
Carbon and Nitrogen Stable Isotope Abundance and Soil Stoichiometry of Zanthoxylum planispinum var. dintanensis Plantations of Different Ages


Abstract
:1. Introduction
2. Materials and Methods
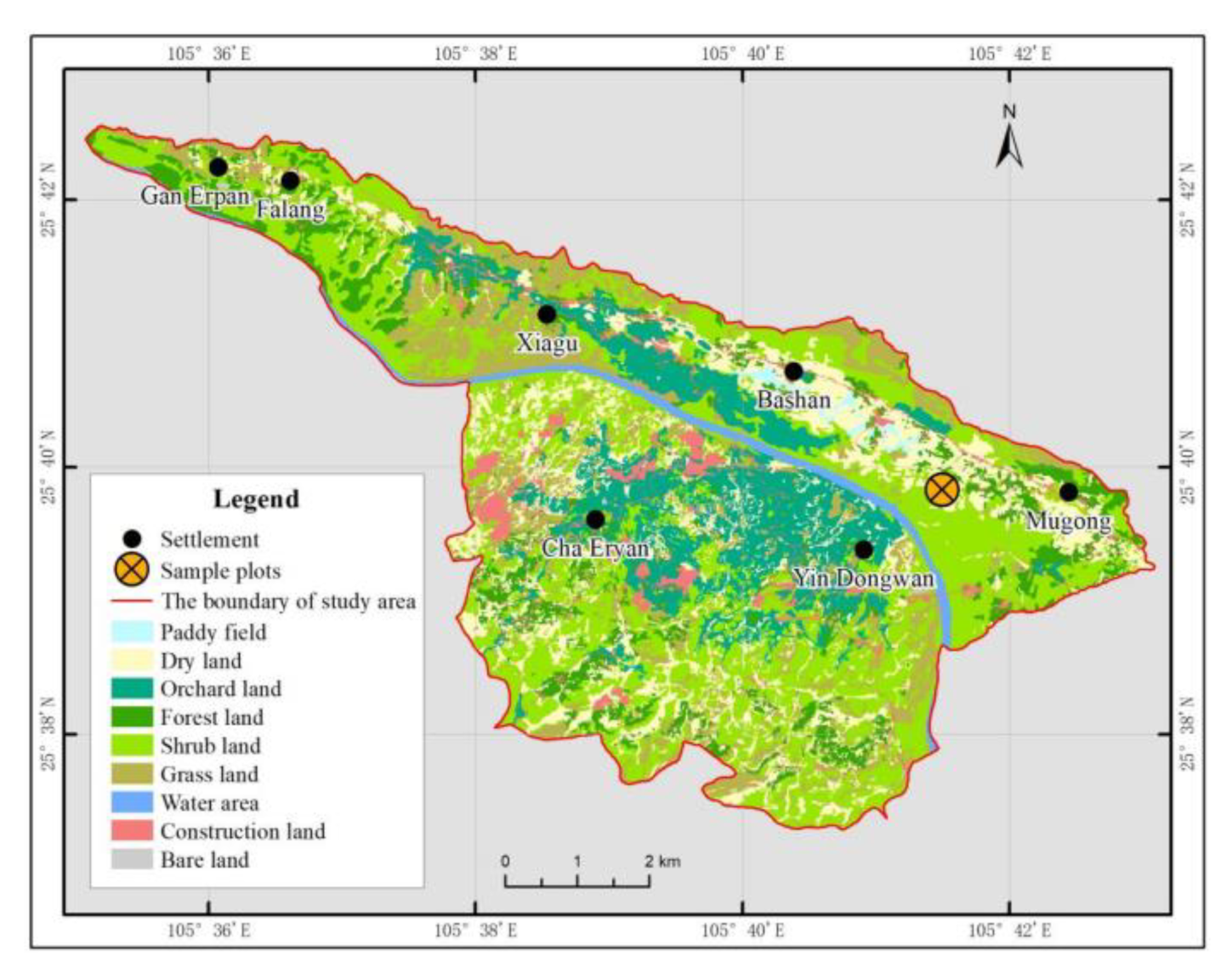
2.1. Overview of the Study Area
2.2. Experimental Design
2.2.1. Sample Plot Setting
2.2.2. Sample Collection
2.2.3. Sample Determination
2.2.4. Statistical Analysis
3. Results
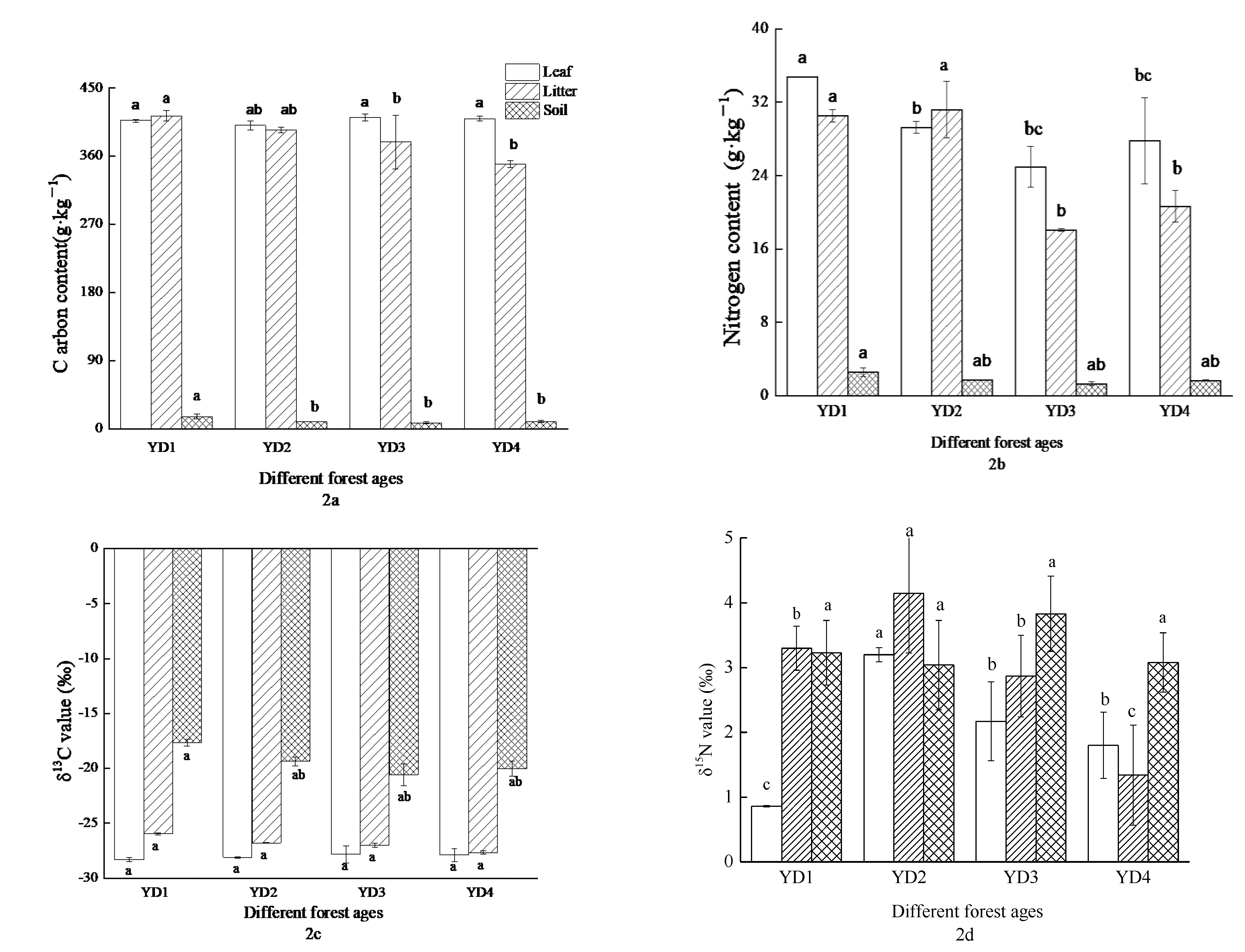
3.1. The Characteristics of Zanthoxylum planispinum var. dintanensis C, N, and Their Stable Isotopes from Different Aged Plantations
3.2. Relationship between Zanthoxylum planispinum var. dintanensis Plantation C, N and Their Stable Isotopes
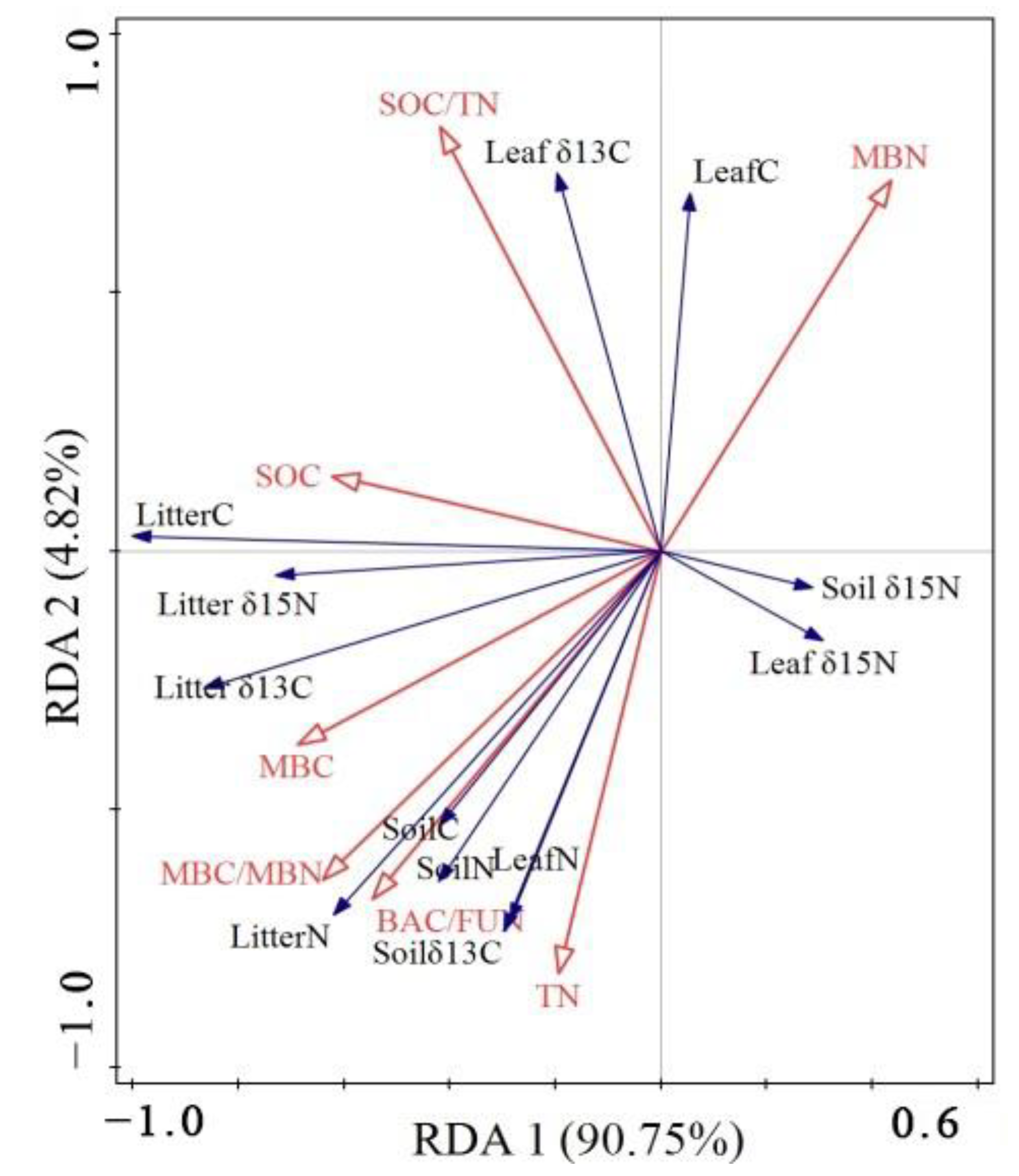
3.3. The Impact of Soil Stoichiometry on Plantation C, N, and Their Stable Isotopes
4. Discussion
4.1. The Abundance Characteristics of δ13C and δ15N in Zanthoxylum planispinum var. dintanensis Plantations of Different Plantation Age
4.2. The Driving Mechanism of Soil Stoichiometry to Plantation C and N Isotopes Fractionation
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, L.; Song, M.H.; Wang, S.Q.; Fan, J.W.; Liu, J.Y.; Zhong, H.P.; Yu, G.R.; Gao, L.P.; Hu, Z.M.; Chen, B.; et al. Patterns of soil 15N and total and their relationships with environmental factors on the Qinghai-Tibetan Plateau. Soil Sci. Soc. Chin. 2014, 24, 232–242. [Google Scholar] [CrossRef]
- Liu, X.Z.; Gao, C.C.; Su, Q.; Zhang, Y.; Song, Y. Altitudinal trends in δ13C value, stomatal density and nitrogen content of Pinnus tabuliformis needles on the southern slope of the middle Qinling Mountains, China. J. Mount. Sci. 2016, 13, 1066–1077. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.Y. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Vogel, J.C. Fractionation of the carbon isotopes during photosynthesis. Architect. Instit. Jpn. 1980, 3, 111–135. [Google Scholar]
- Lin, G.H. Stable Isotope Ecology; Higher Education Press: Beijing, China, 2013. [Google Scholar]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:Pstoichimetry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 39, 127–138. [Google Scholar] [CrossRef]
- Collins, J.G.; Dijkstra, P.; Hart, S.C.; Hungate, B.A.; Flood, N.M.; Schwartz, E. Nitrogen source influences natural abundance 15N of Escherichia coli. FEMS Microbiol. Lett. 2008, 282, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.A.; Jia, Y.F.; Li, W. Effects of environmental and biotic factors on carbon isotopic fractionation during decomposition of soil organic matter. Sci. Rep. 2015, 5, 11043. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cao, X.Q.; Sun, M.Y.; Li, P.X.; Teng, Z.; Xu, X.N. Foliar carbon and nitrogen stoichiometric trait and isotope discrimination of different Cunninghamia lanceolata foliar ages across an altitude gradient. Ecol. Environ. Sci. 2019, 28, 1776–1784. [Google Scholar]
- Tanaka-Oda, A.; Kenzo, T.; Koretsne, S.; Sasaki, H.; Fukuda, K. Ontogenetic changes in water-use efficiency (δ13C) and leaf traits differ among tree species growing in a semiarid region of the Loss Plateau, China. Forest Ecol. Manag. 2010, 259, 953–9757. [Google Scholar] [CrossRef]
- Song, L.N.; Zhu, J.J.; Yan, Q.L.; Li, M.C.; Yu, G.Q. Comparison of intrinsic water use efficiency between different aged Pinus sylvestris var. mongolica wide windbreaks in semiarid sandy land of northern China. Agroforest. Syst. 2015, 89, 477–489. [Google Scholar]
- Zheng, L.J.; Huang, Z.Q.; He, Z.M.; Wang, X.Y.; Liu, X.M.; Liu, R.Q.; Xiao, H.Y. Influence of forest and foliar ages on the composition of stable carbon and nitrogen isotope of Cunninghamia lanceolata in Subtropic China. Sci. Silv. Sin. 2015, 51, 22–28. [Google Scholar]
- Lucas-Borja, M.E.; Hedo, J.; Cerdá, A.; Candel-Pérez, D.; Vinegla, B. Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine (Pinus nigra Ar. ssp. Salzmannii) Forest. Sci. Total Environ. 2016, 562, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wang, K.; Tang, Z.S.; Shangguan, Z.P. Soil organic carbon dynamics following natural vegetation restoration: Evidence from stable carbon isotopes (δ13C). Agric. Ecosyst. Environ. 2016, 221, 235–244. [Google Scholar] [CrossRef]
- Zhao, Y.F.; Wang, X.; Ou, Y.S.; Jia, H.X.; Li, J.; Shi, C.M.; Liu, Y. Variations in soil δ13C with alpine meadow degradation on the eastern Qinghai-Tibet Plateau. Geoderma 2019, 338, 178–186. [Google Scholar] [CrossRef]
- Lorenz, M.; Derrien, D.; Zeller, B.; Udelhoven, T. The linkage of 13C and 15N soil depth gradients with C:N and O:C stoichiometry reveals tree species effects on organic matter turnover in soil. Biogeochemistry 2020, 151, 203–220. [Google Scholar] [CrossRef]
- Yang, L.; Xiong, K.N.; Xiao, S.Z.; Lan, J.C.; Ao, X.H. Carbon sequestration benefits of Zanthoxylum Bungeanum forest in karst rocky desertification areas. Bull. Soil Water Conserv. 2016, 36, 291–297. [Google Scholar]
- Li, A.D.; Yu, L.F.; Wei, X.L. Ecological Water Requirement of Zanthoxylum bungeanum Forestland in Typical Karst Valleys. Soil 2008, 40, 490–494. [Google Scholar]
- Qu, S.; Yang, R.; Wang, Y.; Pei, Y.D.; Li, K.F.; Hu, J.D. Change characteristics of soil nutrients during Zanthoxylum bungeanum var. dingtanensis grow process in karst plateau. Non-Wood For. Res. 2020, 38, 183–192. [Google Scholar]
- Zou, J.; Yu, L.F.; Huang, Z.S. Variation of leaf carbon isotope in plants in different lithological habitats in a karst area. Forest 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.H.; Chen, G.S.; Chen, F.S.; Zhao, Q.; Ji, X.Y. Foliar nutrients and resorption efficiencies in four Pinus sylvestris var. mongolica plantations of different ages on sandy soil. Sci. Silv. Sin. 2005, 41, 21–27. [Google Scholar]
- Yan, T.; Lü, X.T.; Yang, K.; Zhu, J.J. Leaf nutrient dynamics and nutrient resorption: A comparison between larch plantations and adjacent secondary forest in northeast, China. J. Plant Ecol. 2016, 9, 165–173. [Google Scholar]
- Farquhar, G.D.; Leary, M.O.; Berry, J. On the relationship between carbon isotope discrimination and the intercellular carbon isotope concentration in leaves. Austr. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Analysis, 3rd ed.; China Agricultural Publishing House: Beijing, China, 2000. [Google Scholar]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B.; Kou, D.; Liu, L.; Fang, K.; Zhang, B.B.; et al. LinKing microbial C:N:P stoichiometry to microbial community and abiotic factors along a 2500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar]
- Lin, X.G. Principles and Methods of Soil Microbial Research; Higher Education Press: Beijing, China, 2017. [Google Scholar]
- Chen, J.; Chang, S.X.; Anyia, A.O. The physiology and stability of leaf carbon isotope discrimination as a measure of water-use efficiency in barley on the Canadian prairies. J. Agron. Crop Sci. 2011, 197, 1–11. [Google Scholar]
- Heberling, J.M.; Fridley, J.D. Biogeographic constraints on the world-wide leaf economics spectrum. Glob. Ecol. Biogeogr. 2012, 21, 1137–1146. [Google Scholar]
- Kieckbusch, D.K.; Koch, M.S.; Serafy, J.E.; Anderson, W. Trophic linkages among primary producers and consumers in fringing mangroves of subtropical lagoons. Bull. Mar. Sci. 2004, 74, 271–285. [Google Scholar]
- Lee, S. Carbon dynamics of Deep Bay, eastern Pearl River estuary, China. I I: Trophic relationship based on carbon and nitrogen-stable isotopes. Mar. Ecol. Prog. Ser. 2000, 205, 1–10. [Google Scholar]
- Martinelli, L.; Almeida, S.; Brown, I.; Moreira, M.; Victoria, R.; Sternberg, L.; Ferreira, C.; Thomas, W. Stable carbon isotope ratio of tree leaves, boles and fine litter in a tropical forest in Rondonia, Brazil. Oecologia 1998, 114, 170–179. [Google Scholar]
- Peri, P.L.; Ladd, B.; Pepper, D.A.; Bonser, S.P.; Laffan, S.W.; Amelung, W. Carbon (δ13C) and nitrogen (δ15N) stable isotope composition in plant and soil in southern Patagonia’s native forests. Glob. Change Biol. 2012, 18, 311–321. [Google Scholar] [CrossRef]
- Balesdent, J.; Cirardin, C.; Mariotti, A. Site-related δ13C of tree leaves and soil organic matter in a temperate forest. Ecology 1993, 74, 1713–1721. [Google Scholar]
- Högberg, P.; Ekblad, A. Substrate-induced respiration measured in situ in a C3-plant ecosystem using additions of C4-sucrose. Soil Biol. Biochem. 1996, 28, 1131–1138. [Google Scholar] [CrossRef]
- Liao, J.D.; Boutton, T.W.; Jastrow, J.D. Organic matter turnover in soil physical fractions following woody plant invasion of grassland: Evidence from natural 13C and 15N. Soil Biol. Biochem. 2006, 38, 3197–3210. [Google Scholar] [CrossRef]
- Huang, Y.H.; Li, Y.L.; Xiao, Y.; Wenigmann, K.O.; Zhou, G.Y.; Zhang, D.Q.; Wenigmann, M.; Tang, X.L.; Liu, J.X. Controls of litter quality on the carbon sink in soils through partitioning the products of decomposing litter in a forest succession series in South China. Forest Ecol. Manag. 2011, 261, 1170–1177. [Google Scholar] [CrossRef]
- Buchmann, N.; Kao, W.Y.; Ehleringer, J. Influence of stand structure on carbon-13 of vegetation, soils, and canopy air within deciduous and evergreen forests in Utah, United States. Oecologia 1997, 110, 109–119. [Google Scholar] [CrossRef]
- Mariotti, A.; Pierre, D.; Vedy, J.C.; Bruckert, S.; Guillenmot, J. The abundance of natural nitrogen 150in the organic matter of soils along an altitudinal gradient. Catena 1980, 7, 293–300. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Buchmann, N.; Flanagan, L.B. Carbon isotope ratios in belowground carbon cycle processes. Ecol. Appl. 2000, 10, 412–422. [Google Scholar] [CrossRef]
- Tsialtas, J.T.; Handley, L.; Kassioumi, M.T.; Veresoglou, D.S.; Gagianas, A.A. Interspecific variation in potential water- use efficiency and its relation to plant species abundance in a water-limited grassland. Funct. Ecol. 2001, 15, 605–614. [Google Scholar] [CrossRef]
- Zhang, J.; Gu, L.; Cao, Y.; Hao, Y.; He, J.; Li, J.; Li, Y.; Ren, Y.; Wang, F.; Wu, F.; et al. Nitrogen control of 13C enrichment in heterotrophic organs relative to leaves in a landscape-building desert plant species. Biogeosciences 2015, 12, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, C.; Arndt, S.K.; Livesley, S.J.; Edgar, A.C.; White, D.A.; Adams, W.A.; Eamus, D. Estimation of leaf area index in eucalypt forest with vertical foliage, using cover and fullframe fisheye photography. Forest Ecol. Manag. 2007, 242, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Diefendorf, A.F.; Mueller, E.; Wing, S.L.; Koch, P.L.; Freeman, K.H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc. Natl. Acad. Sci. USA 2010, 107, 5738–5743. [Google Scholar] [CrossRef] [Green Version]
- Li, D.J.; Niu, S.L.; Luo, Y.Q. Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: A metanalysis. New Phytol. 2012, 195, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformation. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister, B.S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Houlton, B.Z.; Liu, D.W.; Hou, J.F.; Cheng, W.X.; Bai, E. Stable isotopic constraints on global soil organic carbon turnover. Biogeosciences 2018, 15, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.A.; Partt, R.L.; Schipper, L.A.; Baisden, W.T.; Mudge, P. Relationship between soil 15N, C/N and N losses across land uses in New Zealand. Agric. Ecosyst. Environ. 2010, 139, 736–741. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Carreiro, M.M.; Repert, D.A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry 2002, 60, 1–24. [Google Scholar] [CrossRef]
- Arunachalam, K.; Arunachalam, A.; Melkania, N.P. Influence of soil properties on microbial populations, activity and biomass in humid subtropical mountainous ecosystems of India. Biol. Fertil. Soil 1999, 30, 217–223. [Google Scholar] [CrossRef]
- Nel, J.A.; Craine, J.M.; Cramer, M.D. Correspondence between δ13C and δ15N in soils suggests coordinated fraction processes for soil C and N. Plant Soil 2018, 423, 257–271. [Google Scholar] [CrossRef]
- Billings, S.A.; Richter, D.D. Changes in stable isotopic of soil nitrogen and carbon during 40 years of forest development. Oecologia 2006, 148, 325–333. [Google Scholar] [CrossRef]
- De Rouw, A.; Soulileuth, B.; Huon, S. Stable carbon isotope rations in soil and vegetation shift with cultivation practices (Northern Laos). Agtric. Ecosyst. Environ. 2015, 200, 161–168. [Google Scholar] [CrossRef]
- Wynn, J.C.; Bird, M.I. Environmental controls on the stable carbon isotopic composition of soil organ C3 and C4 plants, Australia. Tellus Ser. B Chem. Phys. Meteorol. 2008, 60, 604–621. [Google Scholar] [CrossRef] [Green Version]
- Wynn, J. Carbon isotope fractionation during decomposition of organic matter in soil and paleosols: Implication for palaeoecological interpretation of paleosols. Palaeogeograph. Palaeoclimatol. Palaeoecol. 2007, 251, 437–448. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plot No | Age | Average Crown Width/m | Height/m | Vegetation Coverage/% | Density/ ikaika (Plant/hm2) | Yield (Plant/Kg) |
|---|---|---|---|---|---|---|
| YD1 | 5–7 | 2.5 × 3 | 2.7 | 100 | 1150 | 6–7 |
| YD2 | 10–12 | 2.5 × 3 | 2.7 | 100 | 1150 | 7–8 |
| YD3 | 20–22 | 3.5 × 3 | 3.5 | 90 | 1000 | 4–5 |
| YD4 | 30–32 | 4 × 5 | 4 | 75 | 650 | 1–1.5 |
| Index | C leaf | δ13C leaf | N leaf | δ15N leaf | C litter | δ13C litter | N litter | δ15N litter | C soil | δ13C soil | N soil |
|---|---|---|---|---|---|---|---|---|---|---|---|
| δ13C leaf | 0.225 | 1 | |||||||||
| N leaf | −0.108 | −0.751 * | 1 | ||||||||
| δ15N leaf | −0.584 | 0.044 | −0.474 | 1 | |||||||
| C litter | −0.043 | 0.219 | 0.260 | −0.307 | 1 | ||||||
| δ13C litter | −0.100 | −0.244 | 0.624 | −0.429 | 0.855 ** | 1 | |||||
| N litter | −0.392 | −0.389 | 0.706 | −0.021 | 0.595 | 0.676 | 1 | ||||
| δ15N litter | −0.205 | 0.210 | 0.007 | 0.275 | 0.727 * | 0.575 | 0.606 | 1 | |||
| C soil | −0.132 | −0.545 | 0.792 * | −0.614 | 0.400 | 0.666 | 0.534 | −0.059 | 1 | ||
| δ13C soil | −0.193 | −0.704 | 0.939 ** | −0.384 | 0.271 | 0.666 | 0.724 * | 0.167 | 0.747 * | 1 | |
| N soil | −0.280 | −0.528 | 0.774 * | −0.484 | 0.400 | 0.648 | 0.628 | 0.041 | 0.976 ** | 0.778 * | 1 |
| δ15N soil | −0.180 | −0.485 | 0.054 | 0.041 | −0.286 | −0.019 | −0.369 | −0.484 | 0.181 | −0.045 | 0.074 |
| Sorting Axis | Axis 1 | Axis 2 | Axis 3 | Axis 4 |
|---|---|---|---|---|
| Explains | 90.75 | 4.82 | 1.79 | 0.54 |
| Pseudo-canonical correlation | 0.9937 | 0.9975 | 0.8307 | 0.9781 |
| Explained variation (cumulative) | 90.75 | 95.57 | 97.35 | 97.89 |
| Explained fitted variation (cumulative) | 92.65 | 97.57 | 99.36 | 99.34 |
| Index | Order of Importance | Explains/% | F | p |
|---|---|---|---|---|
| MBC | 1 | 44.1 | 4.7 | 0.072 |
| BAC/FUN | 2 | 17.6 | 3.0 | 0.144 |
| SOC/TN | 3 | 15.1 | 1.8 | 0.234 |
| MBN | 4 | 8.6 | 4.2 | 0.218 |
| SOC | 5 | 6.7 | 1.2 | 0.368 |
| TN | 6 | 5.8 | 1.1 | 0.416 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Wu, Y.; Song, Y.; Li, Y. Carbon and Nitrogen Stable Isotope Abundance and Soil Stoichiometry of Zanthoxylum planispinum var. dintanensis Plantations of Different Ages. Agronomy 2022, 12, 1248. https://doi.org/10.3390/agronomy12061248
Yu Y, Wu Y, Song Y, Li Y. Carbon and Nitrogen Stable Isotope Abundance and Soil Stoichiometry of Zanthoxylum planispinum var. dintanensis Plantations of Different Ages. Agronomy. 2022; 12(6):1248. https://doi.org/10.3390/agronomy12061248
Chicago/Turabian StyleYu, Yanghua, Yingu Wu, Yanping Song, and Yitong Li. 2022. "Carbon and Nitrogen Stable Isotope Abundance and Soil Stoichiometry of Zanthoxylum planispinum var. dintanensis Plantations of Different Ages" Agronomy 12, no. 6: 1248. https://doi.org/10.3390/agronomy12061248
APA StyleYu, Y., Wu, Y., Song, Y., & Li, Y. (2022). Carbon and Nitrogen Stable Isotope Abundance and Soil Stoichiometry of Zanthoxylum planispinum var. dintanensis Plantations of Different Ages. Agronomy, 12(6), 1248. https://doi.org/10.3390/agronomy12061248