
Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Material
2.2. Sampling and Measurements
2.2.1. Biomass Yield and Determination of Chemical Composition
2.2.2. Color Assessment
2.2.3. Determination of Antinutritional Composition
Phenolic Extracts
Total Phenolics
Hydrolyzed Tannins
Condensed Tannins
Oxalate Content
2.3. Statistical Analyses
3. Results
3.1. Weather Conditions
3.2. Influence of Genotype, Environment, and Their Interaction
3.3. Variation between Genotypes
3.4. Variation between Environments
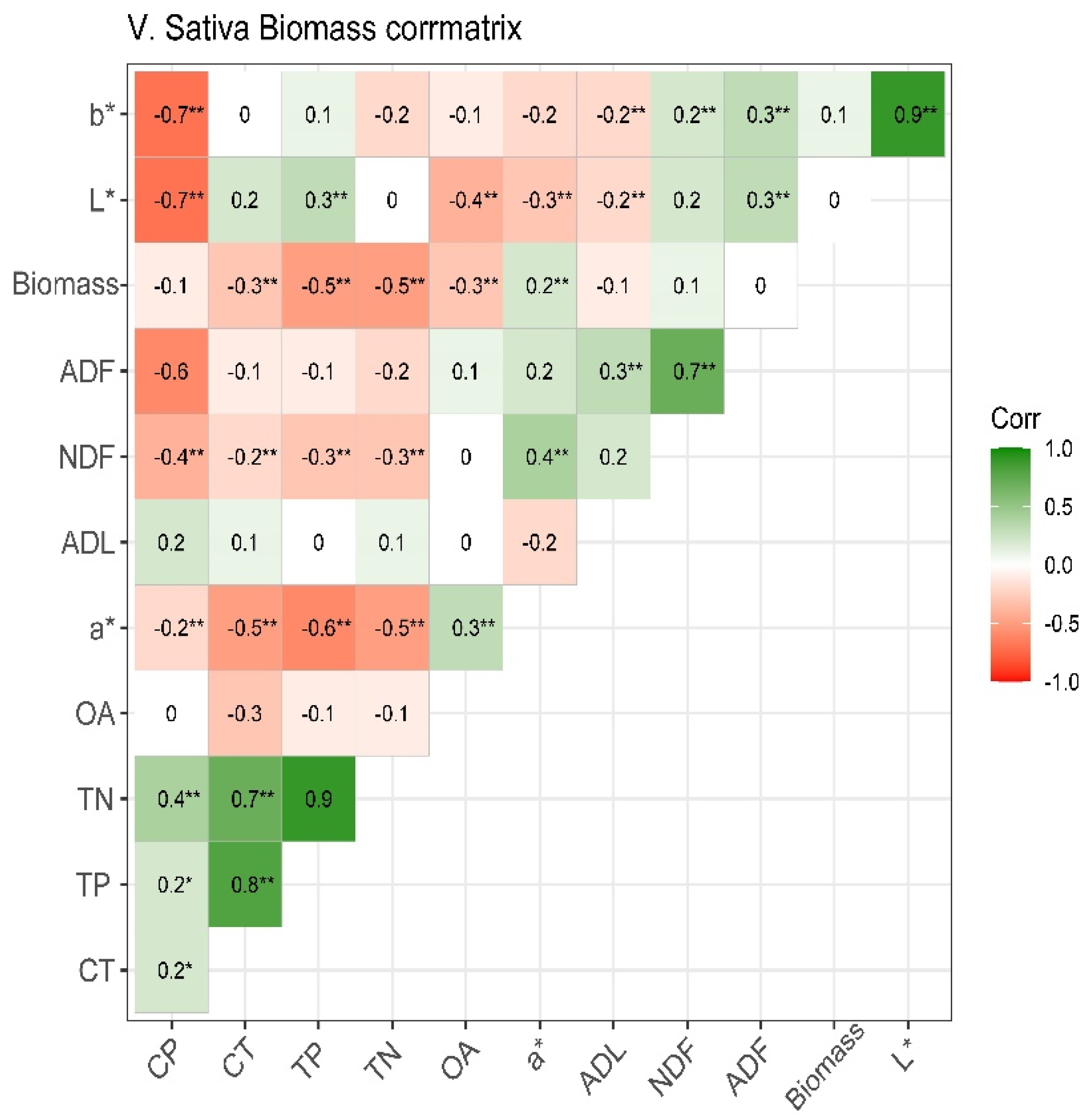
3.5. Correlation Coefficients between Traits
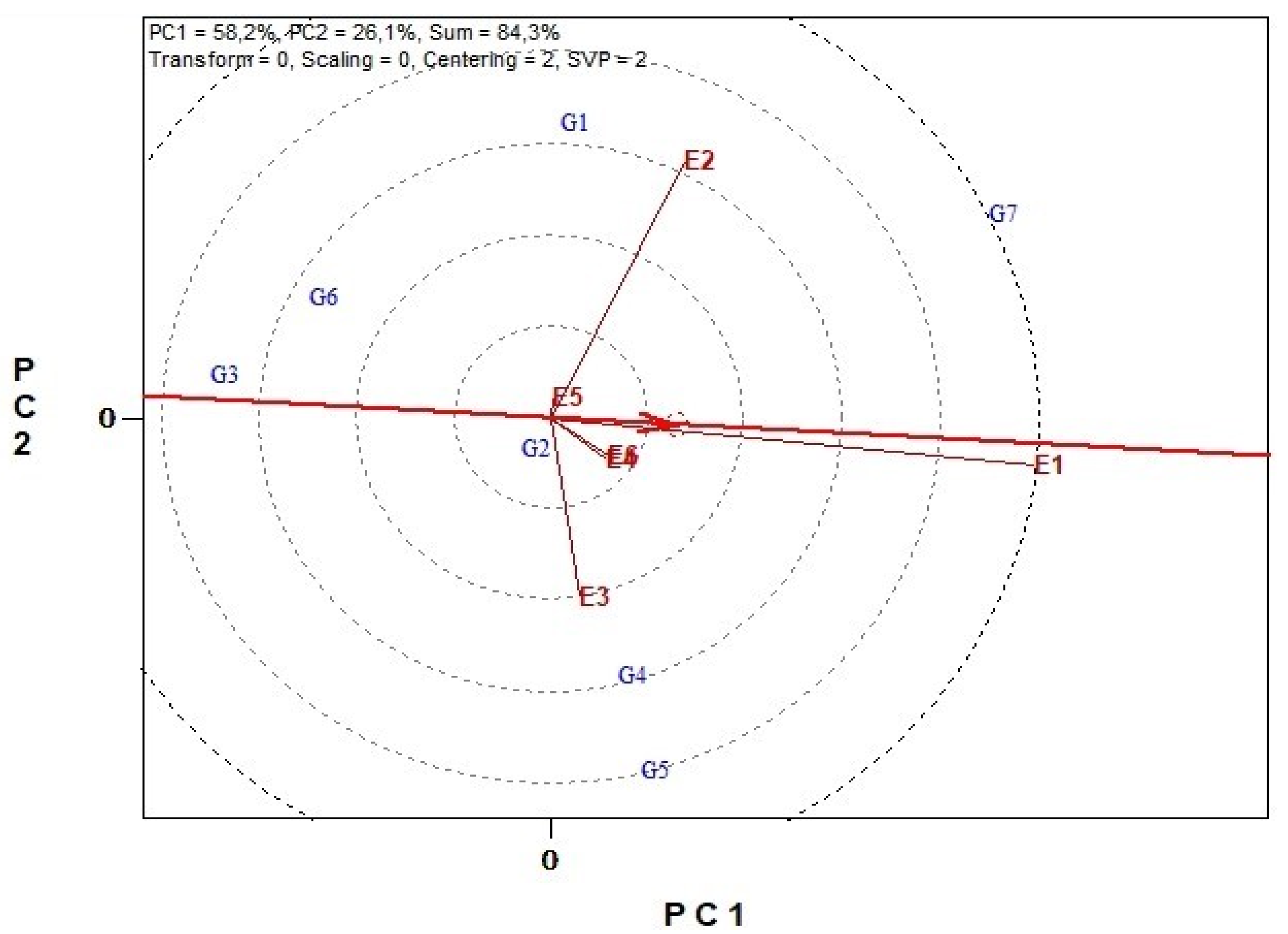
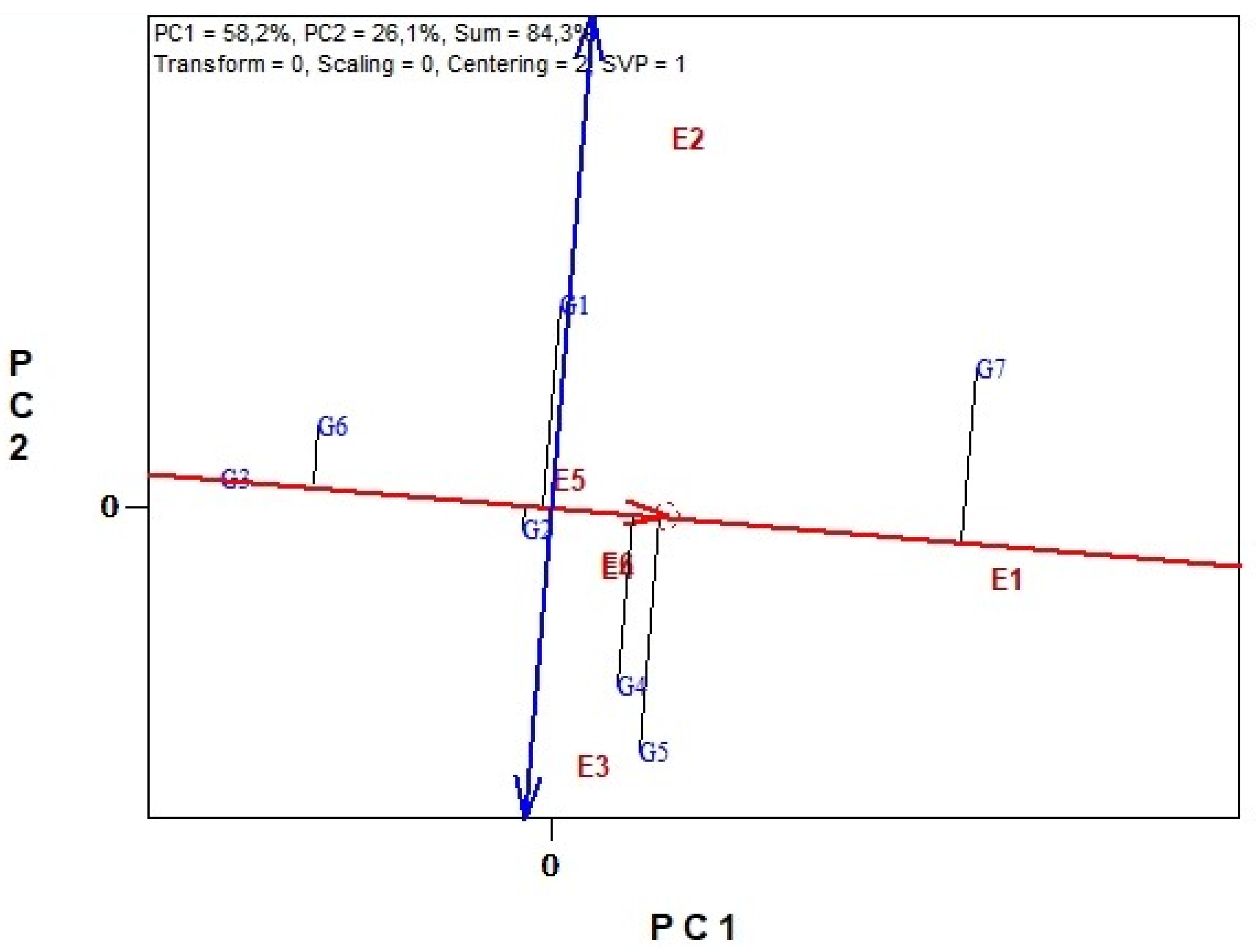
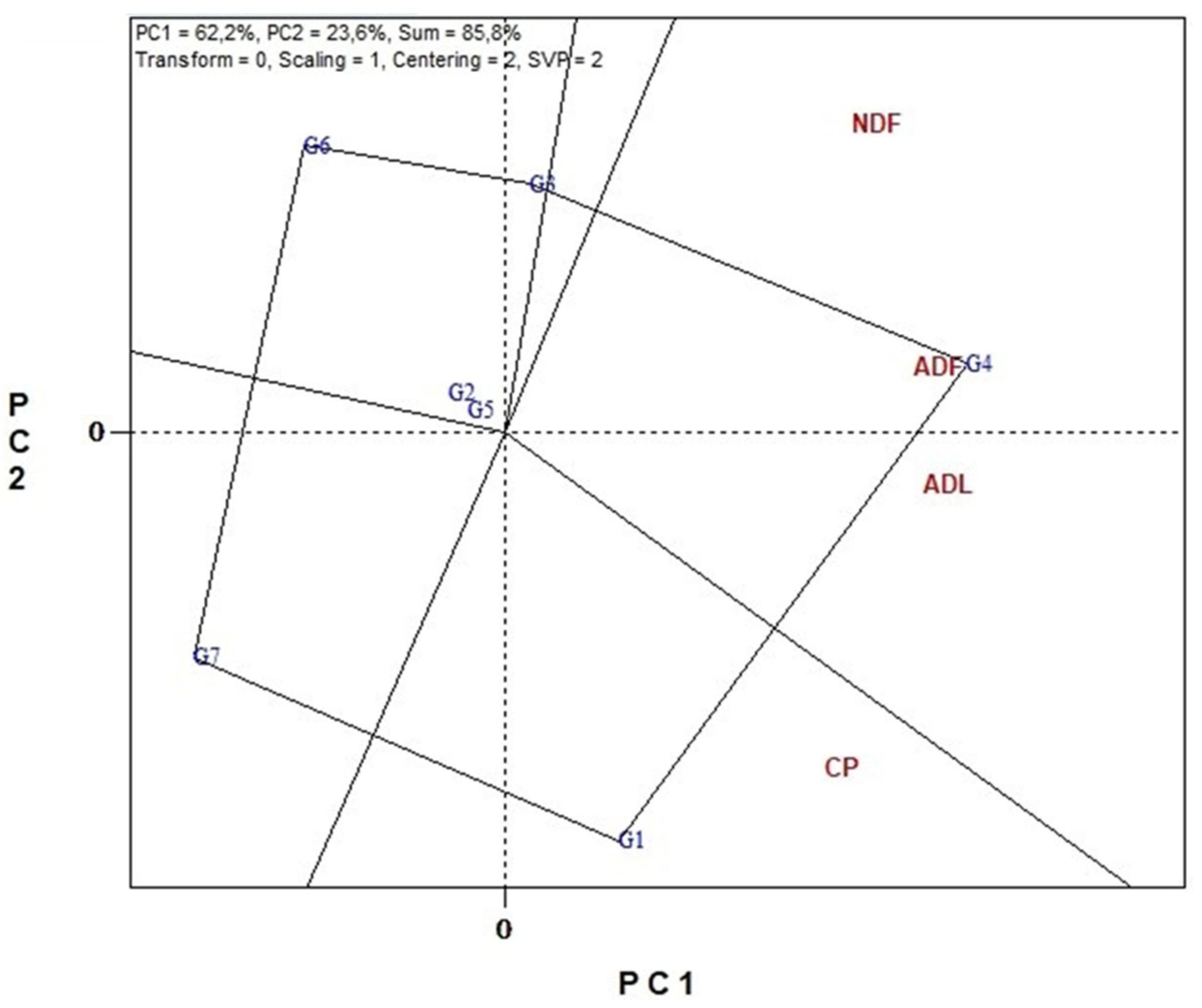
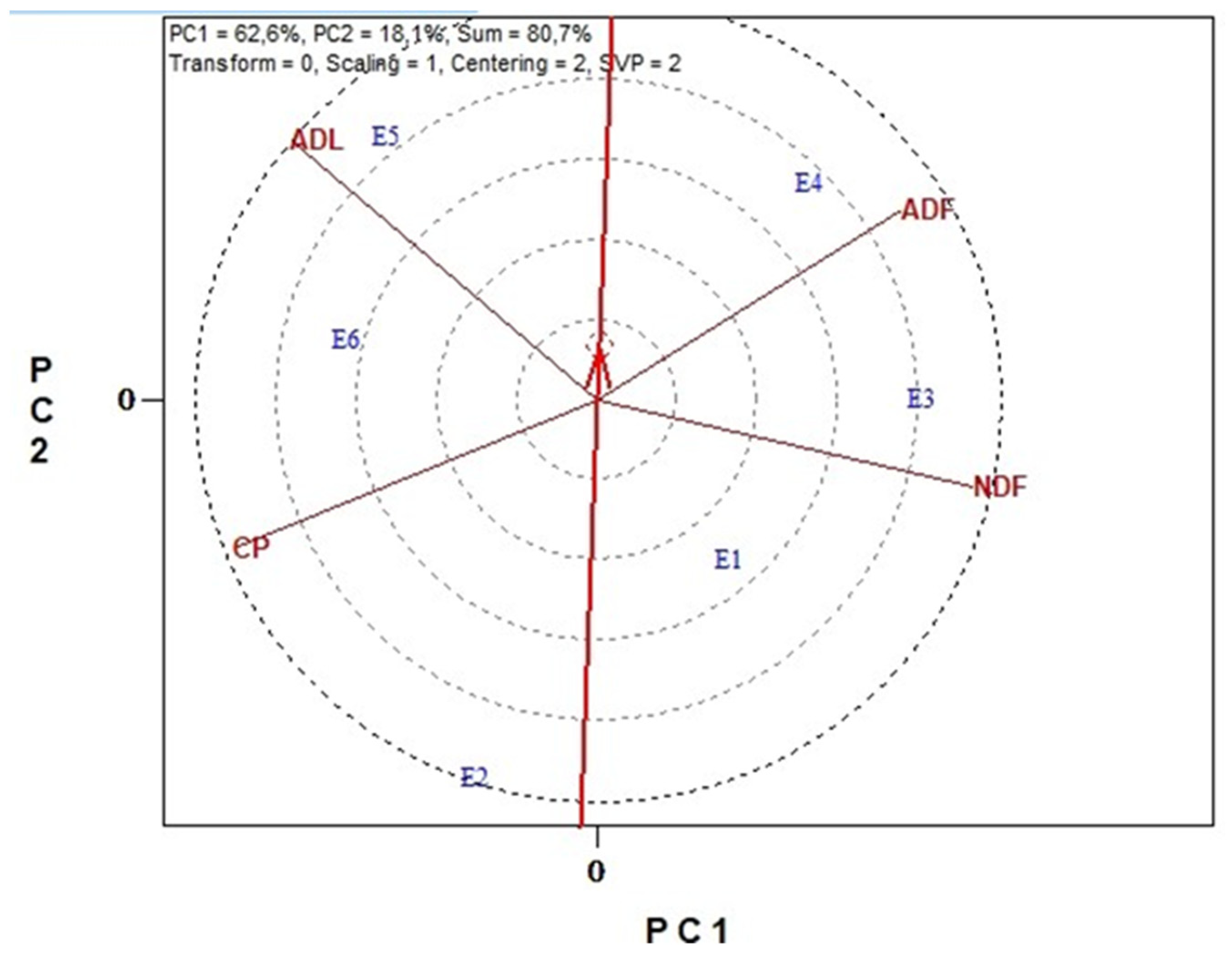
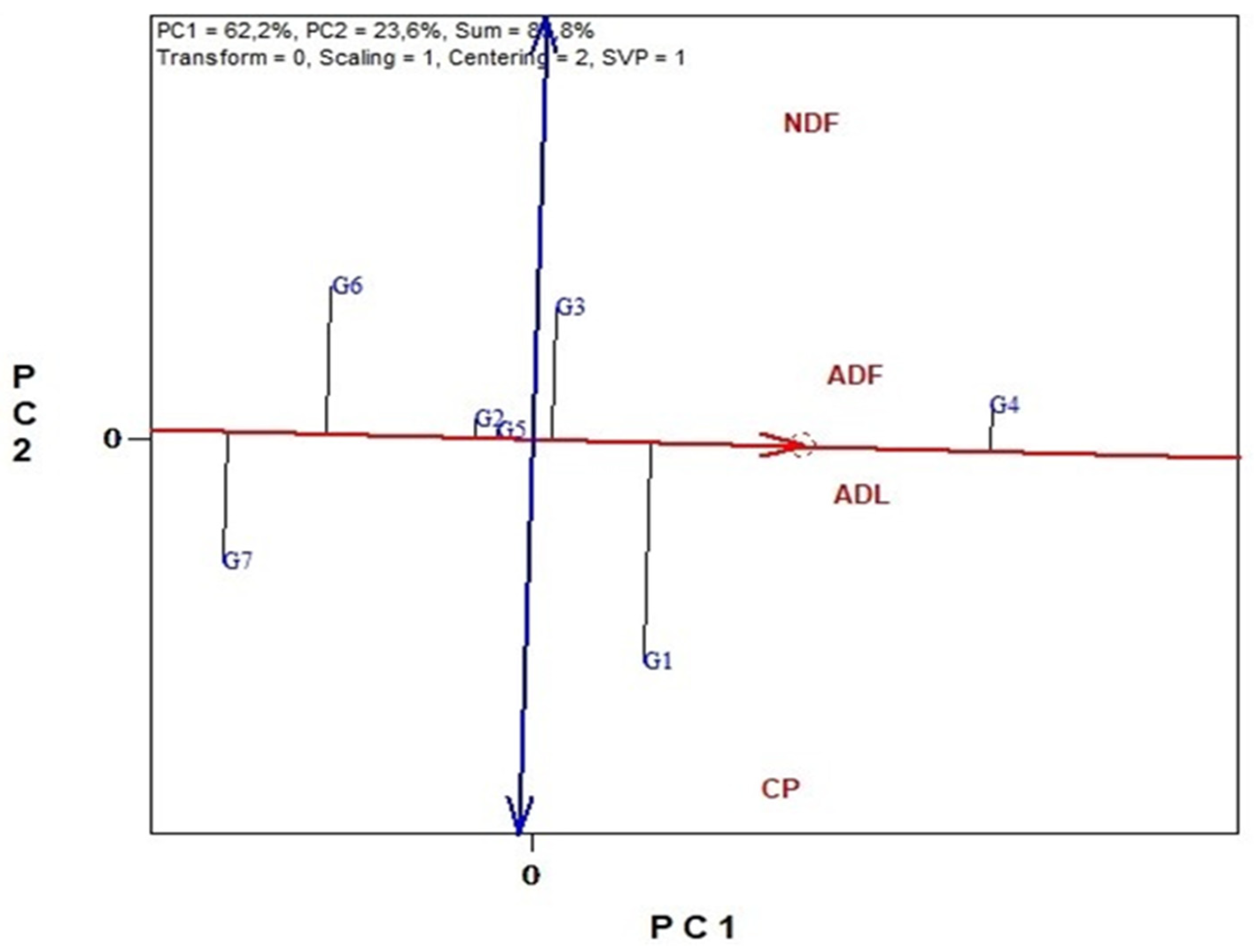
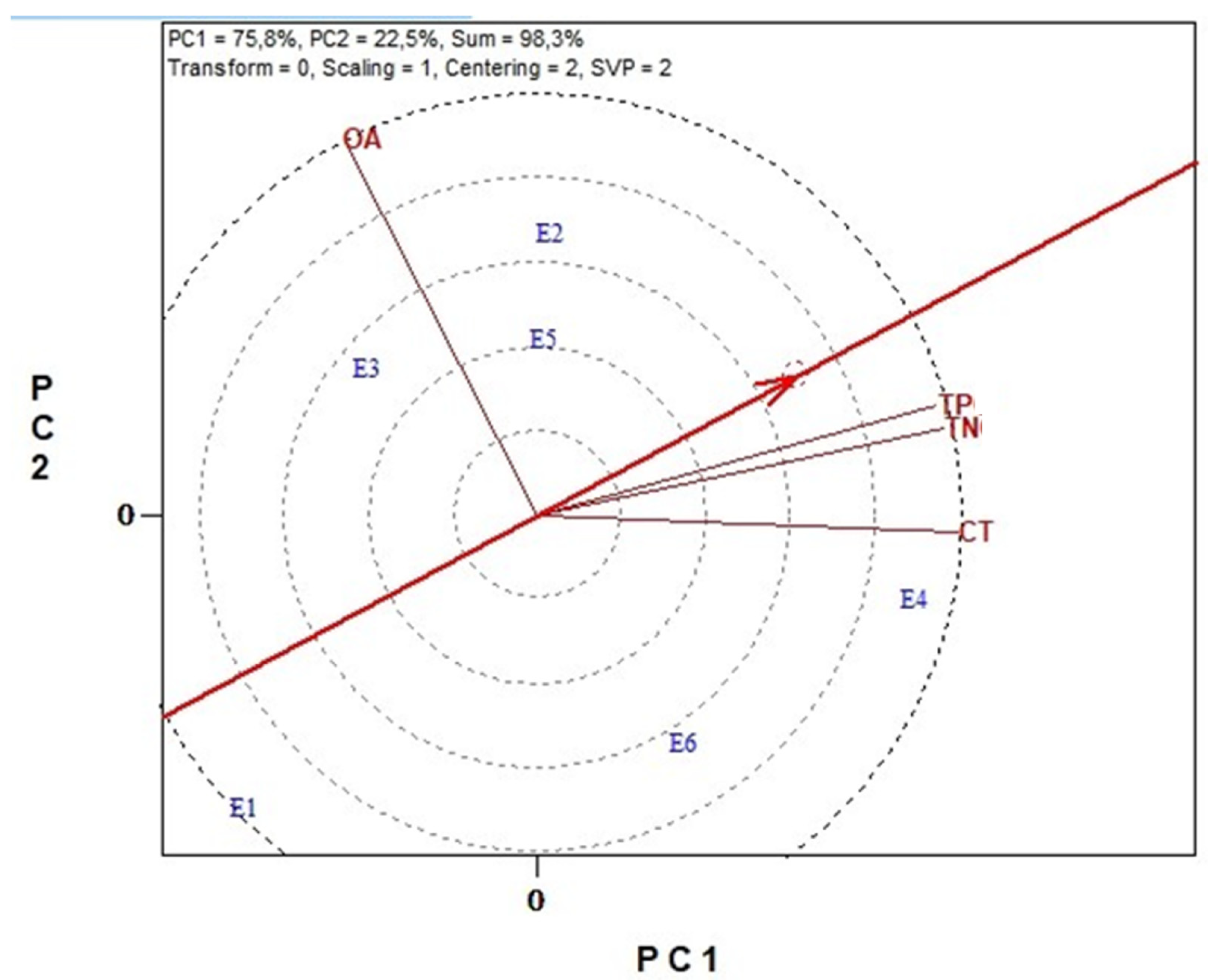
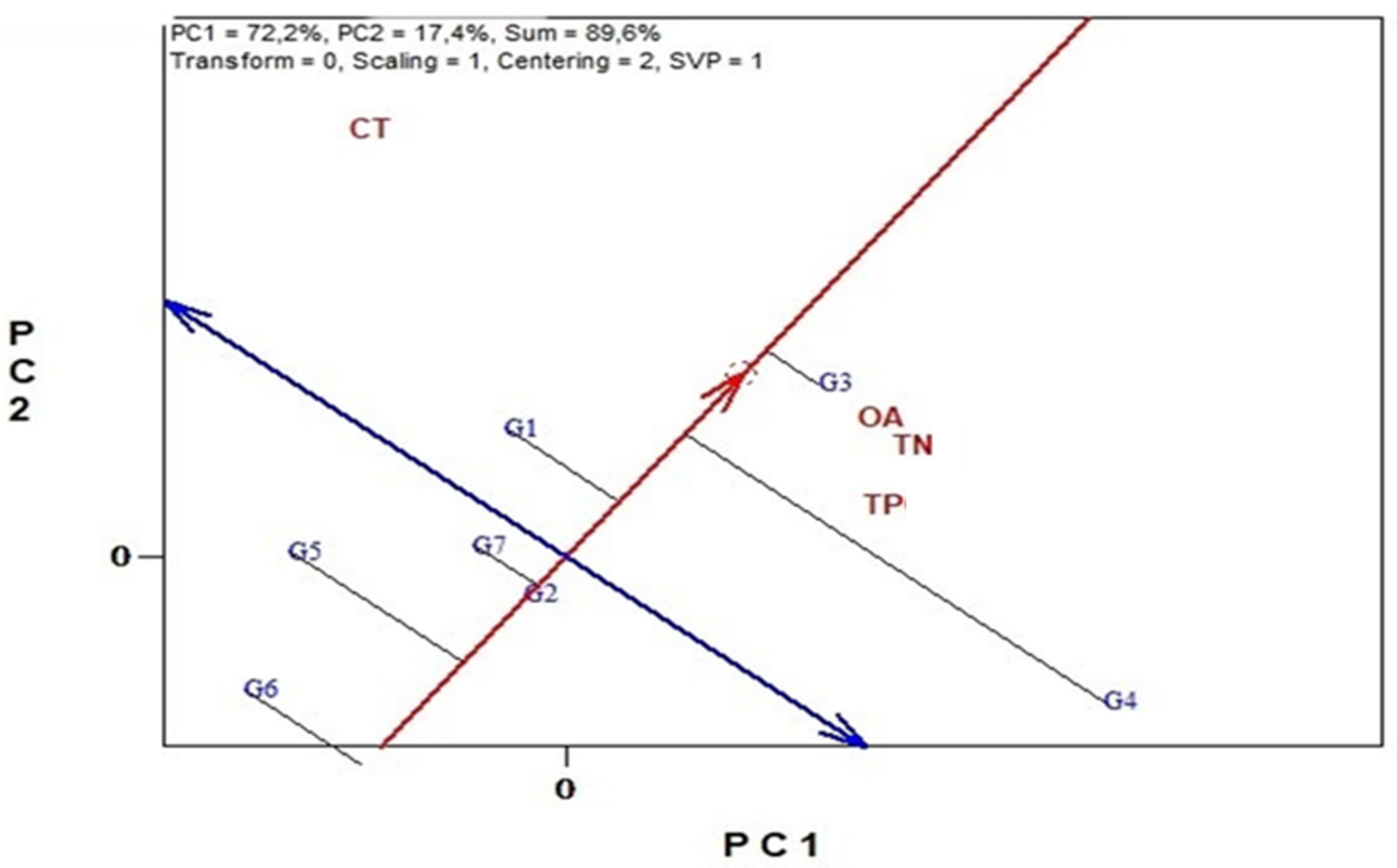
3.6. Genotype Stability and Environment Discrimination with GGE Biplots Analysis
3.6.1. Biomass Yield
3.6.2. Nutritional Traits
3.6.3. Antinutritional Traits
4. Discussion
4.1. Biomass/Colorimetric Traits
4.2. Nutritional Traits
4.3. Antinutritional Traits
Correlation between Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, R.; Jahufer, M.Z.Z.; Dong, D.K.; Wang, Y.R.; Liu, Z.P. Characterisation of the morphological variation for seed traits among 537 germplasm accessions of common vetch (Vicia sativa L.) using digital image analysis. N. Z. J. Agric. Res. 2016, 59, 422–435. [Google Scholar] [CrossRef]
- Abbeddou, S.; Rihawi, S.; Hess, H.D.; Iñiguez, L.; Mayer, A.C.; Kreuzer, M. Nutritional composition of lentil straw, vetch hay, olive leaves, and saltbush leaves and their digestibility as measured in fat-tailed sheep. Small Rumin. Res. 2011, 96, 126–135. [Google Scholar] [CrossRef]
- Larbi, A.; El-Moneim, A.M.; Nakkoul, H.; Jammal, B.; Hassan, S. Intra-species variations in yield and quality determinants in Vicia species: 3. Common vetch (Vicia sativa ssp. sativa L.). Anim. Feed Sci. Technol. 2011, 164, 241–251. [Google Scholar] [CrossRef]
- Huang, Y.F.; Gao, X.L.; Nan, Z.B.; Zhang, Z.X. Potential value of the common vetch (Vicia sativa L.) as an animal feedstuff: A review. J. Anim. Physiol. Anim. Nutr. 2017, 101, 807–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lithourgidis, A.S.; Dordas, C.A.; Damalas, C.A.; Vlachostergios, D.N. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Vlachostergios, D.N.; Lithourgidis, A.S.; Dordas, C.A.; Baxevanos, D. Advantages of mixing common vetch cultivars developed from conventional breeding programs when grown under low-input farming system. Crop Sci. 2011, 51, 1274–1281. [Google Scholar] [CrossRef]
- Vlachostergios, D.N.; Lithourgidis, A.S.; Korkovelos, A.; Baxevanos, D.; Lazaridou, T.; Khah, A.; Mavromatis, A. Mixing ability of conventionally bred common vetch (Vicia sativa L.) cultivars for grain yield under low-input cultivation. Aust. J. Crop Sci. 2011, 5, 1588–1594. [Google Scholar]
- Tigka, E.; Beslemes, D.; Kakabouki, I.; Pankou, C.; Bilalis, D.; Tokatlidis, I.; Vlachostergios, D.N. Seed Rate and Cultivar Effect on Contribution of Vicia sativa L. Green Manure to Soil Amendment under Mediterranean Conditions. Agriculture 2021, 11, 733. [Google Scholar] [CrossRef]
- Cakmakci, S.; Aydinoglu, B.; Karaca, M.; Bilgen, M. Heritability of yield components in common vetch (Vicia sativa L.). Acta Agric. Scand. B Soil Plant Sci. 2006, 56, 54–59. [Google Scholar]
- Ford, R.; Maddeppungeng, A.M.; Taylor, P.W.J. Vetch. In Compendium of Transgenic Crop Plants: Transgenic Legume Seeds and Forages; Kole, C., Hall, T.C., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 163–170. [Google Scholar]
- De la Rosa, L.; López-Román, M.I.; González, J.M.; Zambrana, E.; Marcos-Prado, T.; Ramírez-Parra, E. Common Vetch, Valuable Germplasm for Resilient Agriculture: Genetic Characterization and Spanish Core Collection Development. Front. Plant Sci. 2021, 12, 617873. [Google Scholar] [CrossRef]
- Nguyen, V.; Riley, S.; Nagel, S.; Fisk, I.; Searle, I.R. Common Vetch: A Drought Tolerant, High Protein Neglected Leguminous Crop with Potential as a Sustainable Food Source. Front. Plant Sci. 2020, 11, 818. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, T.; Wang, J.; Wang, Y.; Baskin, C.C.; Baskin, J.M. Seed dormancy in four Tibetan Plateau Vicia species and characterization of physiological changes in response of seeds to environmental factors. Seed Sci. Res. 2013, 23, 133–140. [Google Scholar] [CrossRef]
- Georgieva, N.; Nikolova, I.; Delchev, G. Response of spring vetch (Vicia sativa L.) to organic production conditions. Bulg. J. Agric. Sci. 2020, 26, 520–526. [Google Scholar]
- ELSTAT. Hellenic Statistical Authority Statistics—Agriculture, Livestock and Fishery. 2021. Available online: http://www.statistics.gr/en/statistics (accessed on 30 June 2022).
- Greveniotis, V.; Bouloumpasi, E.; Zotis, S.; Korkovelos, A.; Ipsilandis, C.G. Assessment of Interactions between Yield Components of Common Vetch Cultivars in Both Conventional and Low-Input Cultivation Systems. Agriculture 2021, 11, 369. [Google Scholar] [CrossRef]
- Beyene, Y.; Mugo, S.; Mutinda, C.; Tefera, T.; Karaya, H.; Ajanga, J.; Shuma, J.; Tende, R.; Kega, V. Genotype by environment interactions and yield stability of stem borer resistant maize hybrids in Kenya. Afr. J. Biotechnol. 2011, 10, 4752–4758. [Google Scholar]
- Berger, J.D.; Robertson, L.D.; Cocks, P.S. Genotype x environment interaction for yield and other plant attributes among undomesticated Mediterranean Vicia species. Euphytica 2002, 126, 421–435. [Google Scholar] [CrossRef]
- Yücel, C.; Hizli, H.; Firincioğlu, H.K.; Çil, A.; Anlarsal, A.E. Forage yield stability of common vetch (Vicia sativa L.) genotypes in the Çukurova and GAP regions of Turkey. Turk. J. Agric. 2009, 33, 119–125. [Google Scholar] [CrossRef]
- Aydemir, S.K.; Karakoy, T.; Kokten, K.; Nadeem, M.A. Evaluation of yield and yield components of common vetch (Vicia sativa L.) genotypes grown in different locations of Turkey by GGE biplot analysis. Appl. Ecol. Environ. Res. 2019, 17, 15203–15217. [Google Scholar] [CrossRef]
- Dong, R.; Shen, S.S.; Jahufer, M.Z.Z.; Dong, D.K.; Luo, D.; Zhou, Q.; Chai, X.T.; Luo, K.; Nan, Z.B.; Wang, Y.R.; et al. Effect of genotype and environment on agronomical characters of common vetch (Vicia sativa L.). Genet. Resour. Crop Evol. 2019, 66, 1587–1599. [Google Scholar] [CrossRef]
- Moore, K.J.; Curtiss, C.F.; Lenssen, A.W.; Fales, S.L. Factors Affecting Forage Quality. In Forages Volume II: The Science of Grassland Agriculture; Moore, K.J., Collins, M., Nelson, C.J., Redfearn, D.D., Eds.; Wiley: West Sussex, UK, 2020; pp. 701–717. [Google Scholar]
- Georgieva, N.; Nikolova, I.; Naydenova, Y. Nutritive value of forage of Vetch cultivars (Vicia sativa L., Vicia villosa ROTH). Banat. J. Biotechnol. 2016, VII, 5–12. [Google Scholar] [CrossRef]
- Bakoğlu, A.; Kökten, K.; Kiliç, Ö. Yield and Nutritive Value of Common Vetch (Vicia sativa L.) Lines and Varieties. Turk. J. Agric. Nat. Sci. 2016, 3, 33–37. [Google Scholar]
- Quesenberry, K.H.; Casler, M.D. Achievements and Perspectives in the Breeding of Temperate Grasses and Legumes. In Proceedings of the XIX International Grassland Congress, São Pedro, SP, Brazil, 11–21 February 2001. [Google Scholar]
- Kölliker, R.; Kempf, K.; Malisch, C.S.; Lüscher, A. Promising options for improving performance and proanthocyanidins of the forage legume sainfoin (Onobrychis viciifolia Scop.). Euphytica 2017, 213, 179. [Google Scholar] [CrossRef] [Green Version]
- Atumo, T.T.; Kauffman, R.; Talore, D.G.; Abera, M.; Tesfaye, T.; Tunkala, B.Z.; Zeleke, M.; Kalsa, G.K. Adaptability, forage yield and nutritional quality of alfalfa (Medicago sativa) genotypes. Sustain. Environ. 2021, 7, 1895475. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, Z.; Nan, Z.; Unkovich, M.; Coulter, J.A. Effects of cultivar and growing degree day accumulations on forage partitioning and nutritive value of common vetch (Vicia sativa L.) on the Tibetan plateau. J. Sci. Food Agric. 2020, 101, 3749–3757. [Google Scholar] [CrossRef] [PubMed]
- Papastylianou, P.; Vlachostergios, D.N.; Dordas, C.; Tigka, E.; Papakaloudis, P.; Kargiotidou, A.; Pratsinakis, E.; Koskosidis, A.; Pankou, C.; Kousta, A.; et al. Genotype x environment interaction analysis of faba bean (Vicia faba L.) for biomass and seed yield across different environments. Sustainability 2021, 13, 2586. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnson, C.T.; Sumner, M.E. Methods of Soil Analysis: Part 3-Chemical Methods, 2nd ed.; American Society of Agronomy and Soil Science Society of Agronomy: Madison, WI, USA, 1996; p. 1390. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2002. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventοs, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteu reagent. Methods Enzymol. 1998, 299, 152–178. [Google Scholar]
- Makkar, H.P.S.; Bluemmel, M.; Borowy, N.K.; Becker, R.K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Oyeyinka, B.O.; Afolayan, A.J. Comparative evaluation of the nutritive, mineral, and antinutritive composition of Musa sinensis L. (Banana) and Musa paradisiaca L. (Plantain) fruit compartments. Plants 2019, 720, 598. [Google Scholar] [CrossRef] [Green Version]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Yan, W. GGE biplot—A Windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Yan, W. Singular-value partitioning in biplot analysis of multi-environment trial data. Agron. J. 2002, 94, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.W. GenStat Wiley Interdiscip. Rev. Comput. Stat. 2009, 1, 255–258. [Google Scholar]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Yan, W.; Tinker, N.A. An integrated system of biplot analysis for displaying, interpreting, and exploring genotype x environment interactions. Crop Sci. 2005, 45, 1004–1016. [Google Scholar] [CrossRef]
- Kissing Kucek, L.; Riday, H.; Rufener, B.P.; Burke, A.N.; Eagen, S.S.; Ehlke, N.; Krogman, S.; Mirsky, S.B.; Reberg-Horton, C.; Ryan, M.R.; et al. Pod dehiscence in hairy vetch (Vicia villosa Roth). Front. Plant Sci. 2020, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, C.A.; Reckling, M.; Preissel, S.; Bachinger, J.; Bergkvist, G.; Kuhlman, T.; Lindström, K.; Nemecek, T.; Topp, C.F.E.; Vanhatalo, A.; et al. Grain Legume Production and Use in European Agricultural Systems. Adv. Agron. 2017, 144, 235–303. [Google Scholar]
- Irakli, M.; Kargiotidou, A.; Tigka, E.; Beslemes, D.; Fournomiti, M.; Pankou, C.; Stavroula, K.; Tsivelika, N.; Vlachostergios, D.N. Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.). Agronomy 2021, 11, 1154. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agric. Water Manag. 2017, 179, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Cevher, C.; Altunkaynak, B. Socioeconomic Factors and Sustainable Forage Crops Production in Turkey Aegean Region: A Multivariate Modeling. Sustainability 2020, 12, 8061. [Google Scholar] [CrossRef]
- Seymour, M.; Siddique, K.H.M.; Brandon, N.; Martin, L.; Jackson, E. Response of vetch (Vicia spp.) to plant density in south-western Australia. Aust. J. Exp. Agric. 2002, 42, 1043–1051. [Google Scholar] [CrossRef]
- Archetti, M.; Richardson, A.D.; O’Keefe, J.; Delpierre, N. Predicting climate change impacts on the amount and duration of autumn colors in a New England Forest. PLoS ONE 2013, 8, e57373. [Google Scholar] [CrossRef] [PubMed]
- Alamu, E.O.; Maziya-Dixon, B.; Menkir, A.; Ogunlade, A.O.; Olaofe, O. Harvesting time and roasting effects on colour properties, xanthophylls, phytates, tannins and vitamin C contents of orange maize hybrid. Sci. Rep. 2020, 10, 21327. [Google Scholar] [CrossRef] [PubMed]
- Ammar, H.; López, S.; González, J.S.; Ranilla, M.J. Relationship between chemical composition and in vitro digestibility of some Spanish browse plants species. In Sustainable Grazing, Nutritional Utilization and Quality of Sheep and Goat Products; Molina, A.E., Ben Salem, H., Biala, K., Morand-Fehr, P., Eds.; Options Méditerranéennes: Série, A. Séminaires Méditerranéens; n. 67; CIHEAM: Zaragoza, Spain, 2005; pp. 327–332. [Google Scholar]
- Dewhurst, R.J.; Davies, D.W.R.; Fisher, W.J. Effects of forage NDF content and body condition score on forage intake by Holstein–Friesian dairy cows in the dry period. Animals 2010, 4, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Asekova, S.; Kulkarni, K.P.; Kim, M.; Kim, J.H.; Song, J.T.; Shannon, J.G.; Lee, J.D. Novel quantitative trait loci for forage quality traits in a cross between PI 483463 and ‘Hutcheson’ in soybean. Crop Sci. 2016, 56, 2600–2611. [Google Scholar] [CrossRef]
- Pecetti, L.; Annicchiarico, P.; Scotti, C.; Paolini, M.; Nanni, V.; Palmonari, A. Effects of plant architecture and drought stress level on lucerne forage quality. Grass Forage Sci. 2016, 72, 714–722. [Google Scholar] [CrossRef]
- Rebolé, A.; Alzueta, C.; Ortiz, L.T.; Barro, C.; Rodríguez, M.L.; Caballero, R. Yields and chemical composition of different parts of the common vetch at flowering and at two seed filling stages. Span. J. Agric. Res. 2004, 2, 550–557. [Google Scholar] [CrossRef]
- Huang, Y.; Matthew, C.; Li, F.; Nan, Z. Comparative effects of stovers of four varieties of common vetch on growth performance, ruminal fermentation, and nutrient digestibility of growing lambs. Animals 2020, 10, 596. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.F.; Matthew, C.; Li, F.; Nan, Z.B. Common vetch varietal differences in hay nutritive value, ruminal fermentation, nutrient digestibility and performance of fattening lambs. Animal 2021, 15, 100244. [Google Scholar] [CrossRef]
- Kaya, E. Effect of species on nutritive value and anti-methanogenic potential of vetch hays grown in native pasture in Turkey. Prog. Nutr. 2021, 23, e2021049. [Google Scholar]
- Morand-Fehr, P. Nutrition and Feeding of Goats: Application to Temperate Climatic Conditions. In Goat Production; Gall, C., Ed.; Academic Press: London, UK, 1981; pp. 193–232. [Google Scholar]
- NRC–National Research Council. Nutrient Requirements of Goats: Angora, Dairy and Meat Goats in Temperate and Tropical Countries; National Academies Press: Washington, DC, USA, 1981. [Google Scholar]
- NRC–National Research Council. Nutrient Requirements of Sheep, 6th ed.; National Academies Press: Washington, DC, USA, 1985. [Google Scholar]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. Role of phenols and polyphenols in plant defense response to biotic and abiotic stresses. In Biocontrol Agents and Secondary Metabolites; Jogaiah, S., Ed.; Elsevier Inc.: Duxford, UK, 2021; pp. 419–441. [Google Scholar]
- Naumann, H.D.; Tedeschi, I.O.; Zeller, W.E.; Huntley, N.F. The role of condensed tannins in ruminant animal production: Advances, limitations and future directions. Rev. Bras. Zootec. 2017, 46, 929–949. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Abdullah, R.B.; Wan Khadijah, W.E. A review of oxalate poisoning in domestic animals: Tolerance and performance aspects. J. Anim. Physiol. Anim. Nutr. 2013, 97, 605–614. [Google Scholar] [CrossRef] [PubMed]
- El Shaer, H.M. Halophytes and salt-tolerant plants as potential forage for ruminants in the Near East region. Small Rumin. Res. 2010, 91, 3–12. [Google Scholar] [CrossRef]
- Seguin, P.; Mustafa, A.F.; Donnelly, D.; Gélinas, B. Chemical composition and ruminal nutrient degradability of fresh and ensiled amaranth forage. J. Sci. Food Agric. 2013, 93, 3730–3736. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Ansari, R.; Ali, H.; Gul, B.; Nielsen, B.L. Panicum turgidum, a potentially sustainable cattle feed alternative to maize for saline areas. Agric. Ecosyst. Environ. 2009, 129, 542–546. [Google Scholar] [CrossRef]
- Goyal, M. Oxalate accumulation in fodder crops and impact on grazing animals—A review. Forage Res. 2018, 44, 152–158. [Google Scholar]
- Rahman, M.M.; Nakagawa, T.; Niimi, M.; Fukuyama, K.; Kawamura, O. Effects of feeding oxalate containing grass on intake and the concentrations of some minerals and parathyroid hormone in blood of sheep. Asian Australas. J. Anim. Sci. 2011, 24, 940–945. [Google Scholar] [CrossRef]
- Ncube, B.; Finnie, J.F.; Van Staden, J. Quality from the field: The impact of environmental factors as quality determinants in medicinal plants. S. Afr. J. Bot. 2012, 82, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Iwaasa, A.D.; Wang, Y.; Jin, L.; Han, G.; Zhao, M. Condensed tannins concentration of selected prairie legume forages as affected by phenological stages during two consecutive growth seasons in western Canada. Can. J. Plant Sci. 2014, 94, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Ushio, M.; Adams, J.M. A meta-analysis of the global distribution pattern of condensed tannins in tree leaves. Open Ecol. J. 2011, 4, 18–23. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kawamura, O. Oxalate accumulation in forage plants: Some agronomic, climatic and genetic aspects. Asian Aust. J. Anim. Sci. 2011, 24, 439–448. [Google Scholar] [CrossRef]
- Bernardo, R. What if we knew all the genes for a quantitative trait in hybrid crops? Crop Sci. 2001, 41, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Canbolat, O.; Tamer, E.; Acikgoz, E. Chemical composition, metabolizable energy and digestibility in pea seeds of differing testa and flower colors. J. Biol. Environ. Sci. 2007, 1, 59–65. [Google Scholar]
- Wang, X.; Warkentin, T.D.; Briggs, C.J.; Oomah, B.D.; Campbell, C.G.; Woods, S. Total phenolics and condensed tannins in field pea (Pisum sativum L.) and grass pea (Lathyrus sativus L.). Euphytica 1998, 101, 97–102. [Google Scholar] [CrossRef]
- Menga, V.; Codianni, P.; Fares, C. Agronomic management under organic farming may affect the bioactive compounds of lentil (Lens culinaris L.) and grass pea (Lathyrus communis L.)? Sustainability 2014, 6, 1059–1075. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment | Sand% | Clay% | Loam% | pH | CaCO3 (%) | EC (μS/cm) | Organic Matter (%) |
|---|---|---|---|---|---|---|---|
| AUA, Southern Greece Env1 + Εnv2 | 50 | 26 | 24 | 7.9 | 11.6 | 3.0 | 1.43 |
| IIFC, Central Greece Env3 + Env4 | 20 | 57 | 23 | 8.0 | 1.5 | 4.9 | 1.30 |
| AUTh, Northern Greece Env5 + Env6 | 25 | 27 | 48 | 7.7 | 11.3 | 1.1 | 1.24 |
| SPATA | Year1 | Year2 | 30-Year Average Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tmax | Tmin | Taver | Rainfall | Tmax | Tmin | Taver | Rainfall | Tmax | Tmin | Taver | Rainfall | |
| Nov | 17.6 | 12.3 | 14.8 | 56.8 | 21.3 | 13.9 | 17.2 | 166.6 | 18.2 | 10.4 | 14.7 | 58.8 |
| Dec | 13.5 | 6.8 | 9.8 | 85.6 | 15.3 | 8.9 | 11.8 | 169.6 | 14.2 | 7.1 | 11.1 | 71.2 |
| Jan | 12.3 | 5.5 | 8.7 | 178 | 12.7 | 5.9 | 9.1 | 55.2 | 12.7 | 5.4 | 9.5 | 54.1 |
| Feb | 13.1 | 6.5 | 9.5 | 98.2 | 15.1 | 6.9 | 10.8 | 63.0 | 13.2 | 5.6 | 10.1 | 55.6 |
| Mar | 17.6 | 9.2 | 13.1 | 16.8 | 16.8 | 8.7 | 12.5 | 73.2 | 15.6 | 7.3 | 12.2 | 43.9 |
| Apr | 19.1 | 10.6 | 14.4 | 107 | 19.2 | 10.2 | 14.5 | 50.0 | 19.6 | 10.2 | 15.7 | 28.1 |
| May | 25.2 | 14.4 | 19.3 | 2.8 | 26.7 | 15.0 | 20.3 | 21.4 | 24.8 | 14.3 | 20.4 | 19.1 |
| June | 31.4 | 21.2 | 26.0 | 24.0 | 30.3 | 19.2 | 24.1 | 24.6 | 29.7 | 18.8 | 24.9 | 14.8 |
| LARISSA | ||||||||||||
| Nov | 16.7 | 9.7 | 12.8 | 66.8 | 20.5 | 11.6 | 15.3 | 70.8 | 15.9 | 5.8 | 11.8 | 52.4 |
| Dec | 11.2 | 2.8 | 6.5 | 27.6 | 12.5 | 6.4 | 9.0 | 124 | 11.0 | 1.9 | 7.0 | 50.0 |
| Jan | 8.7 | 1.1 | 4.6 | 68.8 | 16.6 | 1.1 | 6.1 | 1.2 | 9.8 | 0.7 | 5.7 | 32.7 |
| Feb | 14.1 | 4.1 | 8.6 | 19.2 | 16.3 | 4.4 | 9.84 | 17.8 | 12.1 | 1.3 | 7.9 | 39.1 |
| Mar | 19.9 | 6.9 | 13.0 | 19.2 | 18.8 | 6.5 | 12.1 | 57.0 | 14.9 | 3.3 | 10.7 | 40.9 |
| Apr | 21.4 | 10.3 | 15.2 | 18.0 | 21.4 | 9.1 | 14.8 | 48.6 | 19.8 | 6.3 | 15.0 | 26.4 |
| May | 27.5 | 14.7 | 20.4 | 35.4 | 29.1 | 15.2 | 21.3 | 32.0 | 25.8 | 11.0 | 20.3 | 39.6 |
| June | 33.8 | 20.7 | 26.8 | 21.2 | 31.9 | 19.2 | 25.0 | 19.8 | 31.1 | 15.3 | 25.4 | 29.1 |
| THESSALONIKI | ||||||||||||
| Nov | 16.6 | 10.6 | 13.1 | 26.8 | 19.3 | 13.4 | 16.0 | 43.6 | 15.4 | 6.9 | 12.3 | 48.0 |
| Dec | 10.4 | 4.1 | 7.1 | 29.6 | 12.3 | 7.6 | 9.8 | 54.4 | 10.9 | 3.0 | 7.5 | 44.8 |
| Jan | 7.9 | 2.4 | 4.9 | 54.8 | 10.9 | 3.7 | 7.0 | 2.4 | 9.3 | 1.4 | 5.9 | 33.7 |
| Feb | 12.3 | 4.6 | 8.0 | 4.0 | 13.8 | 5.8 | 9.3 | 13.2 | 11.0 | 2.3 | 7.9 | 39.6 |
| Mar | 17.1 | 7.9 | 11.9 | 10.0 | 15.3 | 7.2 | 10.8 | 107 | 14.3 | 4.6 | 10.7 | 47.5 |
| Apr | 18.9 | 10.8 | 14.5 | 73.6 | 18.0 | 9.0 | 13.1 | 81.8 | 19.2 | 7.6 | 14.8 | 31.8 |
| May | 24.6 | 15.0 | 19.3 | 24.2 | 24.3 | 15.1 | 19.1 | 35.0 | 24.5 | 12.3 | 20.0 | 38.4 |
| June | 31.6 | 21.3 | 25.8 | 22.8 | 28.6 | 19.3 | 23.5 | 15.6 | 29.3 | 16.5 | 24.7 | 34.0 |
| Genotype (G) | Environment (E) | GXE | ||||
|---|---|---|---|---|---|---|
| MS | EV% | MS | EV% | MS | EV% | |
| DF | 6 | 5 | 30 | |||
| Agronomic/Colorimetric traits | ||||||
| BY | 23.4 *** | 7.0 | 283.2 *** | 71.5 | 14.2 *** | 21.5 |
| L* | 13.2 *** | 3.5 | 406.0 *** | 89.7 | 5.1 *** | 6.8 |
| a* | 4.9 *** | 10.9 | 39.0 *** | 71.9 | 1.5 *** | 17.2 |
| b* | 3.3 *** | 2.9 | 113.3 *** | 83.8 | 3.0 *** | 13.3 |
| Nutritional traits | ||||||
| NDF | 2691.2 *** | 12.5 | 8486.0 *** | 32.5 | 2405.1 *** | 55.2 |
| ADF | 1206.5 *** | 7.1 | 10,842.1 *** | 52.8 | 1373.4 *** | 40.1 |
| ADL | 280.6 *** | 14.1 | 798.6 *** | 33.5 | 207.9 *** | 52.4 |
| CP | 9597.1 *** | 16.6 | 52,382.6 *** | 75.7 | 878.8 *** | 7.6 |
| Antinutritional traits | ||||||
| TP | 2.8 *** | 2.6 | 106.2 *** | 82.5 | 3.2 *** | 14.9 |
| TN | 1.7 *** | 4.1 | 35.6 *** | 72.6 | 1.9 *** | 23.3 |
| CT | 0.03 *** | 5.0 | 0.5 *** | 75.3 | 2.2 *** | 19.7 |
| OA | 121.8 *** | 3.4 | 3661.7 *** | 84.3 | 82.1 *** | 12.3 |
| Genotypes | Tukey (0.05) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| G1 | G2 | G3 | G4 | G5 | G6 | G7 | |||
| Biomass/colorimetric traits | BY | 12.2 b | 13.2 bc | 11.3 a | 13.1 abc | 12.7 ab | 11.6 ab | 14.6 c | 3.568 |
| L* | 47.6 a | 47.9 b | 48.5 c | 47.5 a | 47.9 b | 50.0 d | 48.6 c | 0.095 | |
| a* | 2.6 a | 3.4 c | 4.0 e | 2.7 a | 3.8 d | 3.4 c | 3.2 b | 0.130 | |
| b* | 21.9 b | 21.6 a | 22.3 c | 22.2 c | 22.0 b | 22.9 e | 22.6 d | 0.320 | |
| Nutritional traits | NDF | 346.0 b | 353.4 bc | 358.5 c | 368.1 d | 350.6 bc | 356.1 c | 328.7 a | 76.332 |
| ADF | 229.2 b | 228.7 b | 239.2 c | 243.5 c | 225.7 ab | 221.3 a | 223.8 ab | 30.297 | |
| ADL | 59.0 c | 54.4 b | 58.5 c | 60.2 c | 57.0 bc | 50.7 a | 50.7 a | 13.142 | |
| CP | 229.2 e | 190.6 c | 170.4 a | 217.5 d | 185.3 b | 167.5 a | 184.6 b | 15.791 | |
| Antinutritional traits | TP | 9.9 bc | 9.7 b | 10.4 d | 10.5 d | 9.4 a | 9.7 b | 10.1 c | 0.630 |
| TN | 4.8 cd | 4.7 bc | 5.1 de | 5.2 e | 4.5 ab | 4.3 a | 4.7 bc | 0.490 | |
| CT | 0.39 e | 0.34 b | 0.38 de | 0.27 a | 0.37 cde | 0.35 bc | 0.37 cde | 0.001 | |
| OA | 39.9 c | 41.1 d | 43.2 e | 43.5 e | 39.7 c | 36.4 a | 38.1 b | 1.087 | |
| Environments | Tukey (0.05) | |||||||
|---|---|---|---|---|---|---|---|---|
| E1 | E2 | E3 | E4 | E5 | E6 | |||
| Biomass/Colorimetric traits | BY | 19.6 d | 11.5 b | 10.9 ab | 11.2 b | 9.2 a | 13.8 c | 3.568 |
| L* | 50.7 e | 43.1 a | 48.0 c | 54.9 f | 49.2 d | 43.8 b | 0.095 | |
| a* | 4.1 d | 4.8 e | 4.5 d | 2.1 b | 1.6 a | 2.7 c | 0.130 | |
| b* | 24.1 e | 20.3 b | 22.4 c | 24.8 f | 23.1 d | 18.7 a | 0.320 | |
| Nutritional traits | NDF | 360.6 d | 343.3 bc | 387.1 e | 350.1 c | 331.1 a | 337.8 ab | 76.332 |
| ADF | 229.8 b | 209.7 a | 269.4 e | 242.4 d | 216.3 b | 213.6 ab | 30.297 | |
| ADL | 51.1 a | 51.1 a | 53.1 a | 52.2 a | 64.5 b | 62.8 b | 13.142 | |
| CP | 161.1 b | 256.6 f | 128.4 a | 169.2 c | 193.1 d | 244.6 e | 15.791 | |
| Antinutritional traits | TP | 6.7 a | 10.3 d | 8.7 b | 13.5 f | 10.6 e | 9.9 c | 0.630 |
| TN | 2.7 a | 5.3 d | 3.9 b | 6.4 f | 4.7 c | 5.5 e | 0.790 | |
| CT | 0.15 a | 0.35 c | 0.22 b | 0.60 e | 0.35 c | 0.46 d | 0.001 | |
| OA | 33.5 b | 54.5 e | 52.2 d | 26.4 a | 49.2 c | 25.8 a | 1.087 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parissi, Z.; Irakli, M.; Tigka, E.; Papastylianou, P.; Dordas, C.; Tani, E.; Abraham, E.M.; Theodoropoulos, A.; Kargiotidou, A.; Kougiteas, L.; et al. Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch. Agronomy 2022, 12, 1678. https://doi.org/10.3390/agronomy12071678
Parissi Z, Irakli M, Tigka E, Papastylianou P, Dordas C, Tani E, Abraham EM, Theodoropoulos A, Kargiotidou A, Kougiteas L, et al. Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch. Agronomy. 2022; 12(7):1678. https://doi.org/10.3390/agronomy12071678
Chicago/Turabian StyleParissi, Zoi, Maria Irakli, Evangelia Tigka, Panayiota Papastylianou, Christos Dordas, Eleni Tani, Eleni M. Abraham, Agisilaos Theodoropoulos, Anastasia Kargiotidou, Leonidas Kougiteas, and et al. 2022. "Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch" Agronomy 12, no. 7: 1678. https://doi.org/10.3390/agronomy12071678
APA StyleParissi, Z., Irakli, M., Tigka, E., Papastylianou, P., Dordas, C., Tani, E., Abraham, E. M., Theodoropoulos, A., Kargiotidou, A., Kougiteas, L., Kousta, A., Koskosidis, A., Kostoula, S., Beslemes, D., & Vlachostergios, D. N. (2022). Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch. Agronomy, 12(7), 1678. https://doi.org/10.3390/agronomy12071678