1. Introduction
Spearmint (Mentha spicata L.), is a creeping rhizomatous, glabrous and herbaceous perennial plant, with a pungent smell. The commercial interest in spearmint essential oil, which is among the 10 most commercialized in the world, has increased the demand in mint cultivation. Spearmint indeed contributes greatly to the economy due to its extensive use in food, perfumery, toothpaste, the pharmaceutical industry and the production of essential oil (EO).
The EO of spearmint has antimicrobial [
1], insecticidal [
2], larvicidal [
3], antioxidant [
4] and mutagenic activity due to the abundance of terpenes [
5].
The crop needs sufficient water availability during summer months to ensure good growth and high yields. In spearmint, sustained deficit irrigation, reducing water availability, clearly decreased mint hay yields and affected oil yield and quality [
3,
6,
7].
The positive effect of water availability on herb yields of various mint species and aromatic grasses has been reported [
8,
9,
10]. Meanwhile, in medicinal plants, a moderate water-stress condition, while reducing the accumulation of dry matter, increases the yield of essential oil and modifies its composition in respect to the well-watered plants [
8,
11,
12,
13].
Severe water deficit limits crop growth and yield with negative economic consequences [
14]. In the last few years, rainfall has declined in some parts of the Mediterranean region due to a climatic change [
15] causing substantial shortage in the available water resources for agricultural production. Furthermore, the uneven distribution of rainfall has become the main limitation to sustainable crop yields in drought-prone areas [
16]. Thus, the improved utilization of limited seasonal rainfall resources through reduction in soil water loss and increased water-utilization efficiency is needed.
A number of proven mulching techniques exist to entrap and conserve rainwater, such as plastic film mulch, straw mulch and ridge-furrow mulch [
17]. The use of plastic film mulch, however, leaves a large amount of plastic in the field, which has obvious negative effects on the environment and soil structure [
18].
Instead, the application of straw in agriculture will improve the agricultural ecological environment, because the need for open-field straw burning will be reduced or eliminated. Straw mulch has also been shown to reduce the needed quantities of chemical fertilizers [
19] as it increases soil organic matter and improves soil structure [
20].
Organic mulching plays a vital role in sustainably conserving soil moisture under low water availability in a manner that enhances crop yields and water-use efficiency [
16,
21]. Furthermore, conserving moisture through mulching and reduction in the frequency of water supplies may subsequently reduce the losses of N through leaching and minimize NH
3 volatilization by restricting soil temperature rise.
In the present study, we investigated the effect of straw mulch on rainfed spearmint (Mentha spicata L.) in environmental conditions typical of the Mediterranean region. In particular, we focused on plant growth, photosynthesis and oil yield. We hypothesized that mulch is useful and necessary to support plant growth and activity, and to avoid oil-yield reduction in spearmint.
2. Materials and Methods
2.1. Experimental Site
The study was conducted during 2020 and 2021 growing season at an experimental field site in Baranello (Molise Region, Italy, latitude 41°31′ N, longitude 14°33′ E, altitude 630 m a.s.l.). The area has an average annual rainfall of 845 mm and mean annual temperature of 12.7 °C. Molise Region is positioned on the eastern side of the Apennines watershed, and has a typical Mediterranean climate of southern Italy.
Before fertilizer applications, soil samples (0–30 cm) were taken from each block and analyzed according to standard procedures [
8]. The soil was characterized by a clay-sand texture (45% clay, 10% silt and 45% sand) and the organic-matter content was 1.5%. The soil profile was overall uniform, containing medium amount of total N (nitrogen, 0.11%), medium amount of available P (phosphorous, 19.5 µg g
−1) and medium quantity of exchangeable K (potassium, 133 µg g
−1). Soil had very low active CaCO
3, and pH was average neutral; salinity was low. Bean (
Phaseolus vulgaris L.) was the previous crop within both years.
2.2. Plant Material and Experimental Design
Ten-month old spearmint seedlings were transplanted during mid-April in both years, followed by a light irrigation after planting in rows 50 cm apart (plant density of 10 plants m−2). The field was surrounded by a buffer strip to allow for uniform growing conditions.
The experiment was arranged in a randomized block design with five replications (5 m2 each plot). After tillage was completed, the land was smoothed and firmed using a roller harrow.
The recommended doses of N (100 kg ha
−1), P
2O
5 (70 kg ha
−1) and K
2O (100 kg ha
−1), were applied before planting [
22].
The following treatments were designed: plants cultivated under natural rainfed conditions without mulch (hereafter, R plants), plants cultivated under natural rainfed conditions with straw mulch (hereafter, RM plants), plants grown under well-watered conditions, considered as control (hereafter, W plants) and plants grown under well-watered conditions with straw mulch (hereafter, WM plants). Within the straw mulch treatments, the wheat straw covered all the soil surface at the rates of 1 kg m−2 (5 cm straw length). The wheat straw was applied annually. Plants cultivated under well-watered conditions served as a control.
In the control plots, the crop evapotranspiration (ET) was fully restored by drip irrigation. After plant establishment (at 10 May, in both years), irrigation was withheld in R and RW plots [0 days after treatment (DAT)]. Briefly, potential evapotranspiration was calculated from micrometeorological data using the Penman–Monteith formula [
23]. Weeds were manually controlled.
2.3. Meteorological Measurements and Yield Biomass
Main meteorological parameters were measured by means of a standard weather station, located 20 m from the experimental field. The daily mean temperature ranged between 9.1–23.5 °C, while the corresponding minimum and maximum temperatures were 7.6 and 34.7 °C, respectively. Only 10.3 mm and 11.5 mm rainfall were received during the entire spearmint growth season in the years 2020 and 2021, respectively.
The crop was harvested manually on 0, 20, 40, 60 and 80 DAT in both years. The herbal-yield biomass was weighed, after which the plant samples were heated at 105 °C for one hour and then dried at 75 °C to constant weight.
2.4. Photosynthetic Gas Exchange
Gas-exchange activities, as net photosynthesis (Pn, µmol m
−2 s
−1) and stomatal conductance (g
s, mol m
−2 s
−1) were measured using a portable photosynthesis system, at ambient CO
2 concentration and at time of maximum solar radiation (e.g., 12:00 to 2:00 pm) as described by Delfine [
24]. Measurements were recorded on five fully expanded leaves selected randomly on different plants. For photosynthesis measurements, the leaf temperature was maintained at 30 °C as in the environmental conditions. Mesophyll conductance (g
m, mol m
−2 s
−1) of CO
2 was determined as described by Delfine [
8]. Briefly, the electron-transport rate calculated by gas exchange (Jc) and that measured by chlorophyll fluorescence (Jf) were compared at an irradiance of 800 µmol m
−2 s
−1 for accurate measurements of intercellular CO
2 concentration.
2.5. Relative Water Content
The relative water content (RWC) of leaves was measured according to Di Stasio [
25]. The third fully expanded leaves from the top were measured. The fresh leaves were weighed (FW), then submerged in distilled water for 12 h to obtain turgid weight (TW). Finally, the leaves were kept at 105 °C for 24 h until a constant weight was reached (DW). The formula RWC (%) = (FW − DW)/(TW − DW) × 100 was used to calculate the relative leaf water content.
2.6. Soil Water Content
The water content to a depth of 1.0 m was measured every 20 days from the beginning of treatment (DAT) to observe the dynamics in soil water content of the 0–0.3 and 0.3–1.0 m layers. The soil was sampled at 0.1 m intervals by manual coring between adjacent plants in the same row, and the gravimetric water content of the soil samples was calculated on the basis of oven-dried weights (dried at 105 °C).
2.7. Essential-Oil Analysis
Essential oil was extracted by hydrodistillation of fresh samples through Clevenger’s apparatus. The steam distillation was performed for 60 min. The separated spearmint EO collected was dried over anhydrous sodium sulfate and stored in dark-brown vials at 4 °C until used. Three extractions were performed for each treatment [
26]. Samples from every treatment and replicate were collected and analyzed separately for oil composition.
The leaf and stem dry biomass, RWC, photosynthesis, stomatal and mesophyll conductance, essential-oil content and essential-oil yield were recorded with interval of 20 days from 0 DAT.
2.8. GC and GC–MS Analysis
Analytical gas chromatography was carried out on a Perkin–Elmer Sigma 115 gas chromatograph fitted with a HP-5 MS capillary column (30 m × 0.25 mm), 0.25 µm film thickness. Helium was the carrier gas (1 mL/min). Column temperature was initially kept at 40 °C for 5 min, then gradually increased to 250 °C at 2 °C/min rate, held for 15 min and finally raised to 270 °C at 10 °C/min. Diluted samples (1/100 v/v, in n-pentane) of 1 µL were injected at 250 °C, manually and in the splitless mode. Flame-ionization detection (FID) was performed at 280 °C. Analysis was also run by using a fused silica HP Innowax polyethylenglycol capillary column (50 m × 0.20 mm), 0.20 µm film thickness and operating as described above. GC–MS analysis was performed on an Agilent 6850 Ser. II apparatus, fitted with a fused silica DB-5 capillary column (30 m × 0.25 mm), 0.33 µm film thickness, coupled to an Agilent Mass Selective Detector MSD 5973; ionization voltage 70 eV; electron-multiplier energy 2000 V. Gas chromatographic conditions were as given; transfer line temperature, 295 °C.
2.9. Identification of Compounds
Most constituents were identified by gas chromatography by comparison of their retention indices (Ki) with those of the literature [
27,
28] or with those of authentic compounds available in our laboratories. The retention indices were determined in relation to a homologous series of n-alkanes (C8–C24) under the same operating conditions. Further identification was made by comparison of their mass spectra on both columns with those stored in NIST 02, Wiley 275 libraries and our home-made library or with mass spectra from literature [
27,
29]. Component relative percentages were calculated based on GC peak areas without using correction factors. The primary oil components that were compared were limonene, carvone, 1,8-cineole and myrcene.
2.10. Data Statistics
All data were presented as means of the five replicates, while the data for essential oil content and yield and primary oil components are presented as means of the three replicates. SPSS 18.0 was used to conduct a one-way analysis of variance, and significant differences between treatments were compared by the least-significant-difference test at p < 0.05.
3. Results
As the measured parameters did not show significant differences between growing seasons, data averaged over 2 years, 2020 and 2021, were presented.
3.1. Plant Phenology and Soil Water Contents
No differences in phenology were observed between the spearmint plants exposed to the different treatments up to 40 DAT. Specifically, at the beginning of the study (0 DAT) the plants, of all treatments, were in the pre-flowering stage. After 20 DAT, the plants were in early bloom, and at 40 DAT in full bloom, in all four overall treatments. At 60 DAT the W, WM and RM plants were at the end of flowering and at 80 DAT in flower withering; while the R plants were already withering from 60 DAT until the end of the cropping cycle (80 DAT).
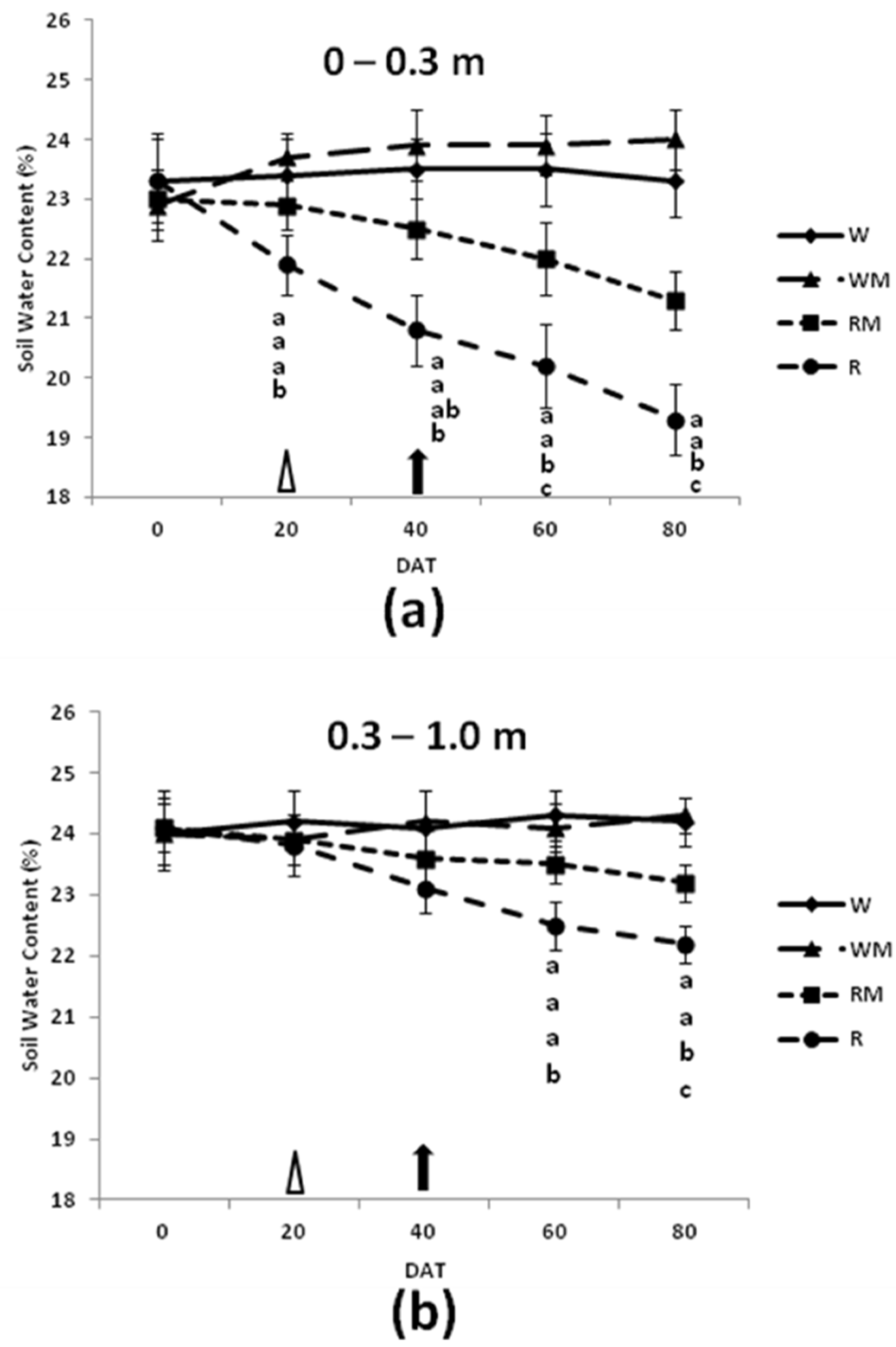
The soil water contents (SWC) (
Figure 1) showed that the rainfed mulching treatments increased the SWC within the 0–0.3 m soil profile compared with the rainfed treatment during the growing seasons of spearmint plants.
At the end of the spearmint growth stage (flower-withering stage, 80 DAT), the SWC within the 0–0.3 m soil profile under the rainfed treatments was lower in comparison to the RM and control treatment. The SWC reductions indicated that under the mulching treatments, more water was stored within the 0–0.3 m soil profile compared with the rainfed treatment. Over the studied cropping cycle (0–80 DAT), there were no significant differences in SWC between well-watered and well-watered mulching treatments. The amplitude of the variation was much larger in the 0–0.3 m than the 0.3–1.0 m layer.
3.2. Biomass Yields
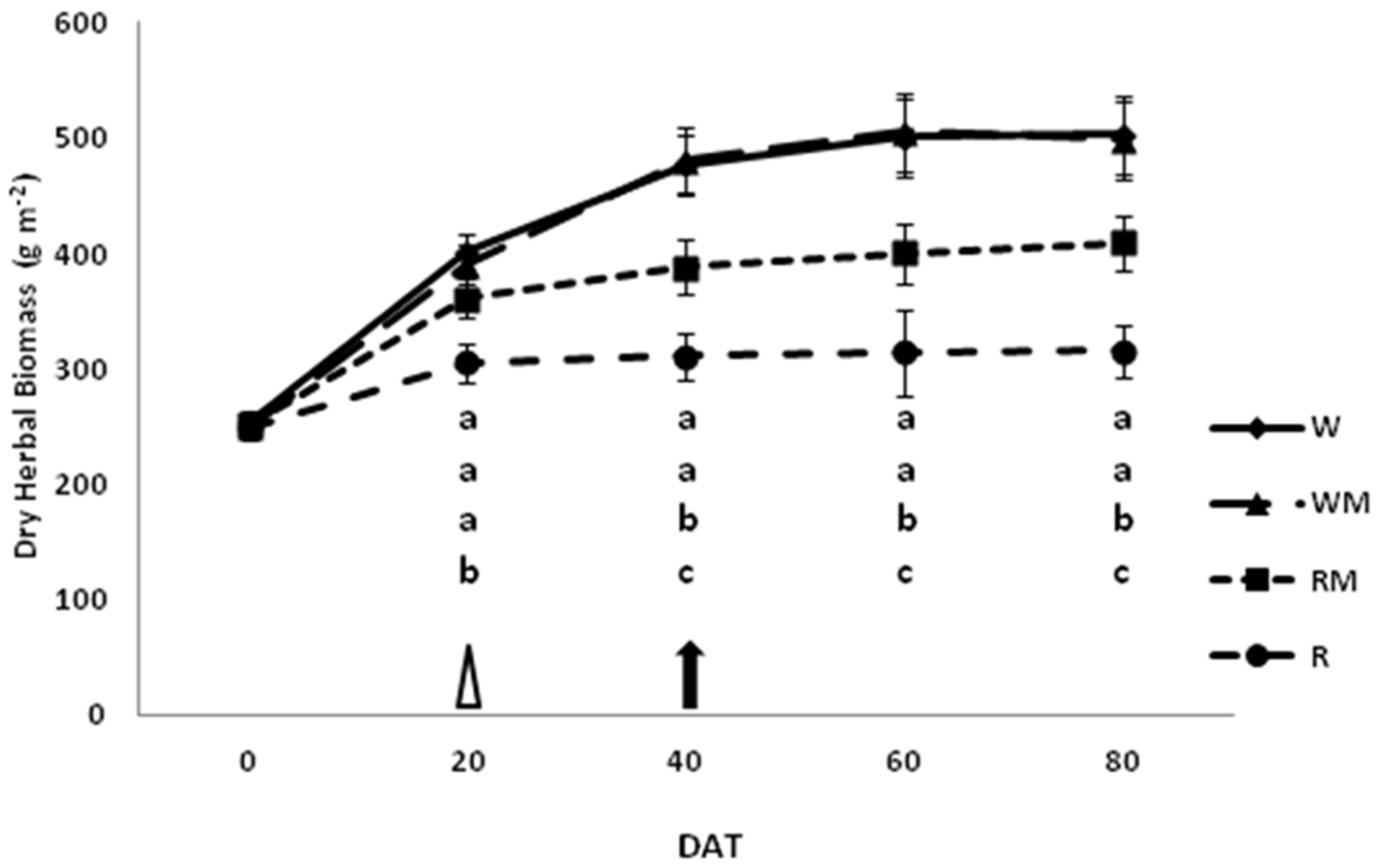
The dry herbal biomass yields of spearmint were significantly affected by the straw mulch treatment during the growth season (
Figure 2).
In W and WM plants, the dry herbal biomass content had increased over time, showing the maximum yield in full bloom (DAT 40) and no significant difference between the two treatments. The herbal yield of RM-treated plants tended to increase but significantly less than in WM plants, and the dry herbal biomass was about 20% less at the end of the experiments (DAT 80). Instead, the development of R plants stopped at the beginning of flowering, showing a reduction of about 38% at the end of the experiments compared to the control (W).
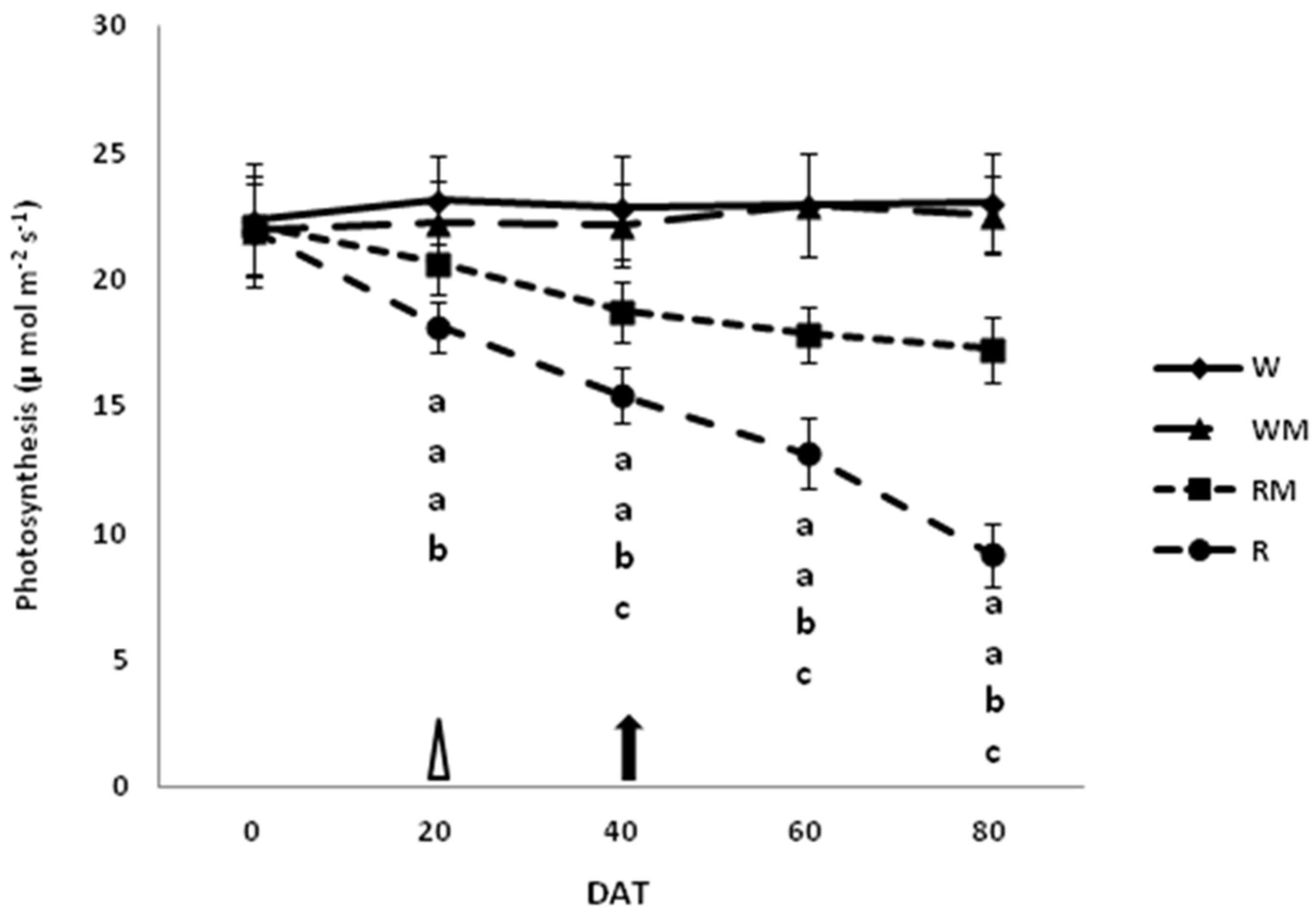
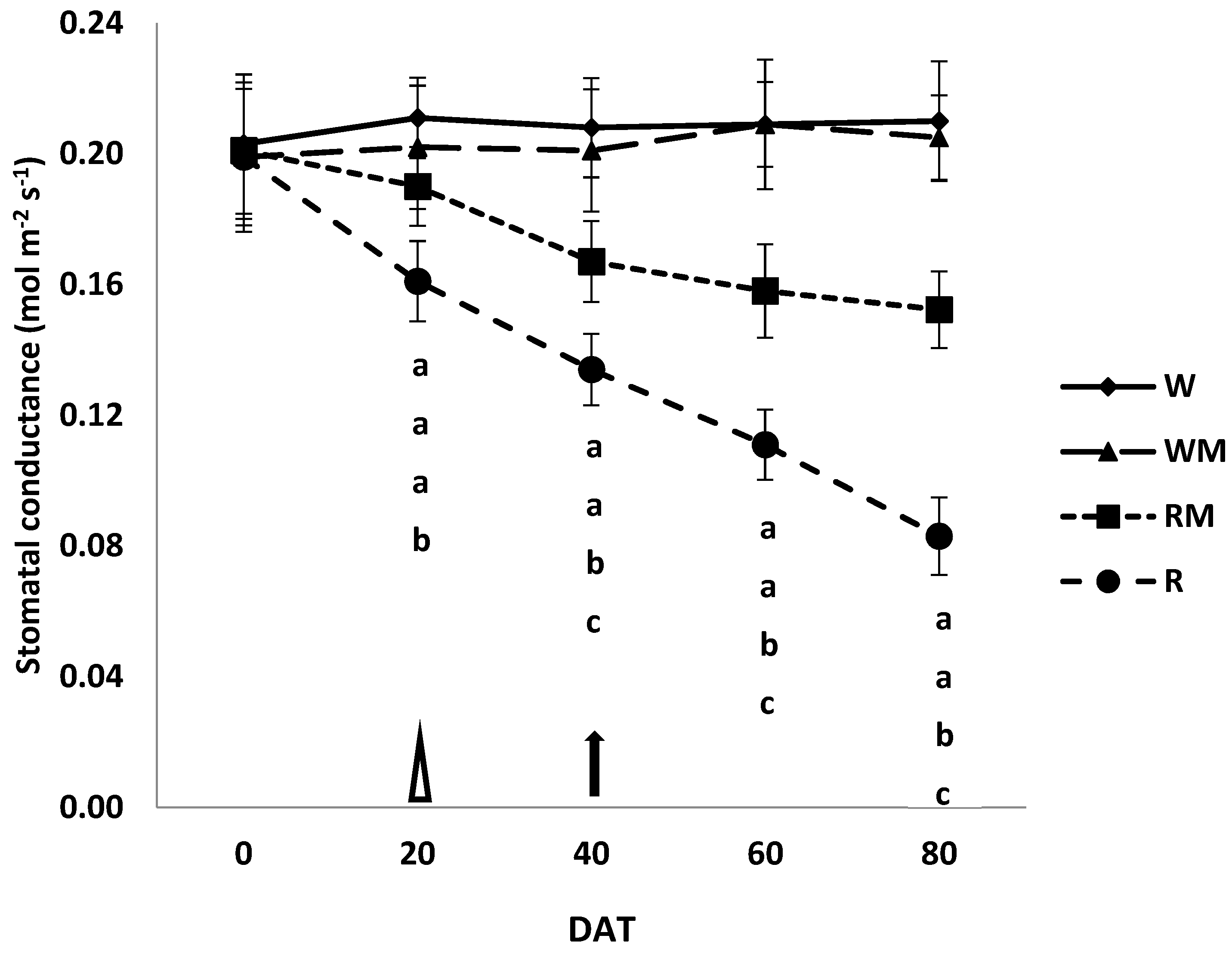
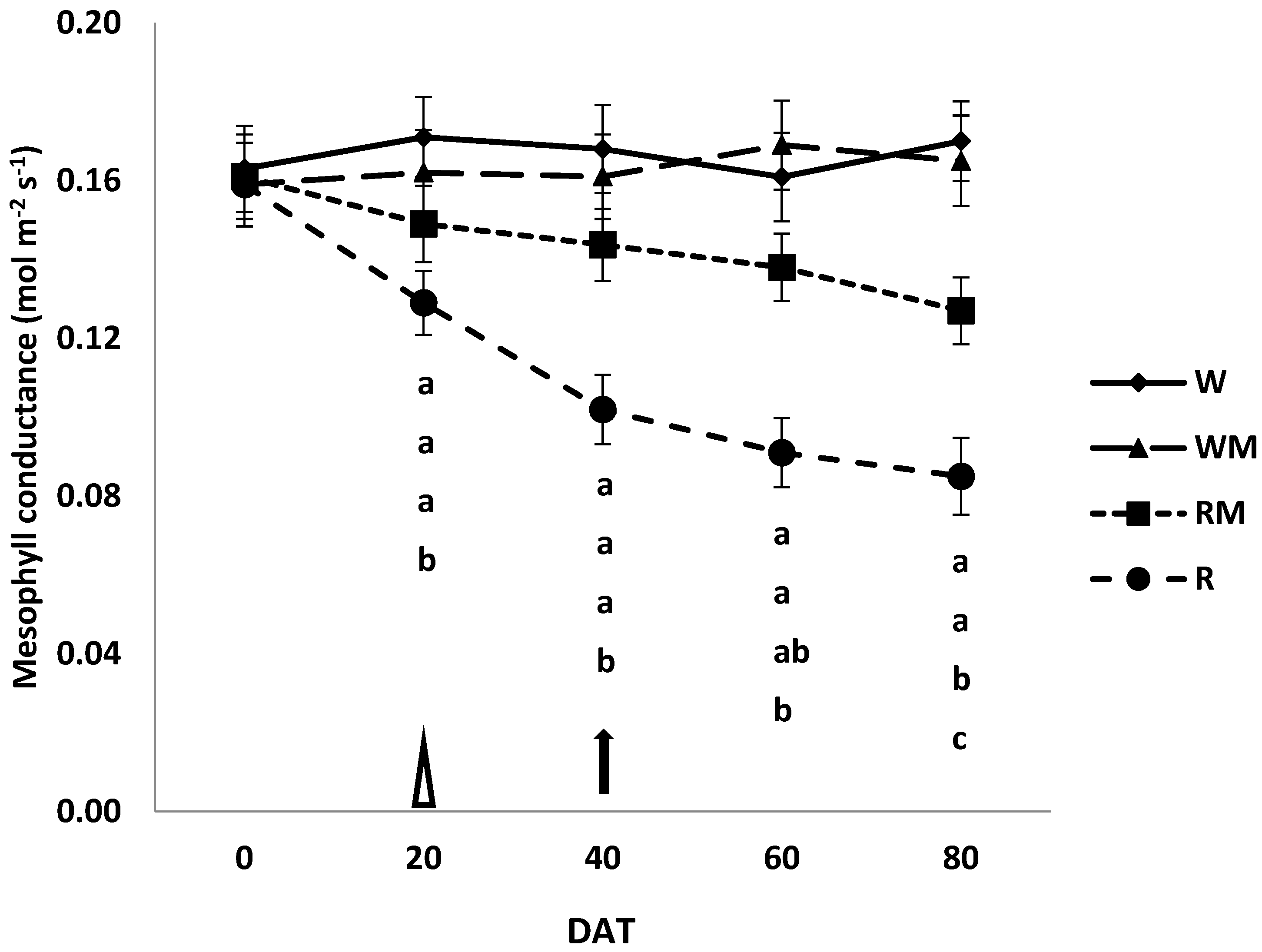
3.3. Photosynthesis, Stomatal and Mesophyll Conductances
The trends in photosynthesis (Pn), stomatal (g
s) and mesophyll (g
m) conductances among the different treatments are illustrated in
Figure 3,
Figure 4 and
Figure 5, respectively. The mean photosynthesis was 60% and 26% lower in R and RM plants compared to the corresponding controls (W and WM), respectively, at DAT 80. There were no significant differences in photosynthesis between the W and WM control plants over the studied cropping cycle (
Figure 3).
The same trend was observed for stomatal conductance (
Figure 4).
At the 20 DAT, mesophyll conductance was significantly lower only in the leaves of R plants with respect to control W (
Figure 5).
On the contrary, the mesophyll conductance of the RM treatments decreased and was significantly lower (p < 0.05), with respect to control WM, only at the 80 DAT. Over the studied treatment cycle, the mesophyll conductance of control W was similar to those of plants grown under well-watered conditions with straw mulch (WM).
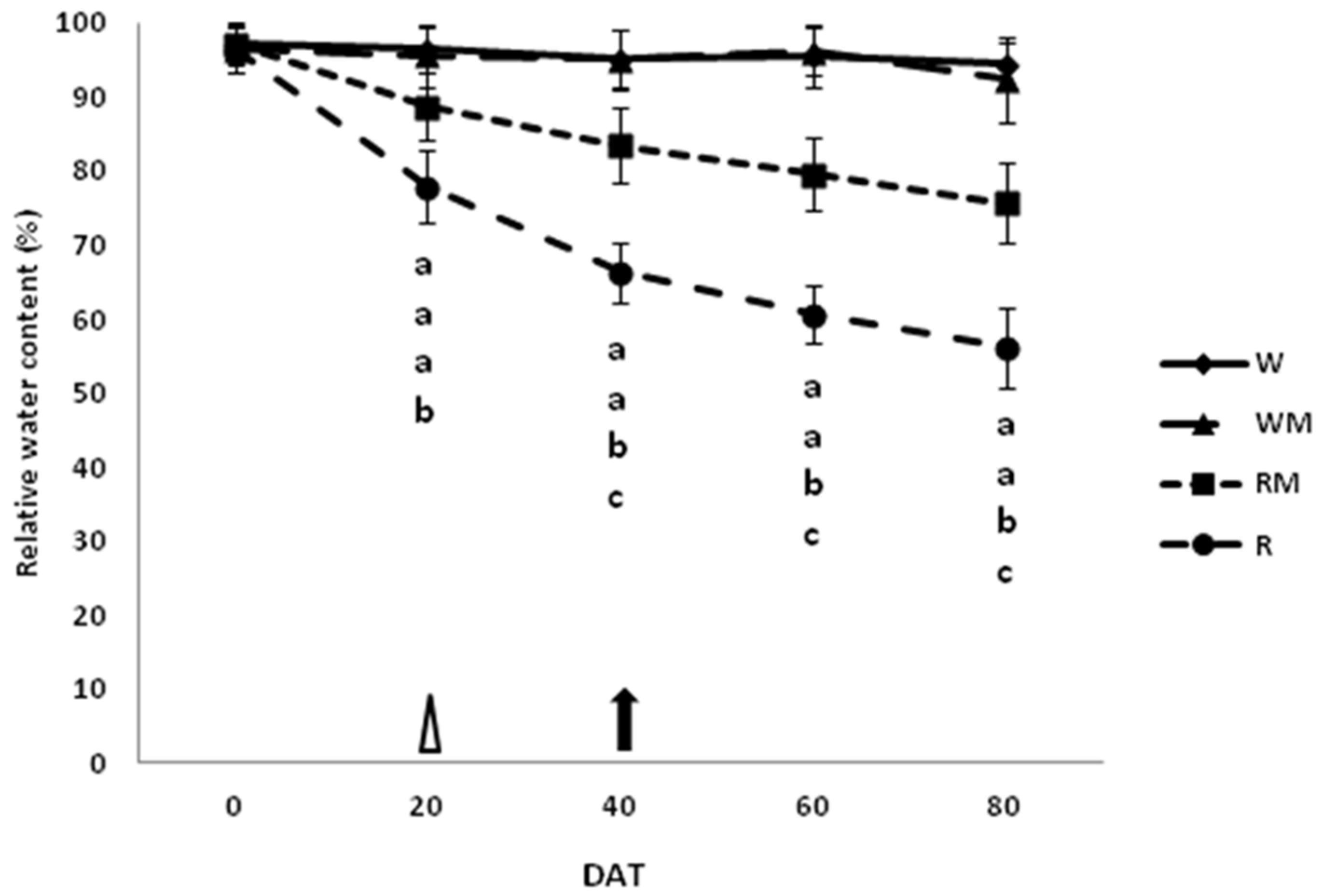
3.4. Relative Water Content
The results of the RWC indicated that RM plants generally had preserved tissue water content compared to R plants over the growing seasons (
Figure 6).
At the end of the spearmint growth stage (flower-withering stage, 80 DAT), the average RWC was significantly (
p < 0.05) lower in R and RM plants, by about 40% and 20%, respectively, in comparison to the control W and WM plants. No significant differences in RWC between the W and WM control plants over the cropping cycle were observed (
Figure 6).
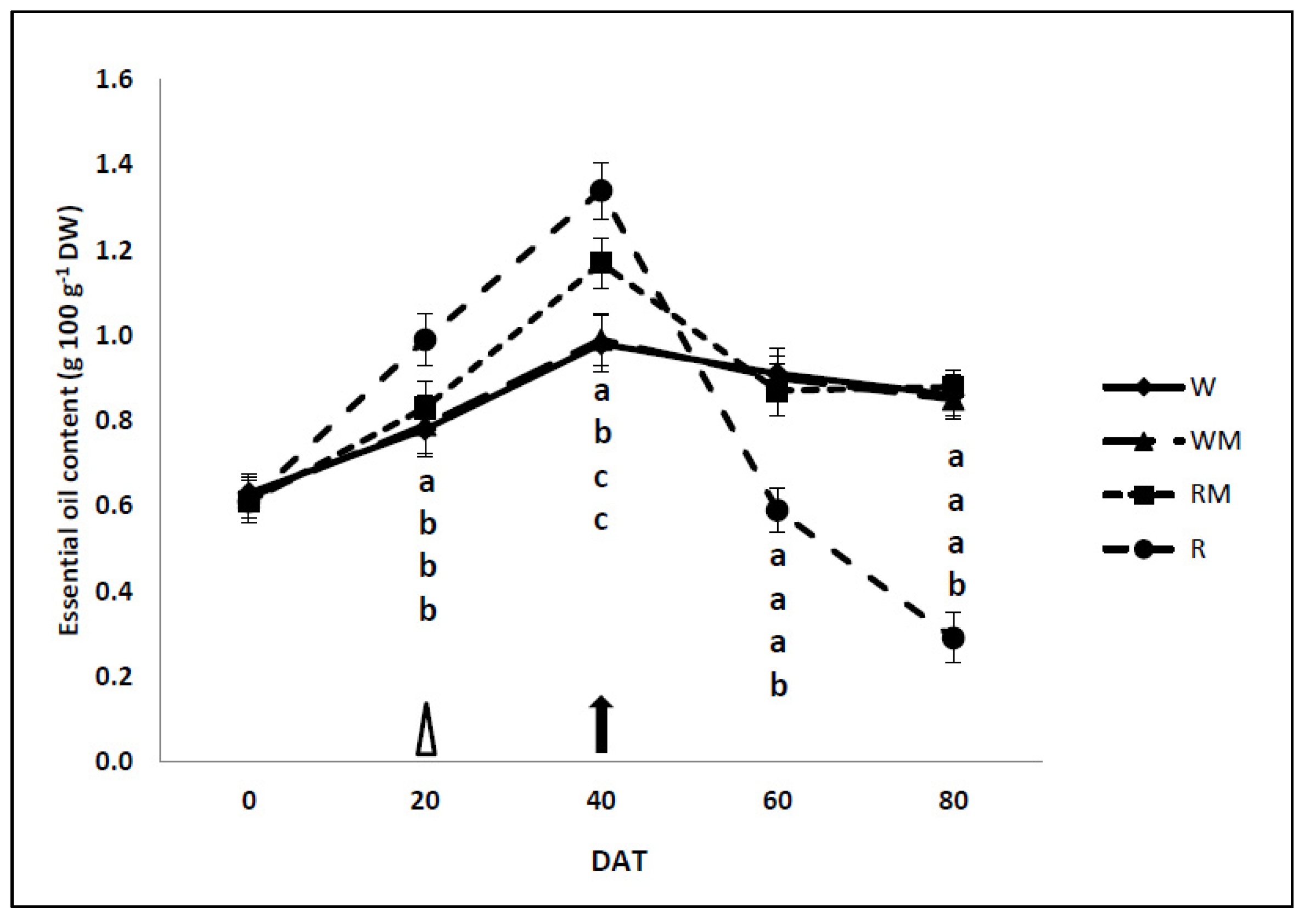
3.5. Essential-Oil Content and Essential-Oil Yield
The mulching treatments increased the EO content (+16%) in the RM compared to WM plants during the full bloom (40 DAT) (
Figure 7).
There were no differences in EO content (EOC) between the W and WM plants over the cropping cycles. During the full-bloom stages (40 DAT), the EO content of the W (−26%), WM (−26%) and RM (−14%) plants were significantly lower in comparison to R plants. While EO content was strongly reduced in R plants (about-77%) compared to W plants at the last harvest (80 DAT). RM loses a negligible quantity of oils compared to WM. At 80 DAT, EO content in W and WM plants dropped by about 5% compared to full bloom (40 DAT). EOC of RM dropped by 3%, while in R plants EOC decreased by 51%, almost disappearing in leaf tissues.
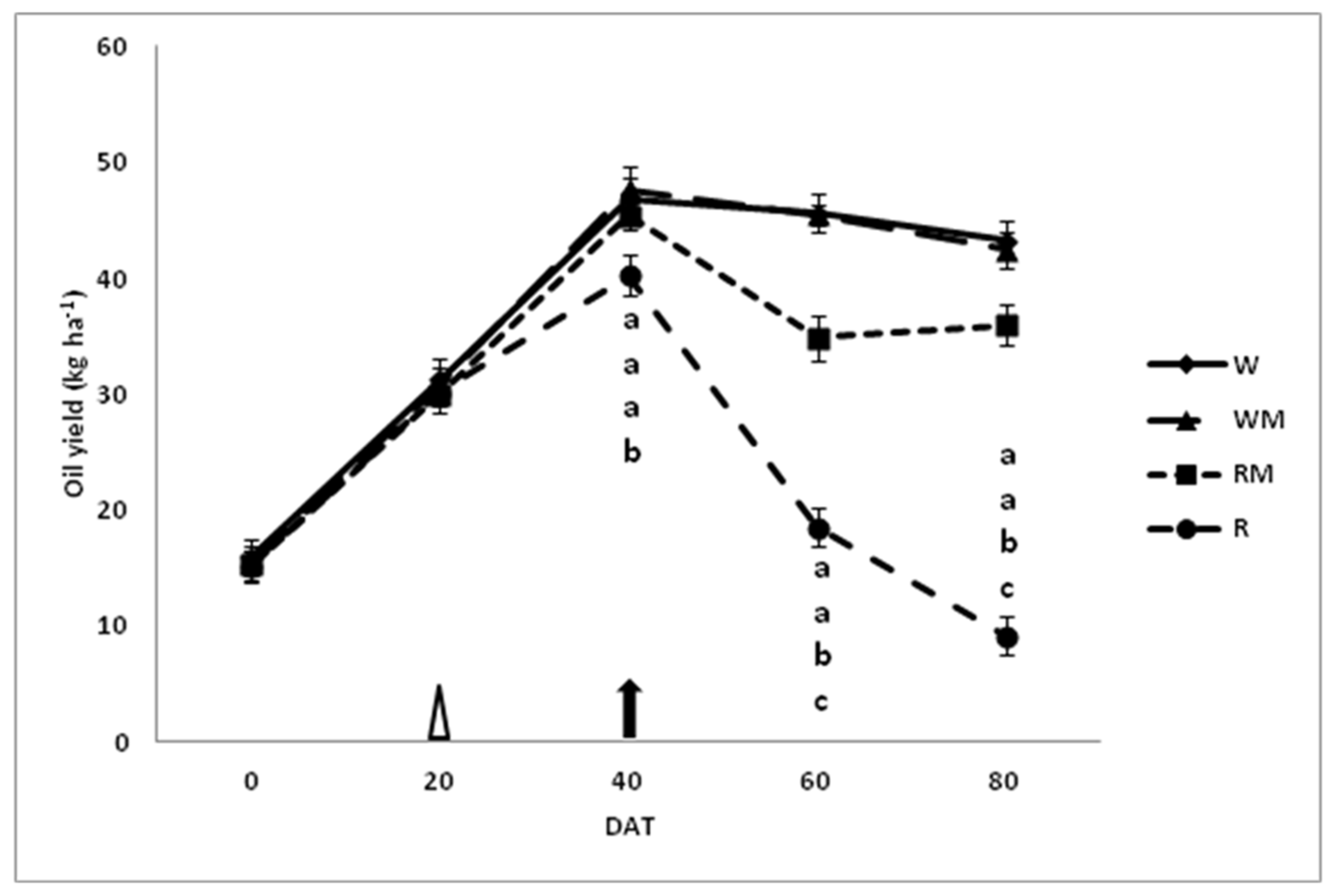
The mean essential-oil yields (
Figure 8) show significant decreases with increasing water deficit across the harvest time.
At 40 DAT, the essential-oil yield was significantly higher in W, WM and RM than in R, whereas the data did not differ significantly among W, WM and RM. The decrease in rainfed treatment at the end of harvest time (80 DAT) was much greater than rainfed mulching treatment compared to the respective control treatments (W and WM conditions). At the same time (80 DAT), the variation of essential-oil yields did not differ significantly between W and WM plants.
3.6. Major Oil Components
The major oil components that were compared were myrcene, limonene, 1,8-cineole and carvone, which comprise over 75% of the significant monoterpenes (
Table 1).
The percentage of myrcene did not differ significantly among treatments at 40 DAT. The percentages of limonene and 1,8-cineole, at 40 DAT, were significantly higher in R than in W, WM and RM, which did not differ significantly from each other, whereas at 40 DAT, the percentage of carvone was significantly lower in R than in W, WM and RM, which did not differ significantly from each other.
Even after 80 DAT, the percentage of myrcene did not differ significantly among the treatments. The increase in the percentage of limonene and 1,8-cineole in rainfed treatment, at the end of harvest time (80 DAT), was much greater than rainfed mulching treatment compared to the respective control treatments (W and WM conditions). At the same time (80 DAT), the variation of percentage of limonene and 1,8-cineole did not differ significantly between W and WM. At 80 DAT, the decrease in the percentage of carvone in rainfed treatment was much greater than rainfed mulching treatment compared to the respective control treatments (W and WM). The variation of the percentage of carvone did not differ significantly between W and WM. From 40 to 80 DAT, the result showed a significant increase in the myrcene, 1,8-cineole and carvone, and a significant decrease in the limonene percentage over the ranges of applied treatments (
Table 1).
4. Discussion
During the whole time of the experimentation, the scant precipitations occur in the form of low-intensity precipitation events. Thus, the precipitation quantities delivered in this manner cannot satisfy the normal growth of crops. Therefore, in these drought-prone environments, the soil-surface mulch practices may reduce the evaporation losses from the soil surface [
16,
21]. Thus, the improvement of the soil-moisture conditions of dry-land agriculture is of particular importance [
30,
31]. Our study indicated that rainfed mulching treatments increased the SWC within the 0–0.3 m soil profile compared with the rainfed conditions in the growing season of spearmint (
Figure 1). The SWC for the rainfed treatment was even lower when compared with the mulching treatment from the full bloom (40 DAT) to the end of the cropping cycle (80 DAT). Moreover, already after 20 DAT, the SWC within the 0–0.3 m soil profile under the rainfed treatment was generally lower than under well-watered treatment.
In our study, the mulch treatments of rainfed plants significantly increased the dry herbal biomass yields during the crop-growing stages (
Figure 2) in comparison to the rainfed treatment. Similar results have been reported on a menthol mint plants studied under different moisture regimes and nitrogen fertilization in the presence of sugarcane trash mulched [
10]. Instead, there were no significant differences in the dry herbal biomass yields between W and WM plants. It should be noted that as expected, well-irrigated plants (W and WM) had higher dry herbal biomass yields than rainfed ones (R and RM). The rainfed mulching treatment conserves soil water and reduces soil evaporation, thus demonstrating that it is suitable in a rainfed climate for the utilization of water [
32]. The dry herbal biomass accumulation was closely associated with photosynthetic gas-exchange parameters and leaf RWC during different crop-growth stages. There were no significant differences between W and WM plants during the entire spearmint growing season, which may be attributed to the similar tissue water content (
Figure 3,
Figure 4,
Figure 5 and
Figure 6).
The mulching rainfed treatment likely enhanced the accumulation of biomass by improving the photosynthesis to accelerate crop growth compared to the rainfed treatment. The photosynthesis limitations highlighted in R and RM plants compared to W and WM plants were mostly due to diffusive resistances, including the mesophyll component [
8]. Indeed, until 40 DAT, stomatal conductance in our study significantly decreased in R and RM plants in comparison to the well-watered controls (W and WM). To conclude that the reduction in photosynthesis observed until full bloom (40 DAT) is only attributable to stomatal closure, one should observe a consequent additional resistance to CO
2 diffusion toward the chloroplasts that is generally caused by the mesophyll components [
24]. This additional resistance may increase under stress conditions [
8]. However, mesophyll resistance is likely to be controlled by mesophyll anatomical structure and particularly by the chloroplast surfaces exposed to gas exchanges [
33,
34]. Mesophyll resistance was similar in controls (W and WM) and rainfed mulching treatment at 40 DAT (
Figure 5). Thus, it did not contribute to increasing resistance to CO
2 diffusion in RM leaves, indicating that mesophyll resistances to CO
2 diffusion did not directly affect photosynthesis [
8]. In this condition, in which it is only the stomatal conductance that limits photosynthesis, it did not cause permanent damage to the carbon fixation apparatus, as has been observed in other studies [
8,
24], and therefore, it is possible to consider the stress condition as moderate. Moreover, contrary to what was found at 40 DAT, we observed that at 60 and mostly at 80 DAT, the CO
2 mesophyll conductance of R leaves was significantly lower than that of the rainfed mulching treatment. Thus, mesophyll resistance only contributed to the total resistance to CO
2 diffusion from 60 DAT [
24]. During the harvesting period, when comparing R and RM plants it became clear that the total diffusive limitations were fewer in RM compared to R plants. Thus, the greater water availability increased the stomatal conductance, ensuring a greater input of carbon dioxide, which in turn increased photosynthetic activity in RM compared to R plants. These differences significantly accelerated the growth of the crop canopy and resulted in the significantly increased dry herbal biomass values that are shown in
Figure 1 [
35].
In our study, the rainfed treatments (without and with mulch) significantly increased the essential oil content, during the full bloom (40 DAT) (
Figure 7). The rainfed treatment significantly increased the essential-oil content and storage in green tissues in comparison to the W, WM and RM treatments, before and during the early growth stage (20 DAT) [
8]. During the withering stages, there were no significant differences in the essential-oil content between the well-watered control plants (W and WM) and RM plants, while the essential-oil content of R plants strongly decreased. Our study indicated that mulching rainfed treatment (RM) significantly increased the essential-oil content compared to the control treatment (WM) in the full-bloom phase and kept it constant until the end of the growing season during withering, extending the oil-extraction period (
Figure 7).
Essential oil yield increased substantially with the increasing of bloom for all treatments (
Figure 8). At 40 DAT, compared with rainfed treatment, mulching rainfed spearmint (RM) clearly yielded the same amount of essential oil as the W and WM controls. Moreover, after 80 DAT, the reduction in essential-oil yield in the mulching rainfed treatment (RM, −15%), although significant, was much less evident than in the rainfed treatment (R, −78%) compared to the respective controls (WM and W).
This indicates a possibility, at the full-bloom stage (40 DAT), for decreased grower costs through mulching rainfed treatment by cutting costs for water, harvesting, transportation and distilling spearmint hay; and for increasing profits if the oil yield and quality remain fairly constant. This encouraging result for mulching rainfed treatment (RM) is attributable to the presence of higher soil water contents compared to rainfed treatment (R), which helped sustain the crop at these rainfed conditions in mulching treatment.
The percentage of four major monoterpenes (limonene, carvone, 1,8-cineole and myrcene) showed a low sensitivity to rainfed conditions if at moderate stress levels (
Table 1), or 40 DAT and in the presence of mulch (RM). Over time, from 40 to 80 DAT, the percentage of carvone significantly increased, whereas that of limonene decreased over the ranges of applied treatments (
Table 1). Instead, at both 40 and 80 DAT, the result shows a significant decrease in the carvone and a significant increase in limonene percentage with the increase in water-stress conditions due to reduced water availability (R). This indicates that rainfed plants may show different metabolic maturity as limonene is a precursor to carvone [
3,
6].












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}