corn kernel silage. J. Appl. Microbiol. 2017, 122, 589–600. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S RRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackbrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. ISBN 0-471-92906-9. [Google Scholar]
- Bozzola, J.J.; Russel, L.D. Electron Microscopy: Principles and Techniques for Biologists; Jones & Bartlett Learning: Burlington, MA, USA, 1999; ISBN 0763701920. [Google Scholar]
- Alves, E. Curso Introdutório à Microscopia Eletrônica de Varredura; Editora Universitária-Uniersidade Federal de Lavras: Lavras, Brazil, 2005. [Google Scholar]
- Nemec, A.; Radolfová-Křížová, L.; Maixnerová, M.; Shestivska, V.; Španělová, P.; Higgins, P.G. Acinetobacter silvestris sp. nov. discovered in forest ecosystems in Czechia. Int. J. Syst. Evol. Microbiol. 2022, 72, 005383. [Google Scholar] [CrossRef]
- Mujumdar, S.; Joshi, P.; Karve, N. Production, characterization, and applications of bioemulsifiers (BE) and biosurfactants (BS) produced by Acinetobacter spp.: A review. J. Basic Microbiol. 2019, 59, 277–287. [Google Scholar] [CrossRef]
- Guardabassi, L.; Dalsgaard, A.; Olsen, J.E. Phenotypic characterization and antibiotic resistance of Acinetobacter spp. isolated from aquatic sources. J. Appl. Microbiol. 1999, 87, 659–667. [Google Scholar] [CrossRef]
- Pérez-Flores, P.; Valencia-Cantero, E.; Altamirano-Hernández, J.; Pelagio-Flores, R.; López-Bucio, J.; García-Juárez, P.; Macías-Rodríguez, L. Bacillus methylotrophicus M4-96 isolated from maize (Zea mays) rhizoplane increases growth and auxin content in Arabidopsis thaliana via emission of volatiles. Protoplasma 2017, 254, 2201–2213. [Google Scholar] [CrossRef]
- Shi, Y.; Lou, K.; Li, C. Growth promotion effects of the endophyte Acinetobacter johnsonii strain 3-1 on sugar beet. Symbiosis 2011, 54, 159–166. [Google Scholar] [CrossRef]
- Sachdev, D.; Nema, P.; Dhakephalkar, P.; Zinjarde, S.; Chopade, B. Assessment of 16S rRNA gene-based phylogenetic diversity and promising plant growth-promoting traits of Acinetobacter community from the rhizosphere of wheat. Microbiol. Res. 2010, 165, 627–638. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, F.M.; Pereira, T.D.A.; Souza, T.P.; Guimarães, P.H.S.; Martins, A.D.; Schwan, R.; Pasqual, M.; Dória, J. Beneficial effects of inoculation of growth-promoting bacteria in strawberry. Microbiol. Res. 2019, 223–225, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Martínez-Viveros, O.; Jorquera, M.; Crowley, D.; Gajardo, G.; Mora, M. Mechanisms and Practical Considerations Involved in Plant Growth Promotion by Rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Singh, I. Plant Growth Promoting Rhizobacteria (PGPR) and Their Various Mechanisms for Plant Growth Enhancement in Stressful Conditions: A Review. Eur. J. Biol. Res. 2018, 8, 191–213. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of Plant Growth Promoting Rhizobacteria (PGPRs) with Multiple Plant Growth Promoting Traits in Stress Agriculture: Action Mechanisms and Future Prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Lizárraga, A.; Fraga, M.; Ascasíbar, J.; González, M.L. In Vitro Propagation and Recovery of Eight Apple and Two Pear Cultivars Held in a Germplasm Bank. Am. J. Plant Sci. 2017, 8, 2238–2254. [Google Scholar] [CrossRef] [Green Version]
- de Araújo, R.C.; Rodrigues, F.A.; Nadal, M.C.; de Souza Ribeiro, M.; Antônio, C.A.C.; Rodrigues, V.A.; de Souza, A.C.; Pasqual, M.; Dória, J. Acclimatization of Musa spp. Seedlings Using Endophytic Bacillus spp. and Buttiauxella agrestis Strains. Microbiol. Res. 2021, 248, 126750. [Google Scholar] [CrossRef]
- Gilbert, S.; Poulev, A.; Chrisler, W.; Acosta, K.; Orr, G.; Lebeis, S.; Lam, E. Auxin-Producing Bacteria from Duckweeds Have Different Colonization Patterns and Effects on Plant Morphology. Plants 2022, 11, 721. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Rootstocks | IAA Production (mg L−1) |
|---|---|---|
| Acinetobacter septicus strain 23 * | ‘OH×F87′ | 19.48 |
| Buttiauxella sp. strain 27 * | ‘OH×F87′ | 10.82 |
| Acinetobacter ursingii strain 24 * | ‘PDW’ | 10.16 |
| Acinetobacter ursingii strain 25 * | ‘PDW’ | 8.99 |
| Acinetobacter ursingii strain 26 * | ‘OH ×F87′ | 8.60 |
| Acinetobacter ursingii strain 5 ** | ‘PDW’ | 7.61 |
| Acinetobacter ursingii strain 6 ** | ‘PDW’ | 6.50 |
| Acinetobacter ursingii strain 7 ** | ‘OH×F87′ | 5.55 |
| Acinetobacter ursingii strain 8 ** | ‘OH×F87′ | 5.16 |
| Acinetobacter ursingii strain 9 ** | ‘OH×F87′ | 4.00 |
| Acinetobacter ursingii strain 10 ** | ‘OH×F87′ | 3.44 |
| Acinetobacter ursingii strain 11 ** | ‘OH×F87′ | 2.89 |
| Acinetobacter ursingii strain 12 ** | ‘OH×F87′ | 2.45 |
| Acinetobacter ursingii strain 13 ** | ‘OH×F87′ | 2.06 |
| Acinetobacter ursingii strain 14 ** | ‘OH×F87′ | 1.78 |
| Acinetobacter ursingii strain 15 ** | ‘OH×F87′ | 1.39 |
| Acinetobacter ursingii strain 16 ** | ‘OH×F87′ | 1.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadal, M.C.; Ferreira, G.M.d.R.; Andrade, G.V.S.; Buttrós, V.H.; Rodrigues, F.A.; da Silva, C.M.; Martins, A.D.; Rufato, L.; Luz, J.M.Q.; Dória, J.; et al. Endophytic Bacteria Can Replace the Need for Synthetic Auxin during In Vitro Rooting of Pyrus communis. Agronomy 2022, 12, 1226. https://doi.org/10.3390/agronomy12051226
Nadal MC, Ferreira GMdR, Andrade GVS, Buttrós VH, Rodrigues FA, da Silva CM, Martins AD, Rufato L, Luz JMQ, Dória J, et al. Endophytic Bacteria Can Replace the Need for Synthetic Auxin during In Vitro Rooting of Pyrus communis. Agronomy. 2022; 12(5):1226. https://doi.org/10.3390/agronomy12051226
Chicago/Turabian StyleNadal, Michele Carla, Gustavo Magno dos Reis Ferreira, Gracielle Vidal Silva Andrade, Victor Hugo Buttrós, Filipe Almendagna Rodrigues, Caroline Marcela da Silva, Adalvan Daniel Martins, Léo Rufato, José Magno Queiroz Luz, Joyce Dória, and et al. 2022. "Endophytic Bacteria Can Replace the Need for Synthetic Auxin during In Vitro Rooting of Pyrus communis" Agronomy 12, no. 5: 1226. https://doi.org/10.3390/agronomy12051226
APA StyleNadal, M. C., Ferreira, G. M. d. R., Andrade, G. V. S., Buttrós, V. H., Rodrigues, F. A., da Silva, C. M., Martins, A. D., Rufato, L., Luz, J. M. Q., Dória, J., & Pasqual, M. (2022). Endophytic Bacteria Can Replace the Need for Synthetic Auxin during In Vitro Rooting of Pyrus communis. Agronomy, 12(5), 1226. https://doi.org/10.3390/agronomy12051226