
Genome-Wide Identification and Characterization of YABBY Gene Family in Juglans regia and Juglans mandshurica


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of YABBY Transcription Factors in J. regia and J. mandshurica
2.2. Characteristics and Phylogenetic Relationships of Identified YABBYs
2.3. Cis-Acting Element Prediction and Gene Expression Analysis
3. Results
3.1. Genome-Wide Identification and Phylogenetic Analysis of YABBY Gene Family in J. regia and J. mandshurica
3.2. Physicochemical Properties and Subcellular Localization Analysis of YABBY Proteins in J. regia and J. mandshurica
3.3. Protein Domain and Gene Structure Distribution Analysis of YABBY Genes in J. regia and J. mandshurica
3.4. Chromosomal Distribution and Duplication Mode Analysis of YABBY Gene Family in J. regia and J. mandshurica
3.5. Collinearity and Selective Pressure Analysis of YABBY Members in J. regia and J. mandshurica
3.6. Analysis of Cis-Acting Elements in J. regia and J. mandshurica
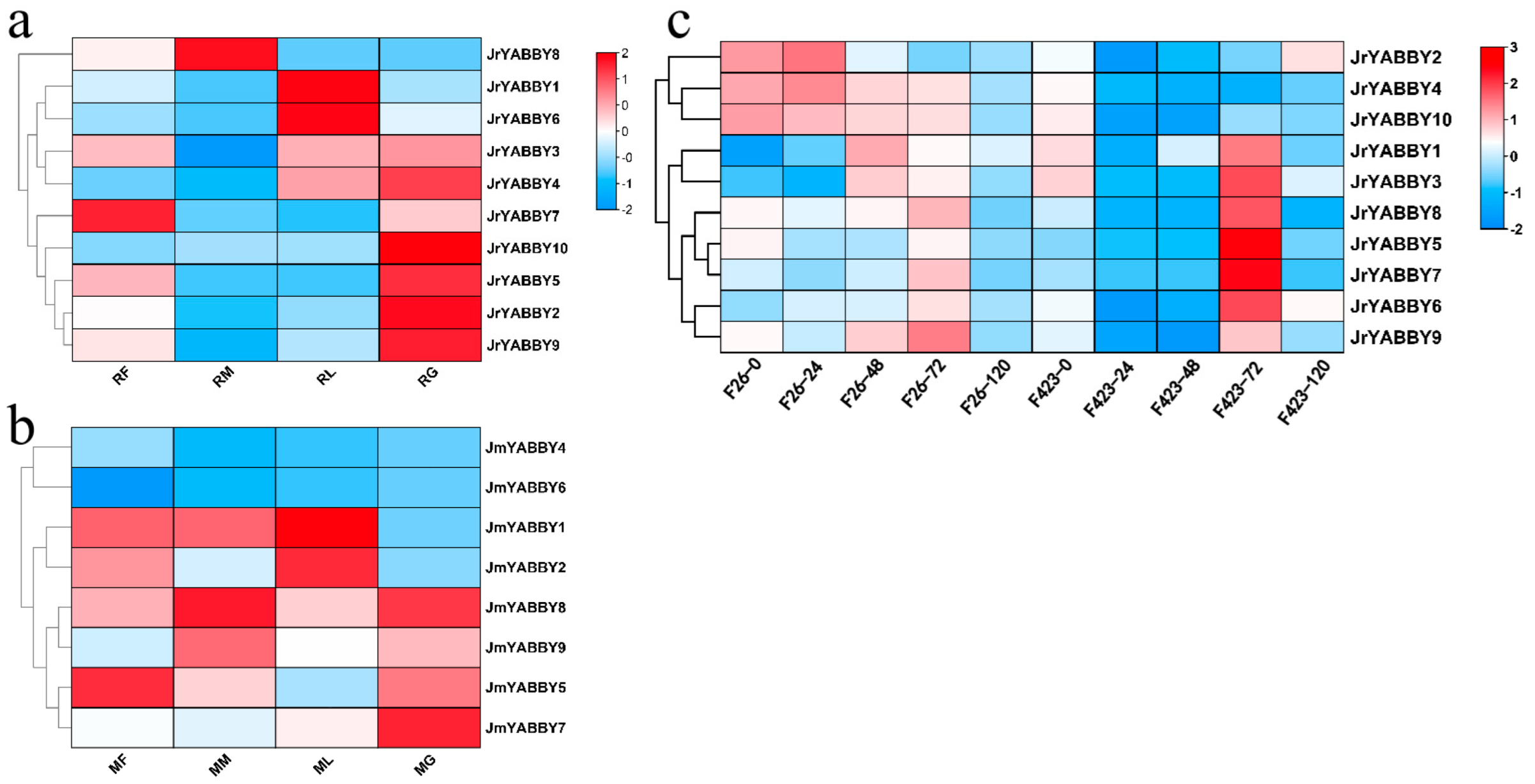
3.7. Gene Expression Profile Analysis of YABBY Genes in J. regia and J. mandshurica
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.; Liao, X.-Y.; Zheng, Y.; Zhu, M.-J.; Yu, X.; Jiang, Y.-T.; Zhang, D.-Y.; Ma, L.; Xu, X.-Y.; Liu, Z.-J.; et al. Genome-Wide Identification of the YABBY Gene Family in Seven Species of Magnoliids and Expression Analysis in Litsea. Plants 2021, 10, 21. [Google Scholar] [CrossRef]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q.; et al. Genome-Wide Study of YABBY Genes in Upland Cotton and Their Expression Patterns under Different Stresses. Front. Genet. 2018, 9, 33. [Google Scholar] [CrossRef]
- Buttar, Z.A.; Yang, Y.; Sharif, R.; Nan Wu, S.; Xie, Y.; Wang, C. Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.). Agronomy 2020, 10, 1189. [Google Scholar] [CrossRef]
- Shamimuzzaman, M.; Vodkin, L. Genome-wide identification of binding sites for NAC and YABBY transcription factors and co-regulated genes during soybean seedling development by ChIP-Seq and RNA-Seq. BMC Genom. 2013, 14, 477. [Google Scholar] [CrossRef]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.-Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet. Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Niu, Y.; Gao, R.; Wang, C.; Liao, W. Genome-Wide Identification and Expression Analysis of SnRK Gene Family under Abiotic Stress in Cucumber (Cucumis sativus L.). Agronomy 2022, 12, 1550. [Google Scholar] [CrossRef]
- di Rienzo, V.; Imanifard, Z.; Mascio, I.; Gasser, C.S.; Skinner, D.J.; Pierri, C.L.; Marini, M.; Fanelli, V.; Sabetta, W.; Montemurro, C.; et al. Functional conservation of the grapevine candidate gene INNER NO OUTER for ovule development and seed formation. Hortic Res. 2021, 8, 29. [Google Scholar] [CrossRef]
- Finet, C.; Floyd, S.K.; Conway, S.J.; Zhong, B.; Scutt, C.P.; Bowman, J.L. Evolution of the YABBY gene family in seed plants. Evol. Dev. 2016, 18, 116–126. [Google Scholar] [CrossRef]
- Tanaka, W.; Toriba, T.; Hirano, H.-Y. Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice. New Phytol. 2017, 215, 825–839. [Google Scholar] [CrossRef]
- Xia, J.; Wang, D.; Peng, Y.; Wang, W.; Wang, Q.; Xu, Y.; Li, T.; Zhang, K.; Li, J.; Xu, X. Genome-Wide Analysis of the YABBY Transcription Factor Family in Rapeseed (Brassica napus L.). Genes 2021, 12, 981. [Google Scholar] [CrossRef]
- Eckardt, N.A. YABBY Genes and the Development and Origin of Seed Plant Leaves. Plant Cell 2010, 22, 2103. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Liu, R.Q.; Li, T.M.; Han, L.J.; Zou, Y.; Xu, T.F.; Wei, J.Y.; Wang, Y.J.; Xu, Y. Isolation and characterization of two VpYABBY genes from wild Chinese Vitis pseudoreticulata. Protoplasma 2013, 250, 1315–1325. [Google Scholar] [CrossRef]
- She, Z.; Huang, X.; Aslam, M.; Wang, L.; Yan, M.; Qin, R.; Chen, Y.; Qin, Y.; Niu, X. Expression characterization and cross-species complementation uncover the functional conservation of YABBY genes for leaf abaxial polarity and carpel polarity establishment in Saccharum spontaneum. BMC Plant Biol. 2022, 22, 124. [Google Scholar] [CrossRef] [PubMed]
- Cong, B.; Barrero, L.S.; Tanksley, S.D. Regulatory change in YABBY-like transcription factor led to evolution of extreme fruit size during tomato domestication. Nat. Genet. 2008, 40, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Won, S.Y.; Park, D.S.; Lee, Y.-H.; Kim, J.S. Comparative Analysis of the YABBY Gene Family of Bienertia sinuspersici, a Single-Cell C4 Plant. Plants 2019, 8, 536. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Li, S.; Gao, Y.; Bartholomew, E.S.; Wang, R.; Yang, H.; Liu, C.; Chen, X.; Wang, Y.; Liu, X.; et al. Genome-Wide Identification of YABBY Gene Family in Cucurbitaceae and Expression Analysis in Cucumber (Cucumis sativus L.). Genes 2022, 13, 467. [Google Scholar] [CrossRef]
- Tanaka, W.; Toriba, T.; Ohmori, Y.; Yoshida, A.; Kawai, A.; Mayama-Tsuchida, T.; Ichikawa, H.; Mitsuda, N.; Ohme-Takagi, M.; Hirano, H.-Y. The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet. Plant Cell 2012, 24, 80–95. [Google Scholar] [CrossRef]
- Zhang, T.; Li, C.; Li, D.; Liu, Y.; Yang, X. Roles of YABBY transcription factors in the modulation of morphogenesis, development, and phytohormone and stress responses in plants. J. Plant Res. 2020, 133, 751–763. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, C.; Ge, D.; Yan, M.; Ren, Y.; Huang, X.; Yuan, Z. Genome-wide identification and expression of YABBY genes family during flower development in Punica granatum L. Gene 2020, 752, 144784. [Google Scholar] [CrossRef]
- Eshed, Y.; Baum, S.F.; Bowman, J.L. Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell 1999, 99, 199–209. [Google Scholar] [CrossRef]
- Villanueva, J.M.; Broadhvest, J.; Hauser, B.A.; Meister, R.J.; Schneitz, K.; Gasser, C.S. INNER NO OUTER regulates abaxial- adaxial patterning in Arabidopsis ovules. Genes Dev. 1999, 13, 3160–3169. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Hsiao, Y.-Y.; Chang, S.-B.; Zhang, D.; Lan, S.-R.; Liu, Z.-J.; Tsai, W.-C. Genome-Wide Identification of YABBY Genes in Orchidaceae and Their Expression Patterns in Phalaenopsis Orchid. Genes 2020, 11, 955. [Google Scholar] [CrossRef]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar] [CrossRef] [PubMed]
- Sawa, S.; Watanabe, K.; Goto, K.; Liu, Y.G.; Shibata, D.; Kanaya, E.; Morita, E.H.; Okada, K. FILAMENTOUS FLOWER, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev. 1999, 13, 1079–1088. [Google Scholar] [CrossRef]
- Bartholmes, C.; Hidalgo, O.; Gleissberg, S. Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biol. 2012, 14, 11–23. [Google Scholar] [CrossRef]
- Hou, H.; Lin, Y.; Hou, X. Ectopic Expression of a Pak-choi YABBY Gene, BcYAB3, Causes Leaf Curvature and Flowering Stage Delay in Arabidopsis thaliana. Genes 2020, 11, 370. [Google Scholar] [CrossRef]
- Bowman, J.L.; Smyth, D.R.; Meyerowitz, E.M. Genes directing flower development in Arabidopsis. Plant Cell 1989, 1, 37–52. [Google Scholar] [CrossRef]
- Lu, Y.-H.; Alam, I.; Yang, Y.-Q.; Yu, Y.-C.; Chi, W.-C.; Chen, S.-B.; Chalhoub, B.; Jiang, L.-X. Evolutionary Analysis of the YABBY Gene Family in Brassicaceae. Plants 2021, 10, 2700. [Google Scholar] [CrossRef]
- Xu, Y.; Qiao, Z.; Zhao, L.; Zhao, W.; Chen, L.; Hu, Z. Genome-wide and Transcriptome Analysis of YABBY Family Gene in Lotus (Nelumbo nucifera Gaertn.). Mol. Plant Breed. 2022. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20210326.1013.006.html (accessed on 14 July 2022).
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.-Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef]
- Marrano, A.; Britton, M.; Zaini, P.A.; Zimin, A.V.; Workman, R.E.; Puiu, D.; Bianco, L.; Pierro, E.A.D.; Allen, B.J.; Chakraborty, S.; et al. High-quality chromosome-scale assembly of the walnut (Juglans regia L.). reference genome. GigaScience 2020, 9, giaa050. [Google Scholar] [CrossRef]
- Yan, F.; Xi, R.-M.; She, R.-X.; Chen, P.-P.; Yan, Y.-J.; Yang, G.; Dang, M.; Yue, M.; Pei, D.; Woeste, K.; et al. Improved de novo chromosome-level genome assembly of the vulnerable walnut tree Juglans mandshurica reveals gene family evolution and possible genome basis of resistance to lesion nematode. Mol. Ecol. Resour. 2021, 21, 2063–2076. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, J.; Liu, H.; Ou, M.; Ye, H.; Zhao, P. Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica. Genes 2022, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.-Q.; Wang, J.; Wong, G.K.-S.; Yu, J. KaKs_Calculator: Calculating Ka and Ks Through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef]
- Feng, S.; Fang, H.; Liu, X.; Dong, Y.; Wang, Q.; Yang, K.Q. Genome-wide identification and characterization of long non-coding RNAs conferring resistance to Colletotrichum gloeosporioides in walnut (Juglans regia). BMC Genom. 2021, 22, 15. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Crepeau, M.W.; Puiu, D.; Gonzalez-Ibeas, D.; Whalen, J.; Stevens, K.A.; Paul, R.; Butterfield, T.S.; Britton, M.T.; Reagan, R.L.; et al. The walnut (Juglans regia) genome sequence reveals diversity in genes coding for the biosynthesis of non-structural polyphenols. Plant J. 2016, 87, 507–532. [Google Scholar] [CrossRef]
- Zhao, S.-P.; Lu, D.; Yu, T.-F.; Ji, Y.-J.; Zheng, W.-J.; Zhang, S.-X.; Chai, S.-C.; Chen, Z.-Y.; Cui, X.-Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef]
- Romanova, M.A.; Maksimova, A.I.; Pawlowski, K.; Voitsekhovskaja, O.V. YABBY Genes in the Development and Evolution of Land Plants. Int. J. Mol. Sci. 2021, 22, 4139. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Van Houten, J.; Gonzalez, G.; Xiao, H.; van der Knaap, E. Genome-wide identification, phylogeny and expression analysis of SUN, OFP and YABBY gene family in tomato. Mol. Genet. Genom. 2013, 288, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Liu, Z.-M.; Li, L.-F.; Leitch, A.R.; Leitch, I.J.; Lohaus, R.; Liu, Z.-J.; Xin, H.-P.; Gong, Y.-B.; Liu, Y.; et al. A genome for gnetophytes and early evolution of seed plants. Nat. Plants 2018, 4, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.M.; Liu, Y.; Rahman, Z.U.; Ali, H.; Yan, C.; Wang, L.; Priyadarshani, S.V.G.N.; Hu, B.; Huang, X.; Xiong, J.; et al. Identification, Characterization and Expression Profiles of Dof Transcription Factors in Pineapple (Ananas comosus L). Trop. Plant Biol. 2018, 11, 49–64. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | No. of Amino Acids | Mol. Wt (Da) | Isoelectric Point (pI) | Instability Index (II) | Aliphatic Index | Grand Average of Hydropathicity (GRAVY) | Subcellular Localization a |
|---|---|---|---|---|---|---|---|
| JrYABBY1 | 192 | 21,332.24 | 8.83 | 37.17 | 72.71 | −0.354 | nucl |
| JrYABBY2 | 211 | 23,466.72 | 7.71 | 49.57 | 77.58 | −0.315 | nucl |
| JrYABBY3 | 213 | 23,467.82 | 8.58 | 50.22 | 77.37 | −0.299 | nucl |
| JrYABBY4 | 177 | 19,751.41 | 8.48 | 48.15 | 68.93 | −0.494 | nucl |
| JrYABBY5 | 215 | 23,918.36 | 7.71 | 47.33 | 79.81 | −0.304 | nucl |
| JrYABBY6 | 187 | 20,819.69 | 8.76 | 37.96 | 76.26 | −0.306 | nucl |
| JrYABBY7 | 168 | 18,543.27 | 9.26 | 41.79 | 72.56 | −0.39 | nucl |
| JrYABBY8 | 186 | 20,892.41 | 5.68 | 71.33 | 71.83 | −0.492 | nucl |
| JrYABBY9 | 207 | 22,630.76 | 8.25 | 48.02 | 75.89 | −0.293 | nucl |
| JrYABBY10 | 181 | 20,172.9 | 7.64 | 52.41 | 65.25 | −0.532 | nucl |
| JmYABBY1 | 202 | 22,227.28 | 8.65 | 35.78 | 76.83 | −0.240 | nucl |
| JmYABBY2 | 182 | 20,381.32 | 8.34 | 40.92 | 79.34 | −0.196 | nucl |
| JmYABBY3 | 186 | 20,887.85 | 5.86 | 59.77 | 93.28 | 0.022 | nucl |
| JmYABBY4 | 122 | 13,404.34 | 6.80 | 42.39 | 82.30 | −0.191 | nucl |
| JmYABBY5 | 211 | 23,466.72 | 7.71 | 49.57 | 77.58 | −0.315 | nucl |
| JmYABBY6 | 215 | 23,887.33 | 8.25 | 45.82 | 82.98 | −0.262 | nucl |
| JmYABBY7 | 207 | 22,700.9 | 8.25 | 48.81 | 78.21 | −0.259 | nucl |
| JmYABBY8 | 138 | 15,184.45 | 6.98 | 44.67 | 74.13 | −0.204 | nucl |
| JmYABBY9 | 155 | 17,011.65 | 8.99 | 45.46 | 80.52 | −0.168 | nucl |
| Gene Name | Whole Genome Duplication (WGD) | Tandem Duplication (TD) | Dispersed Duplication (DSD) | Proximal Duplication (PD) |
|---|---|---|---|---|
| JrYABBY1 | √ | |||
| JrYABBY2 | √ | |||
| JrYABBY3 | √ | |||
| JrYABBY4 | √ | |||
| JrYABBY5 | √ | |||
| JrYABBY6 | √ | |||
| JrYABBY7 | √ | |||
| JrYABBY8 | √ | |||
| JrYABBY9 | √ | |||
| JrYABBY10 | √ | |||
| JmYABBY1 | √ | |||
| JmYABBY2 | √ | |||
| JmYABBY3 | √ | |||
| JmYABBY4 | √ | |||
| JmYABBY5 | √ | |||
| JmYABBY6 | √ | |||
| JmYABBY7 | √ | |||
| JmYABBY8 | √ | |||
| JmYABBY9 | √ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Ye, H.; Wang, J.; Chen, S.; Li, M.; Wang, G.; Hou, N.; Zhao, P. Genome-Wide Identification and Characterization of YABBY Gene Family in Juglans regia and Juglans mandshurica. Agronomy 2022, 12, 1914. https://doi.org/10.3390/agronomy12081914
Liu H, Ye H, Wang J, Chen S, Li M, Wang G, Hou N, Zhao P. Genome-Wide Identification and Characterization of YABBY Gene Family in Juglans regia and Juglans mandshurica. Agronomy. 2022; 12(8):1914. https://doi.org/10.3390/agronomy12081914
Chicago/Turabian StyleLiu, Hengzhao, Hang Ye, Jiangtao Wang, Shenqun Chen, Mengdi Li, Gang Wang, Na Hou, and Peng Zhao. 2022. "Genome-Wide Identification and Characterization of YABBY Gene Family in Juglans regia and Juglans mandshurica" Agronomy 12, no. 8: 1914. https://doi.org/10.3390/agronomy12081914
APA StyleLiu, H., Ye, H., Wang, J., Chen, S., Li, M., Wang, G., Hou, N., & Zhao, P. (2022). Genome-Wide Identification and Characterization of YABBY Gene Family in Juglans regia and Juglans mandshurica. Agronomy, 12(8), 1914. https://doi.org/10.3390/agronomy12081914