
Sulla Powdery Mildew: Phylogeny and Host Range




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Material
2.2. Morphology Assessments
2.3. DNA Extraction and ITS Sequencing
2.4. Phylogenetic Study
2.5. Host Range
3. Results
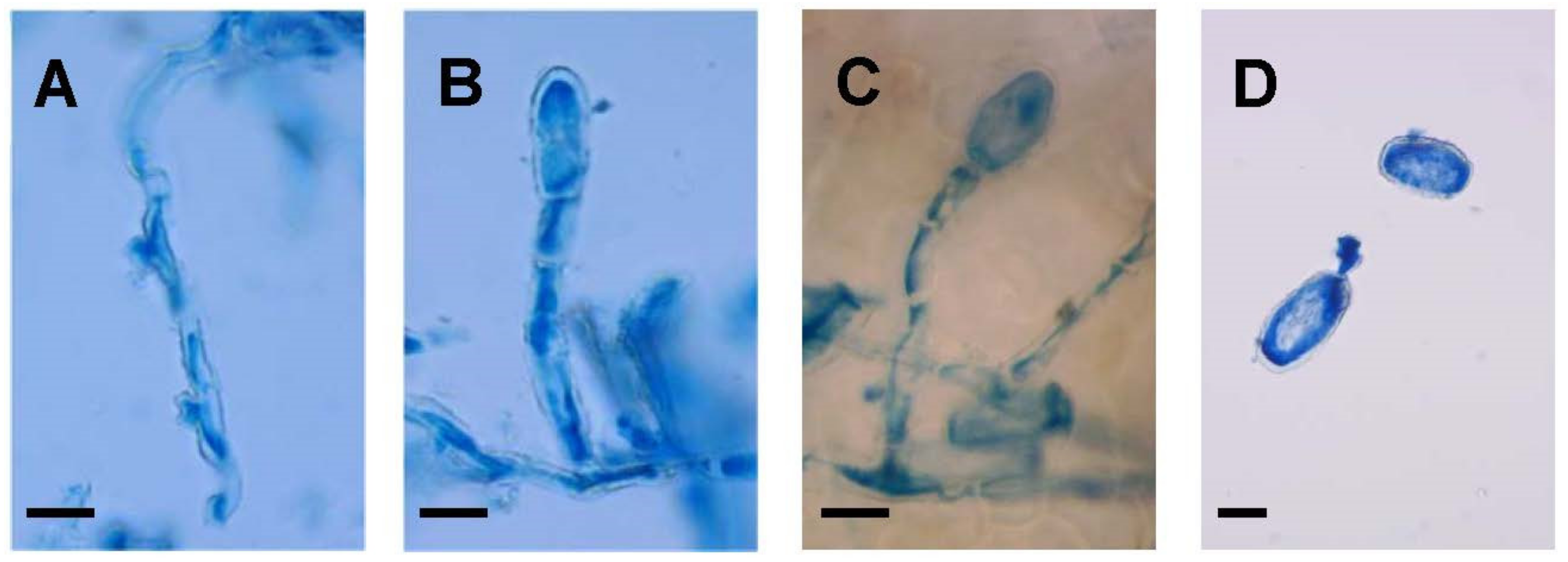
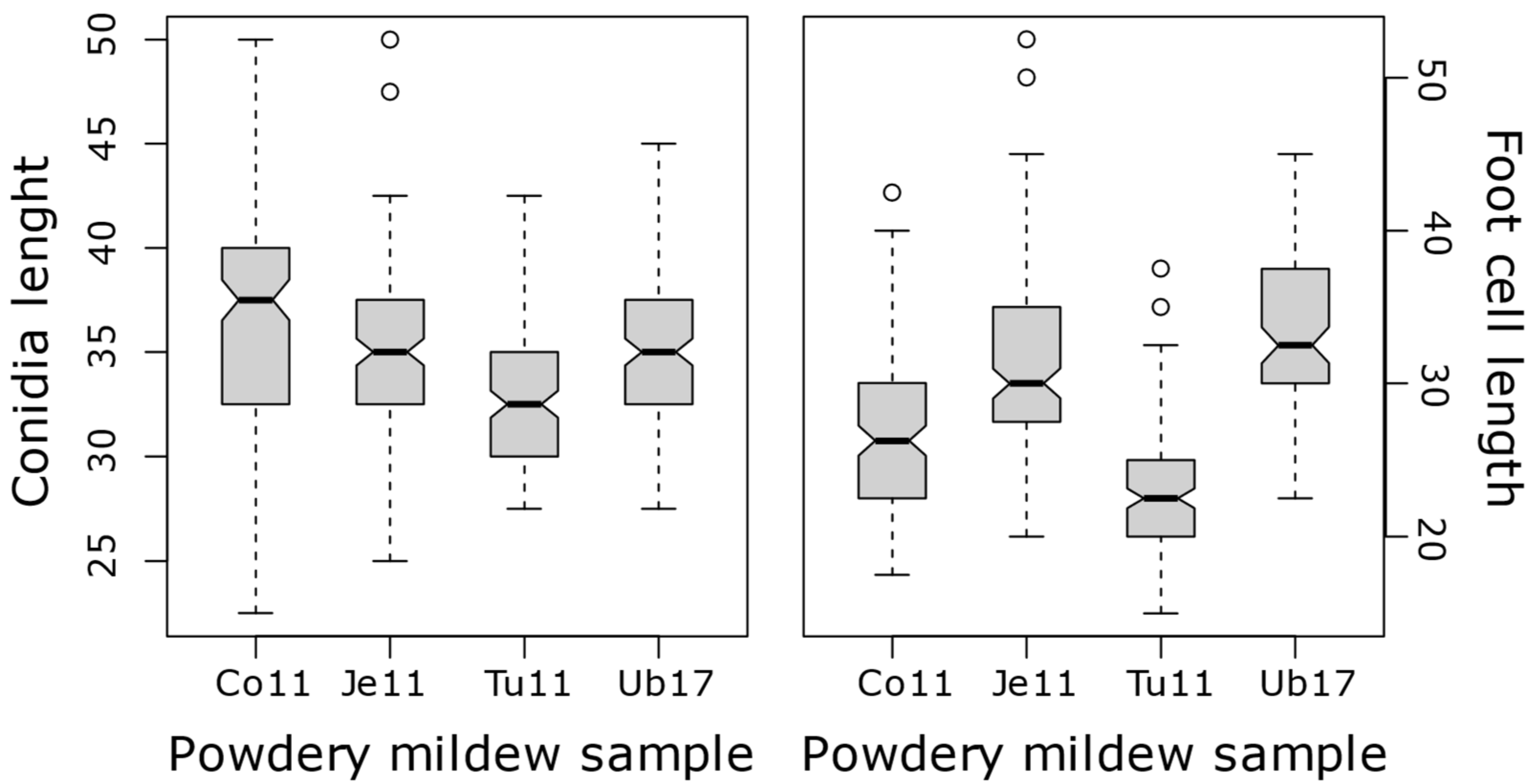
3.1. Mycelial Morphology
3.2. ITS Amplification
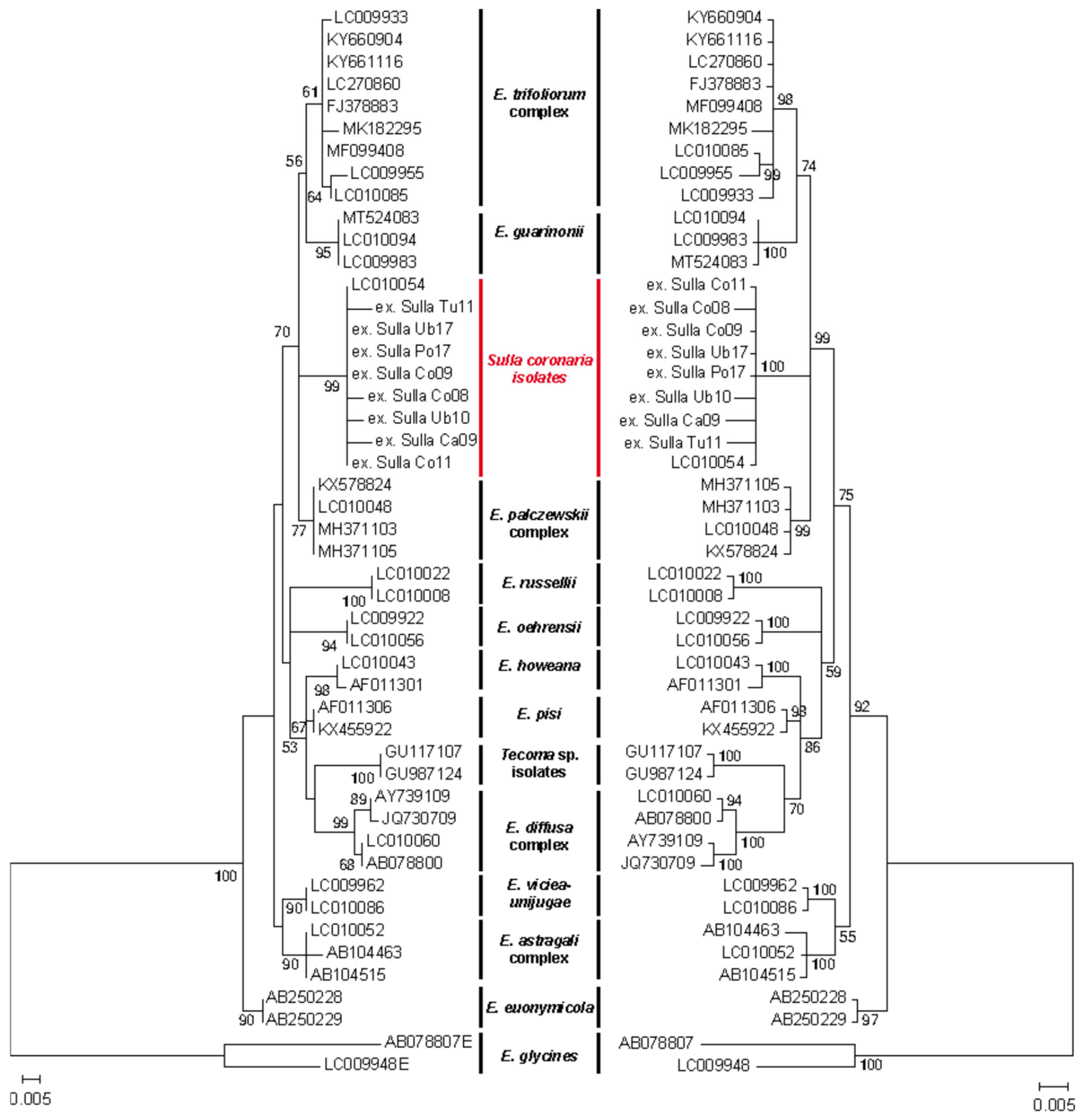
3.3. Phylogenetic Relationship
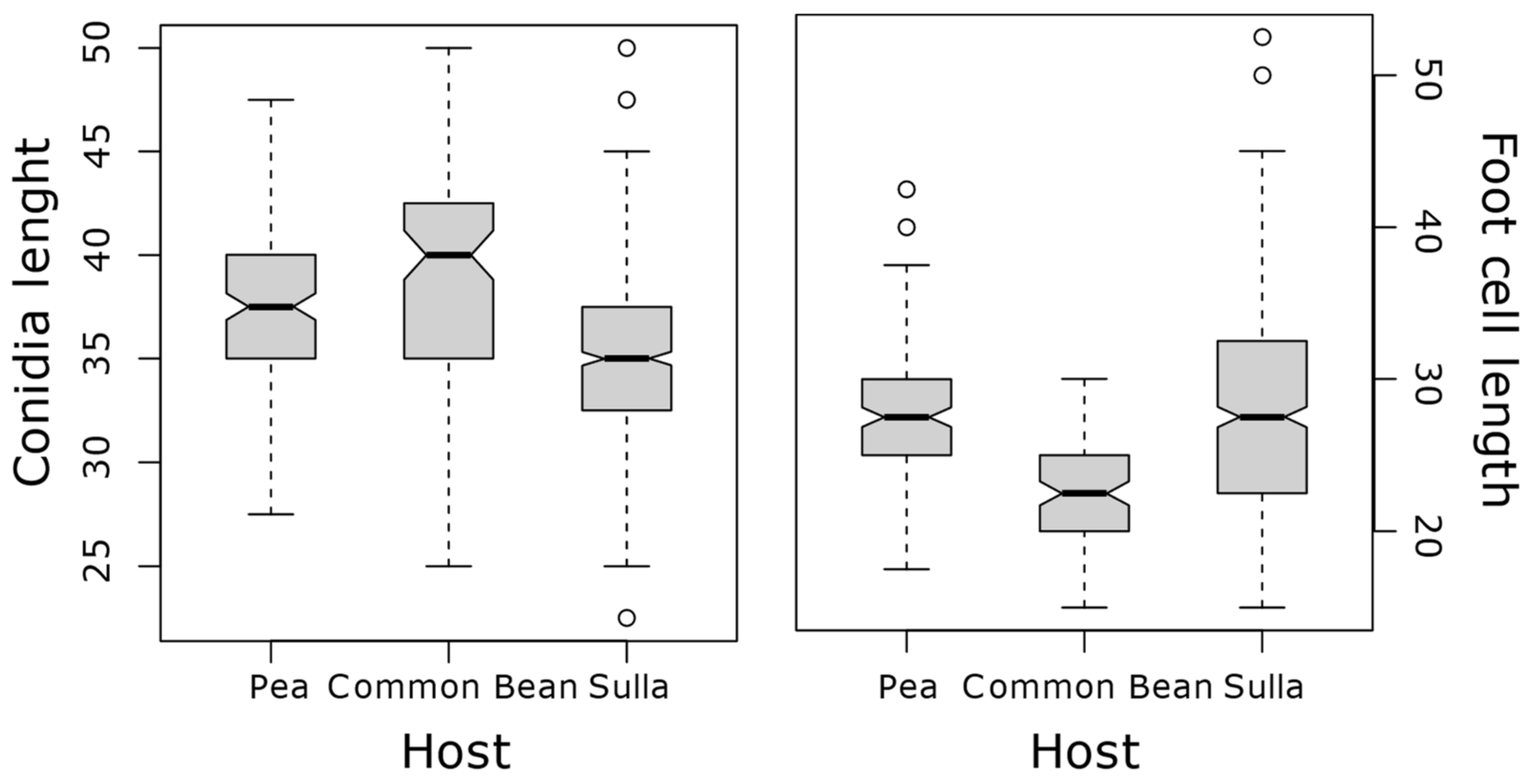
3.4. Host Range
4. Discussions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruisi, P.; Siragusa, M.; Di Giorgio, G.; Graziano, D.; Amato, G.; Carimi, F.; Giambalvo, D. Pheno-morphological, agronomic and genetic diversity among natural populations of sulla (Hedysarum coronarium L.) collected in Sicily, Italy. Gen. Res. Crop Evol. 2011, 58, 245–257. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosuè, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, M.M.; Mojaradi, T.; Shirzadi, T. Effects of Hedysarum coronarium L. (sulla) as a green manure along with nitrogen fertilizer on maize Production. Agritech 2021, 41, 95–106. [Google Scholar] [CrossRef]
- Erwin, M.; Foster, K.; Skinner, P.; Nichols, P.; Nutt, B.; Snowball, R.; Beatty, R. Sulla and Purple Clover as Fodders. Limits on Seed Production. RIRDC Publication No 06/049. 2006. Available online: https://www.agrifutures.com.au/wp-content/uploads/publications/06-049.pdf (accessed on 18 April 2022).
- Crocker, G.; Hackney, B. Sulla. Primefact 745. State of New South Wales through NSW Department of Primary Industries, 2008. ISSN 1832-6668. Available online: www.dpi.nsw.gov.au/primefacts (accessed on 29 April 2022).
- Leto, G.; Todaro, M.; di Noto, A.M.; Alicata, M.L. Comparison of sulla-hay and sulla-silage in the lactating ewes and their effects on milk and cheese characteristics. Small Rum. Res. 2002, 45, 301–306. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Ruisi, P.; Di Miceli, G.; Pecetti, L. Morpho-physiological and adaptive variation of Italian germplasm of sulla (Hedysarum coronarium L.). Crop Pasture Sci. 2014, 65, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Córdoba, E.M.; Nadal, S.; Román, B.; Gónzalez-Verdejo, C.I. Collection, characterization and evaluation of wild (Hedysarum coronarium L.) populations from Andalusia (southern Spain). Aust. J. Crop Sci. 2013, 7, 165–172. [Google Scholar]
- Flores, F.; Gutiérrez, J.; López, J.; Moreno, M.T.; Cubero, M.T. Multivariate analysis approach to evaluate a germplasm collection of Hedysarum coronarium L. Gen. Resour. Crop Evol. 1997, 44, 545–555. [Google Scholar] [CrossRef]
- Marghali, S.; Panaud, O.; Lamy, F.; Ghariani, S.; Sarr, A.; Marrakchi, M.; Trifi-Farah, N. Exploration of intra- and inter-population genetic diversity in Hedysarum coronarium L. by AFLP markers. Gen. Resour. Crop Evol. 2005, 52, 277–284. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Prats, E.; Emeran, A.A.; Rubiales, D. Characterisation of resistance mechanisms to Erysiphe betae in Beta vulgaris. Phytopathology 2009, 99, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Fondevilla, S.; Rubiales, D. Powdery mildew control in pea. A review. Agron. Sustain. Devel. 2012, 32, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Trabanco, N.; Pérez-Vega, E.; Campa, A.; Rubiales, D.; Ferreira, J.J. Genetic resistance to powdery mildew in common bean. Euphytica 2012, 186, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Louati-Namouchi, I.; Louati, M.; Chriki, A. A quantitative study of some agronomic characters in Sulla (Hedysarum coronarium L.). Agronomie 2000, 20, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Braun, U.; Takamatsu, S. Phylogeny of Erysiphe, Microsphaera, Uncinula (Erysipheae) and Cystotheca, Podosphaera, Sphaerotheca (Cystotheceae) inferred from rDNA ITS sequences—Some taxonomic consequences. Schlechtendalia 2000, 4, 1–33. [Google Scholar]
- Braun, U.; Kruse, J.; Wolcan, S.M.; Murace, M. Three new species of the genus Erysiphe (Ascomycota, Erysiphales) on legumes and some new combinations. Mycotaxon 2010, 112, 173–187. [Google Scholar] [CrossRef]
- Watson, M.J. Hedysarum coronarium—A legume with potential for soil conservation and forage. N. Z. J. Agric. Res. 1982, 16, 189–193. [Google Scholar]
- Chen, M.M. Forest Fungi Phytogeography: Forest Fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research & Education Center: Sacramento, CA, USA, 2003; ISBN 0-9723916-6-5. [Google Scholar]
- Braun, U. Descriptions of new species and combinations in Microsphaera and Erysiphe (V). Mycotaxon 1984, 19, 374–383. [Google Scholar]
- Rubiales, D.; Carver, T.W.L. Defence reactions of Hordeum chilense accessions to three formae speciales of cereal powdery mildew fungi. Can. J. Bot. 2000, 78, 1561–1570. [Google Scholar] [CrossRef]
- Attanayake, R.N.; Glawe, D.A.; Dugan, F.M.; Chen, W. Erysiphe trifolii causing powdery mildew of lentil (Lens culinaris). Plant Dis. 2009, 93, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Lebeda, A.; Mieslerova, B.; Sedlarova, M. First report of Erysiphe palczewskii on Caragana arborescens in the Czech Republic. Plant Pathol. 2008, 57, 779. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Notes on powdery mildews (Erysiphales) in Japan: I. Erysiphe Sect. Erysiphe. Mycoscience 2015, 56, 257–266. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Notes on powdery mildews (Erysiphales) in Thailand III. Erysiphe species on Fabaceae, Fagaceae, Hydrangeaceae and Lamiaceae. Tropical Plant Pathol. 2017, 42, 239–249. [Google Scholar] [CrossRef]
- Qiu, P.; Tang, S.; Guan, G.; Li, Y.; Takamatsu, S.; Liu, S. Revisiting a poorly known powdery mildew species: Morphology and molecular phylogeny of Erysiphe longissima. Mycoscience 2019, 60, 250–254. [Google Scholar] [CrossRef]
- Raeder, U.; Broda, P. Rapid preparation of DNA from filamentous fungi. Lett. Appl. Microbiol. 1985, 1, 17–20. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Prats, E.; Llamas, M.J.; Rubiales, D. Characterisation of resistance mechanisms to Erysiphe pisi in Medicago truncatula. Phytopathology 2007, 97, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martín, J.; Rubiales, D.; Prats, E. Resistance to powdery mildew (Blumeria graminis f.sp. avenae) in oat seedlings and adult plants. Plant Pathol. 2011, 60, 846–856. [Google Scholar]
- Takamatsu, S.; Ito, H.A.; Shiroya, Y.; Kiss, L.; Heluta, V. First comprehensive phylogenetic analysis of the genus Erysiphe (Erysiphales, Erysiphaceae) I. The Microsphaera lineage. Mycologia 2015, 107, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, S. Studies on the evolution and systematics of powdery mildew fungi. J. Gen. Plant Pathol. 2018, 84, 422–426. [Google Scholar] [CrossRef]
- Mohan, S.K.; Bijman, V.P.; Miller, D.R. Powdery Mildew Caused by Erysiphe pisi on Alfalfa in Idaho and Oregon. Plant Dis. 2001, 85, 678. [Google Scholar] [CrossRef]
- Cook, R.T.A.; Fox, R.T.V. Erysiphe pisi var. pisi on faba beans and other legumes in Britain. Plant Pathol. 1992, 41, 506–512. [Google Scholar] [CrossRef]
- Dugan, F.M.; Attanayake, K.; Glawe, D.; Chen, W. Powdery Mildew of Chickpea. Compendium of Chickpea and Lentil Diseases and Pests; American Phytopathological Society: Saint Paul, MN, USA, 2011; pp. 51–52. [Google Scholar]
- Leitão, S.T.; Almeida, N.F.; Moral, A.; Rubiales, D.; Vaz Patto, M.C. Identification of resistance to rust (Uromyces appendiculatus) and powdery mildew (Erysiphe difussa) in Portuguese common bean germplasm. Plant Breed. 2013, 132, 654–657. [Google Scholar] [CrossRef]
- Worku, J. Effects of Powdery Mildew (Leveillula Taurica and Erysiphe Polygoni) on yield and yield components of fenugreek in the mid-altitudes of Bale, South Eastern Ethiopia. J. Plant. Sci. 2017, 5, 65–67. [Google Scholar] [CrossRef]
- Bradshaw, M.; Braun, U.; Götz, M.; Jurick, W. Phylogeny and taxonomy of powdery mildew caused by Erysiphe species on Lupinus hosts. Mycologia 2022, 114, 76–88. [Google Scholar] [CrossRef]
- Rubiales, D.; Sillero, J.C.; Emeran, A.A. Response of vetches (Vicia spp.) to specialized forms of Uromyces vicia-fabae and to Uromyces pisi. Crop Prot. 2013, 46, 38–43. [Google Scholar]
- Maxted, N. An ecogeographical study of Vicia subgenus Vicia. In Systematic and Ecogeographic Studies on Crop Genepools; International Plant Genetic Resources Institute: Rome, Italy, 1995; Volume 8. [Google Scholar]
- Leppik, E.E. Evolutionary specialization of rust fungi (Uredinales) on the Leguminosae. Ann. Bot. Fenn. 1972, 9, 135–148. [Google Scholar]
- Van der Merwe, M.; Walker, J.; Ericson, L.; Burdon, J.J. Coevolution with higher taxonomic host groups within the Puccinia/Uromyces rust lineage obscured by host jumps. Mycol. Res. 2008, 112, 1387–1408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Powdery Mildew | Sample | Origin | Genbank |
|---|---|---|---|---|
| Sulla coronaria | Co08 | Córdoba, Spain | ON729986 | |
| S. coronaria | Co09 | Córdoba, Spain | ON729988 | |
| S. coronaria | Co11 | Córdoba, Spain | ON729985 | |
| S. coronaria | Ca09 | Tarifa, Spain | ON729987 | |
| S. coronaria | Ub10 | Ubrique, Spain | ON729991 | |
| S. coronaria | Je11 | Jerez de la Frontera, Spain | ||
| S. coronaria | Tu11 | Hammemet, Tunisia | ON729989 | |
| S. coronaria | Ub17 | Ubrique, Spain | ON729990 | |
| S. coronaria | Po17 | Oerias, Portugal | ON729992 | |
| Pisum sativum | E. pisi | Ps11 | Córdoba, Spain | |
| Phaseolus vulgaris | E. diffusa | Pv11 | Oerias, Portugal | |
| Lathyrus sativus | E. trifoliorum ex. L. sativus | Córdoba, Spain | ||
| Medicago truncatula | E. trifoliorum ex. M. truncatula | Córdoba, Spain | ||
| Vicia articulata | E. trifoliorum ex. V. articulata | Córdoba, Spain | ||
| Albizia julibrissin | Erysiphe cf. trifoliorum | MUMH0133 | Japan | LC010085 |
| Alhagi sp. | E. bremeri | AK113 | Guilan, Iran | AB104463 |
| Astragalus glycyphyllus | E. astragali | MUMH2585 | Ukraine | LC010052 |
| Astragalus sp. | E. astragali | Guilan, Iran | AB104515 | |
| Baptisia australis | E. guarinonii | HAL2337 | Germany | MT524083 |
| Caragana arborescens | E. palczewskii | MUMH2581 | Ukraine | LC010048 |
| C. rosea | E. longissima | HMJAU91780 | China | MH371103 |
| C. rosea | E. longissima | HMJAU91781 | China | MH371105 |
| Coronilla varia | Pseudoidium sp. | MUMH2587 | Ukraine | LC010054 |
| Desmodium incanum | E. diffusa | MUMH3121 | Corrientes, Argentina | LC010060 |
| D. laxum | E. glycines | MUMH396 | Shiga, Japan | LC009948 |
| Euonymus maackii | E. euonymi | HMJAU91794 | China | MK182295 |
| E. japonica | E. euonymicola | MUMH133 | Japan | AB250228 |
| E. japonica | E. euonymicola | MUMH2470 | Argentina | AB250229 |
| Glycine max | E. diffusa | MUMH791 | Oita, Japan | AB078800 |
| G. max | E. glycines | MUMH1462 | Mie, Japan | AB078807 |
| Hardenbergia sp. | P. hardenbergia | VPRI19879 | Australia | LC010094 |
| Hypericum ascyron | E. hypericin | F29454 | Jinan, Korea | MF099408 |
| Labrunum alpinum | E. guarinonii | MUMH1425 | Switzerland | LC009983 |
| Lathyrus latifolium | E. pisi | Belden, USA | AF011306 | |
| L. pratensis | E. ludens | OE2016PMCS45 | Balerno, UK | KY661116 |
| Lens culinaris | E. trifolii | LGH06 | Pullman, USA | FJ378883 |
| Lupinus sp. | E. intermedia | OE2015PMCS297 | Chelmsford, UK | KY660904 |
| Maytenus boaria | E. oehrensii | MUMH2492 | Bariloche, Argentina | LC010022 |
| M. boaria | E. oehrensii | MUMH1936 | Bariloche, Argentina | LC010008 |
| Medicago littoralis | E. trifoliorum | MUMH7038 | Baku, Azerbaijan | LC270860 |
| Oenothera amoena | E. howeana | MUMH2572 | The Netherlands | LC010043 |
| O. biennis | E. howeana | UC1512301 | Redlands, USA | AF011301 |
| Oxalis corniculatus | E. russellii | MUMH0105 | Mie, Japan | LC009922 |
| Phaseolus vulgaris | Erysiphe sp. | EB2004 | Londrina, Brazil | AY739109 |
| Pisum sativum | E. pisi | P1 | India | KX455922 |
| Robinia pseudoacacia | E. palczewskii | ZKEP001 | China | KX578824 |
| Senna septemtrionalis | Erysiphe sp. | 3D | Buenavista, Mexico | JQ730709 |
| Tecoma capensis | Erysiphe sp. | DAG08-36 | Yuma, USA | GU117107 |
| T. capensis | Erysiphe sp. | 08-3618S | Yuma, USA | GU987124 |
| Trifolium arvense | E. trifoliorum | MUMH0701 | Budapest, Hungria | LC009955 |
| Vicia hirsute | E. baeumleri | MUMH0240 | Shiga, Japan | LC009933 |
| V. angustifolius | E. viciae-unijugae | MUMH0817 | Yamanashi, Japan | LC009962 |
| V. unijuga | E. viciae-unijugae | TPU-153 | Japan | LC010086 |
| Xanthoxalis sp. | E. russellii | MUMH2593 | Ukraine | LC010056 |
| Erysiphe sp. ex. Sulla coronaria | E. pisi Ps11 | E. diffusa Pv11 | E. palcewskii [23] | E. longissimi [26] | E. trifoliorum [24] | E. pisi [25] | E. diffusa [22] | |
|---|---|---|---|---|---|---|---|---|
| conidia | ||||||||
| Length (µm) | 22.5–50 | 27.5–47.5 | 25–50 | 20–37.5 | 19.2–48.2 | 25.5–39.5 | 30–44 | 28–36 |
| Width (µm) | 10–22.5 | 12.5–20 | 15–22.5 | 8.75–17.5 | 12.3–17.2 | 12–18 | 18–20 | 12.5–16 |
| Ratio L/W | 1–5 | 1.4–3.8 | 1.1–3.3 | 1.54–3.14 | 1.4–2.9 | 1.4–3.3 | 1.5–2.4 | 1.8–2.9 |
| Foot cell | ||||||||
| Length (µm) | 15–52.5 | 17.5–42.5 | 15–30 | 15–40 | 24.5–54.2 | 13–45 | 35–70 | 25–33 |
| Width (µm) | 5–12.5 | 5–12.5 | 7.5–10 | nd | 6–9.1 | 7–9.5 | 8–12.5 | 7.7–9 |
| Ratio L/W | 1.2–10.5 | 1.4–8.5 | 1.5–4 | nd | 2.69–9.03 | 1.4–6.4 | 2.8–8.75 | 2.8–4.3 |
| Co08 | Co09 | Co11 | Ca08 | Tu11 | Ub17 | Ub10 | Po17 | |
|---|---|---|---|---|---|---|---|---|
| Co08 | 100 | |||||||
| Co09 | 99.4 | 100 | ||||||
| Co11 | 99.2 | 99.7 | 100 | |||||
| Ca09 | 98.9 | 99.1 | 99.1 | 100 | ||||
| Tu11 | 98.9 | 99.2 | 99.2 | 98.8 | 100 | |||
| Ub17 | 99.2 | 99.7 | 99.7 | 99.2 | 99.2 | 100 | ||
| Ub10 | 99 | 99.7 | 99.6 | 99 | 99.1 | 99.7 | 100 | |
| Po17 | 99.4 | 99.9 | 99.9 | 99.3 | 99.4 | 99.9 | 99.7 | 100 |
| Response* to Powdery Mildew Isolate | |||||||
|---|---|---|---|---|---|---|---|
| Crop | Host specie | Number of accessions studied | Erysiphe sp. ex S. coronaria | E. pisi ex P. sativum | E. trifoliorum ex L. sativus | E. trifoliorum ex M. truncatula | E. trifoliorum ex V. articulata |
| Sulla | Sulla coranaria | 5 | xx/xxx | - | - | - | - |
| Pea and related sp. | Pisum sativum | 6 | - | xx/xxx | xx/xxx | -/x | x |
| Pisum fulvum | 2 | - | - | xxx | -/x | - | |
| Lentil | Lens culinaris | 5 | - | - | xx/xxx | x | xxx |
| Grasspea and related sp. | Lathyrus sativus | 5 | - | x/xx | xxx | - | - |
| Lathyrus cicera | 6 | - | x/xxx | x/xxx | - | - | |
| Lathyrus ochrus | 3 | - | -/x | xxx | - | xx | |
| Lathyrus clymenum | 2 | - | -/x | xxx | - | ns | |
| Vetches | Vicia articulata | 1 | - | - | xx | - | xxx |
| Vicia monantha | 2 | - | x | ns | - | xxx | |
| Vicia villosa | 1 | - | ns | - | - | xx | |
| Vicia sativa | 5 | - | - | -/x | - | -/x | |
| Vicia pannonica | 1 | - | - | x | - | - | |
| Vicia tetrasperma | 1 | - | - | - | - | - | |
| Faba bean | Vicia faba | 5 | - | - | - | - | - |
| Fenugreek | Trigonella foenun-graecum | 6 | - | - | -/xx | -/xx | - |
| Annual medics | Medicago truncatula | 1 | - | ns | x | xxx | ns |
| Medicago polymorpha | 1 | - | - | x | xxx | ns | |
| Alfalfa | Medicago sativa | 4 | - | - | - | - | ns |
| Chickpea and related sp. | Cicer arietinum | 5 | - | - | - | - | - |
| Cicer reticulatum | 2 | - | - | - | - | ns | |
| Common bean and related sp. | Phaseolus vulgaris | 9 | - | - | - | - | - |
| Phaselus filiformis | 2 | - | - | - | - | ns | |
| Phaseolus leptostachyus | 2 | - | - | - | - | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubiales, D.; Moral, A.; Rispail, N. Sulla Powdery Mildew: Phylogeny and Host Range. Agronomy 2022, 12, 1852. https://doi.org/10.3390/agronomy12081852
Rubiales D, Moral A, Rispail N. Sulla Powdery Mildew: Phylogeny and Host Range. Agronomy. 2022; 12(8):1852. https://doi.org/10.3390/agronomy12081852
Chicago/Turabian StyleRubiales, Diego, Ana Moral, and Nicolás Rispail. 2022. "Sulla Powdery Mildew: Phylogeny and Host Range" Agronomy 12, no. 8: 1852. https://doi.org/10.3390/agronomy12081852
APA StyleRubiales, D., Moral, A., & Rispail, N. (2022). Sulla Powdery Mildew: Phylogeny and Host Range. Agronomy, 12(8), 1852. https://doi.org/10.3390/agronomy12081852