
Expression Pattern, Molecular Docking and Dynamics Simulation Analysis of CSP4 from Sirex nitobei Provides Molecular Basis of CSP Bound to Scent Molecules



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Total RNA Solation and First-Strand cDNA Synthesis
2.3. PCR Amplification and Sequence Analysis
2.4. Tissue Expression Profile by qPCR
2.5. Homologous Modeling and Models Evaluation
2.6. Molecular Docking
2.7. Molecular Dynamics
3. Results
3.1. Sequence Analysis of SnitCSP4
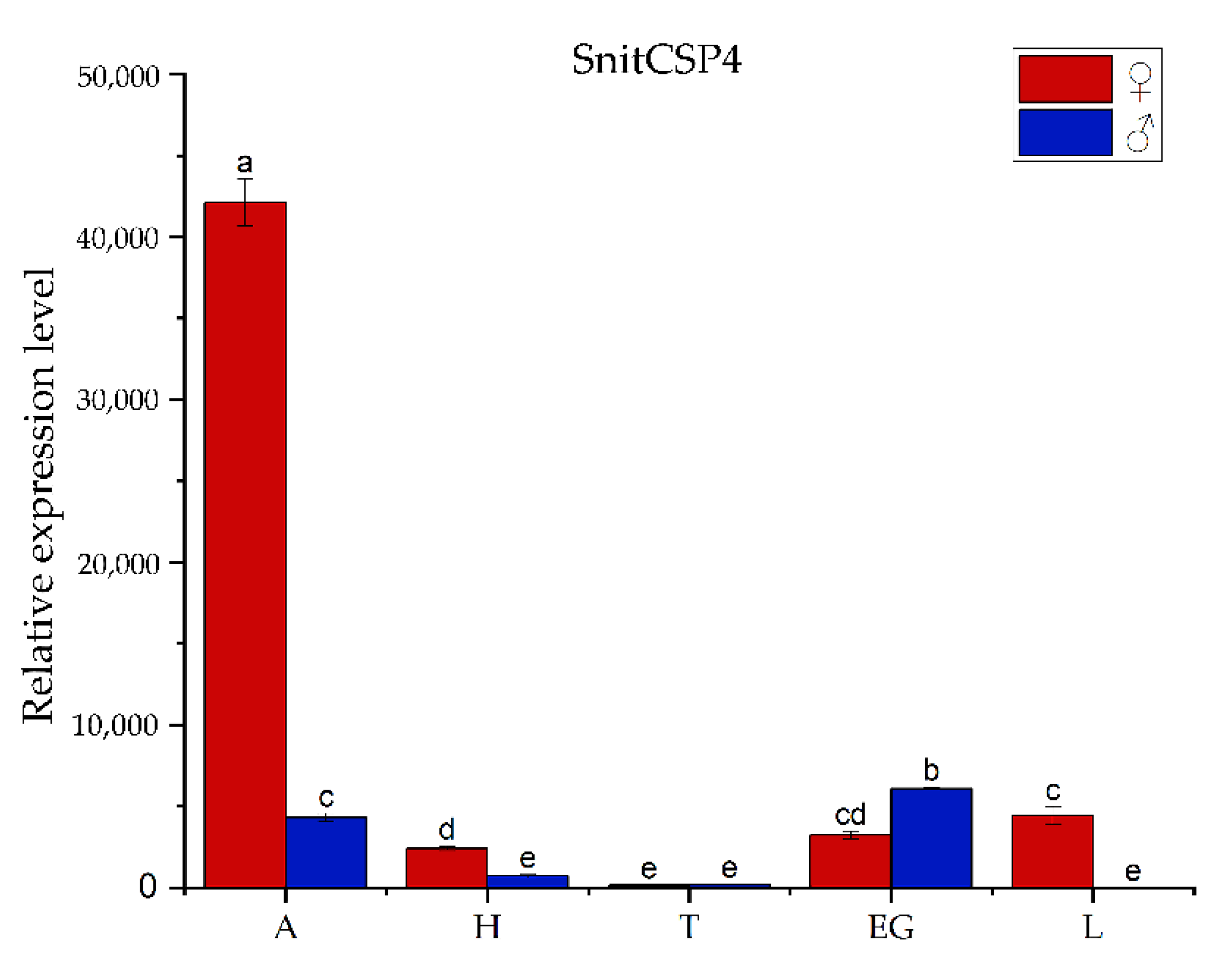
3.2. Tissues-Specificity Expression Analysis
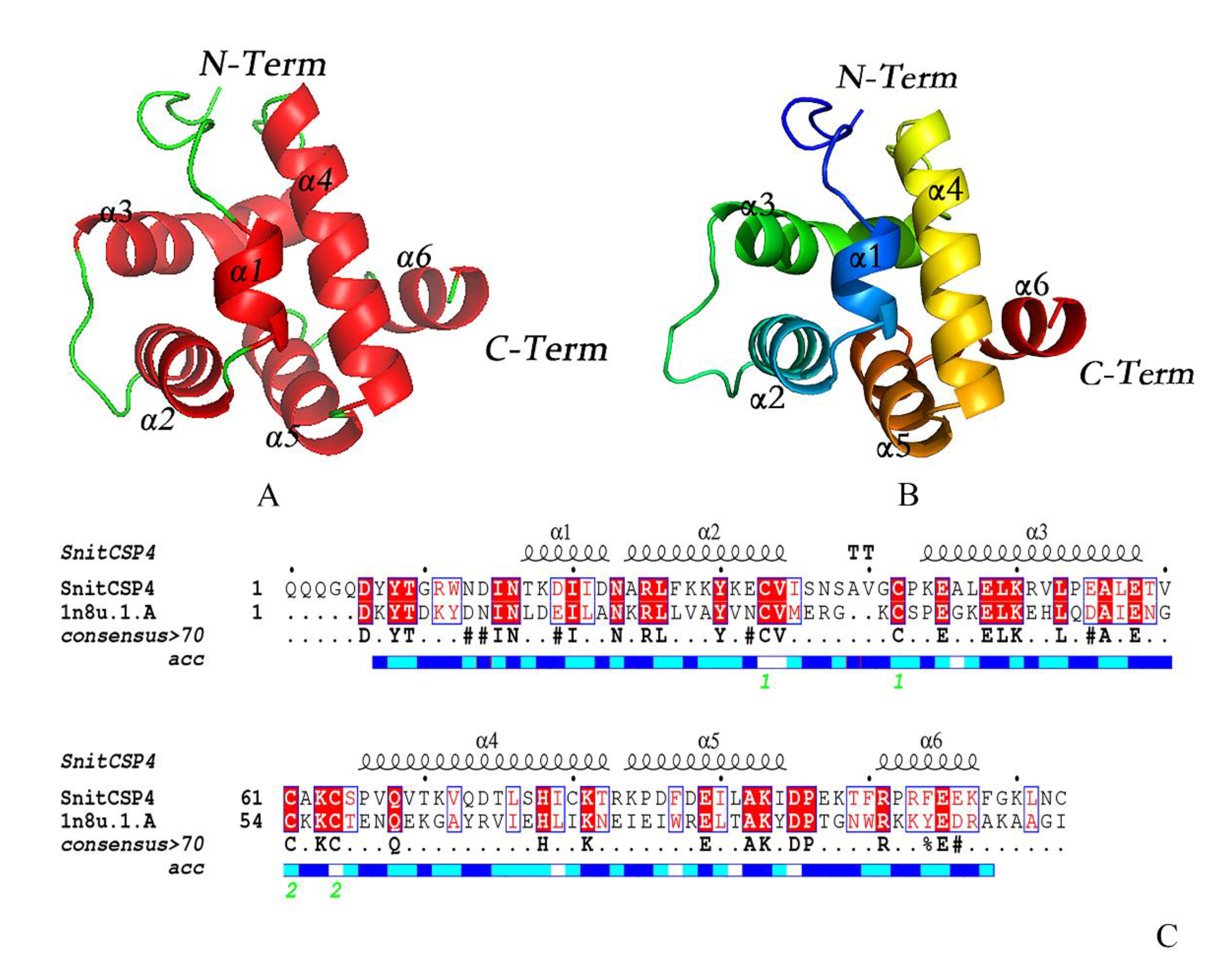
3.3. Homology Modeling Analysis
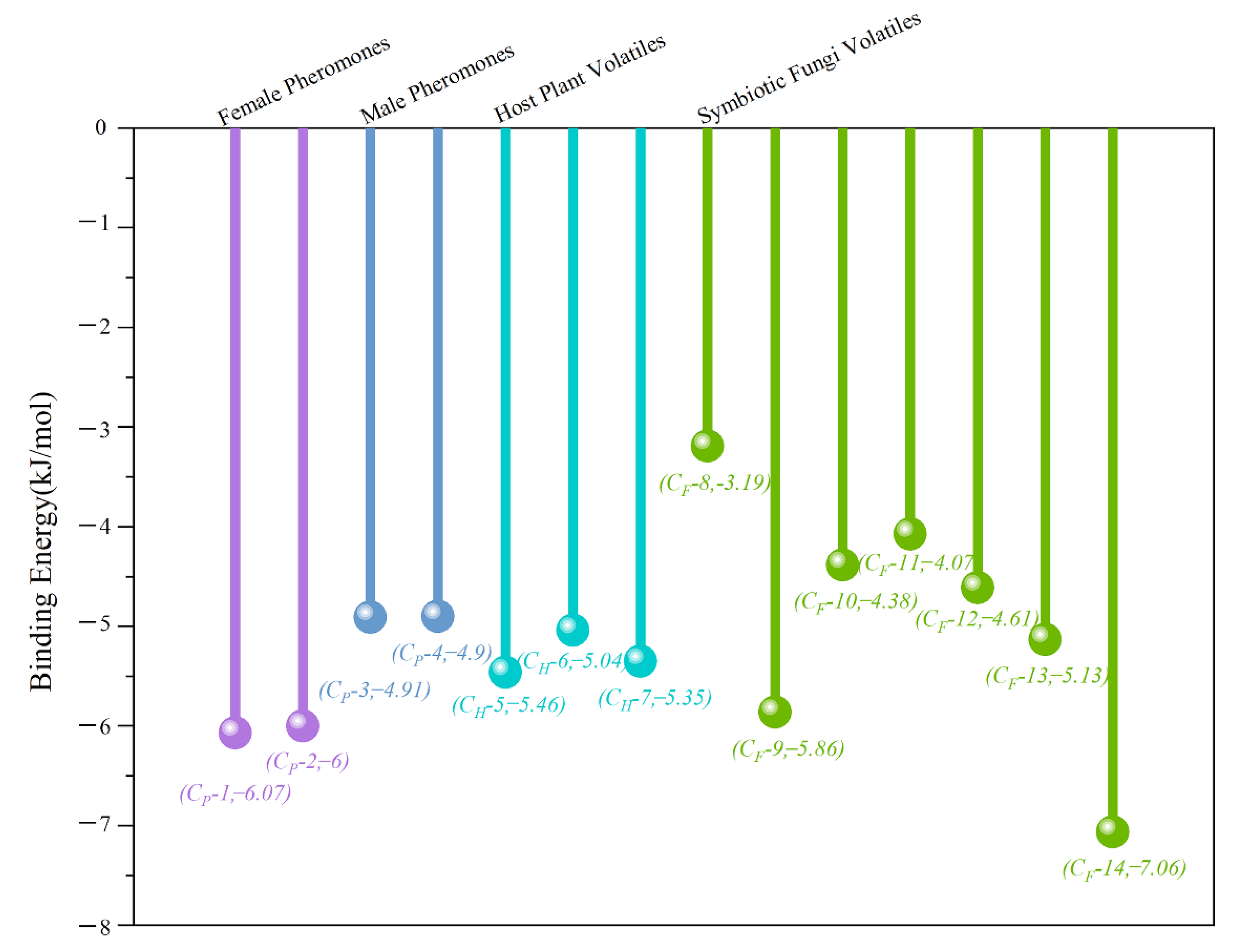
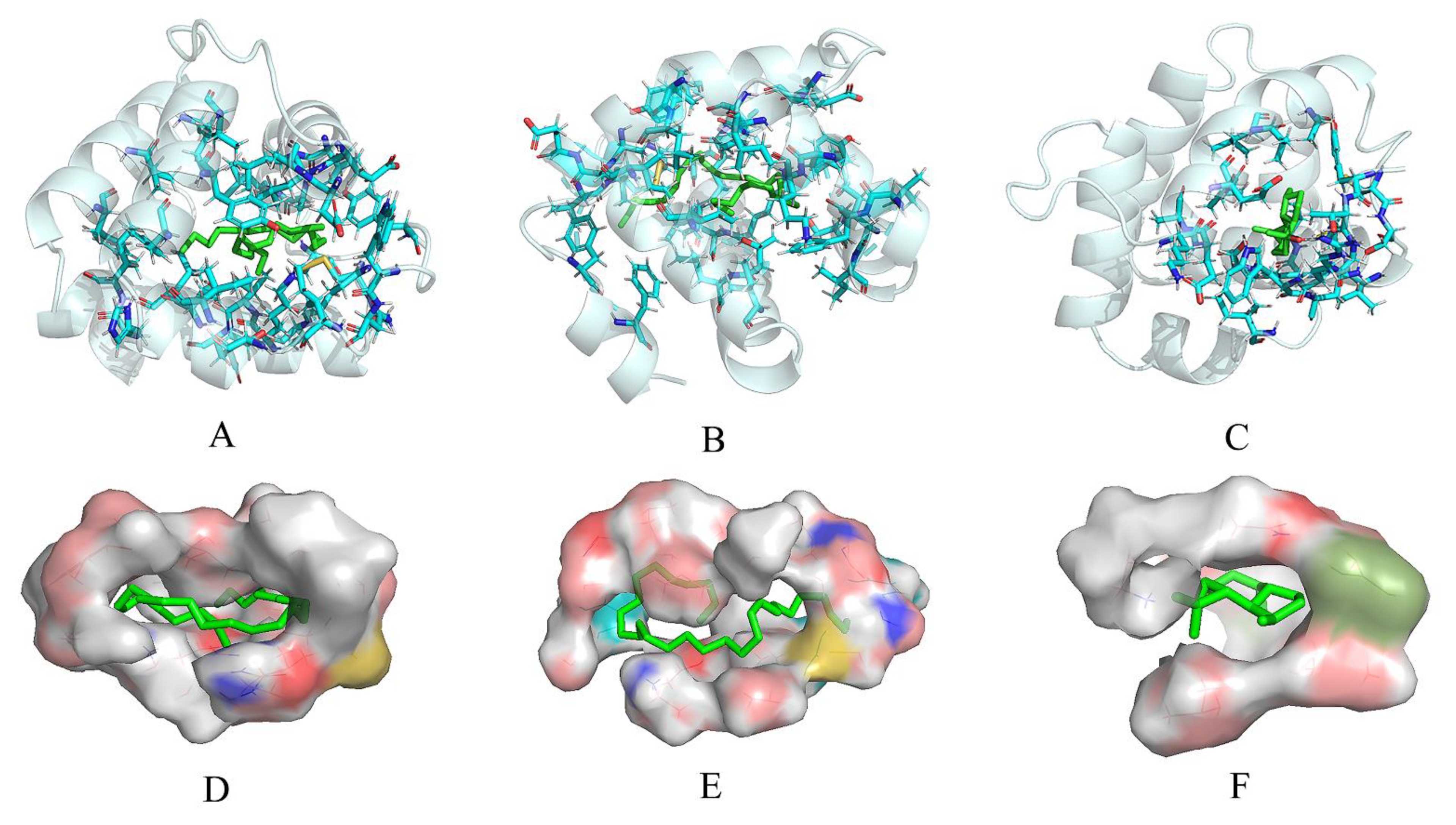
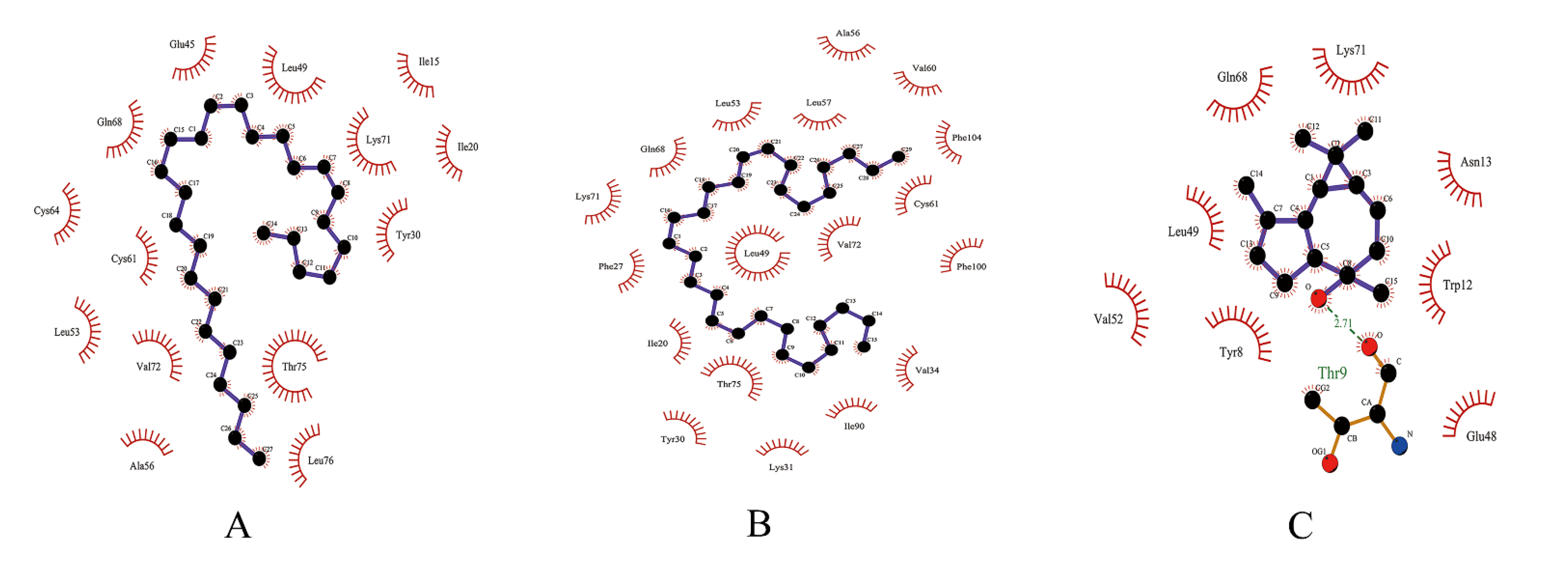
3.4. Molecular Docking Analysis
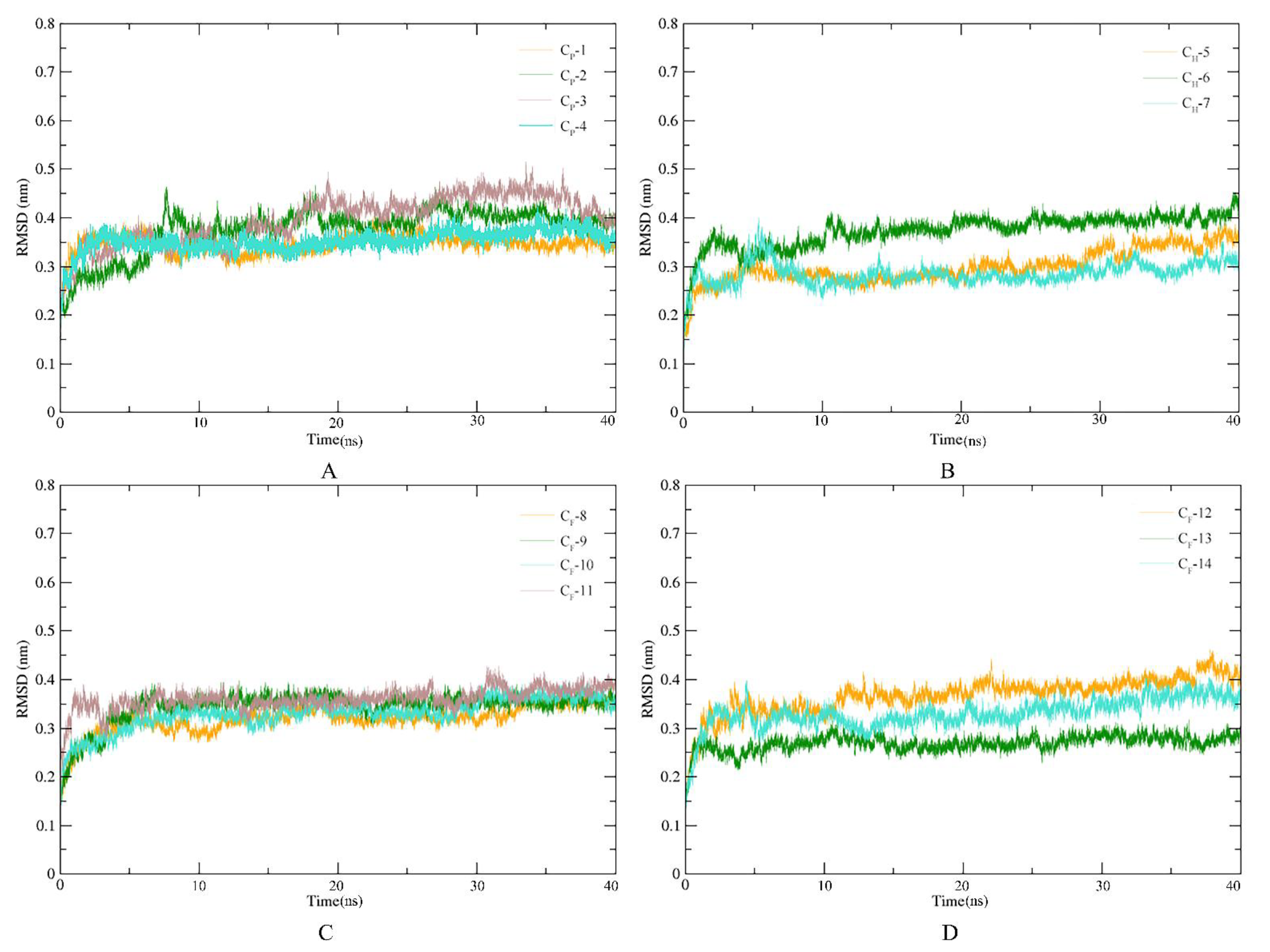
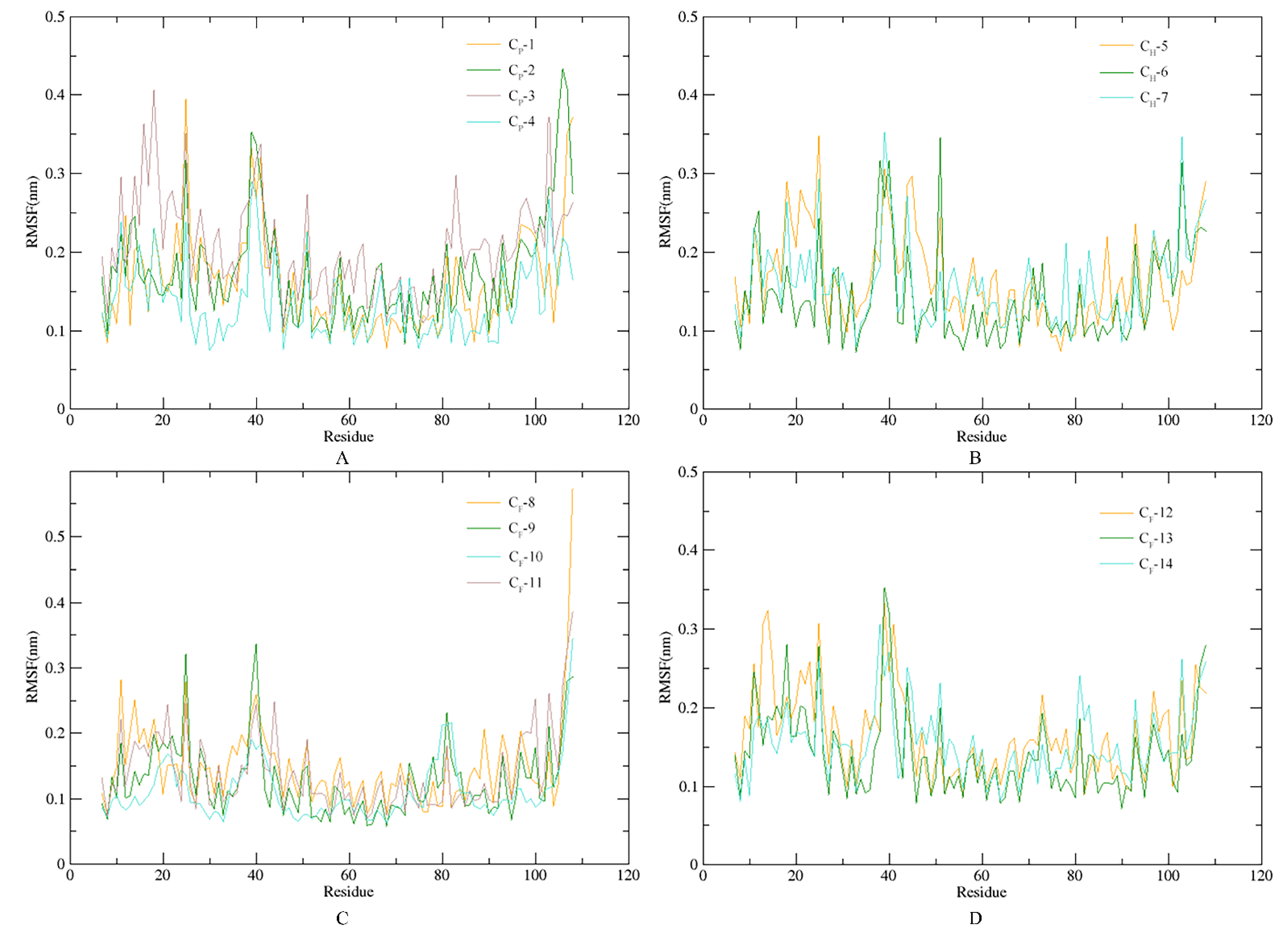
3.5. Stability of SnitCSP4-Ligand Complexes in MD Simulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble Proteins in Insect Chemical Communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- van der Goes van Naters, W.; Carlson, J.R. Insects as Chemosensors of Humans and Crops. Nature 2006, 444, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the Odor World: Olfactory Receptors and Their Role for Signal Transduction in Insects. Cell. Mol. Life Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Pheromone Reception. In The Chemistry of Pheromones and Other Semiochemicals II; Schulz, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 1–36. ISBN 978-3-540-31477-6. [Google Scholar]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Antony, B.; Soffan, A.; Jakše, J.; Abdelazim, M.M.; Aldosari, S.A.; Aldawood, A.S.; Pain, A. Identification of the Genes Involved in Odorant Reception and Detection in the Palm Weevil Rhynchophorus Ferrugineus, an Important Quarantine Pest, by Antennal Transcriptome Analysis. BMC Genom. 2016, 17, 69. [Google Scholar] [CrossRef]
- Pelosi, P. Diversity of Odorant-Binding Proteins and Chemosensory Proteins in Insects. Chem. Senses 2005, 30, i291–i292. [Google Scholar] [CrossRef] [PubMed]
- Angeli, S.; Ceron, F.; Scaloni, A.; Monti, M.; Monteforti, G.; Minnocci, A.; Petacchi, R.; Pelosi, P. Purification, Structural Characterization, Cloning and Immunocytochemical Localization of Chemoreception Proteins from Schistocerca Gregaria. Eur. J. Biochem. 1999, 262, 745–754. [Google Scholar] [CrossRef]
- Jin, R.; Liu, N.; Liu, Y.; Dong, S. A Larval Specific OBP Able to Bind the Major Female Sex Pheromone Component in Spodoptera Exigua (Hübner). J. Integr. Agric. 2015, 14, 1356–1366. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, W.; Zhang, Z.; Zhang, Y.; Guo, Y. Functional Characteristics of a Novel Chemosensory Protein in the Cotton Bollworm Helicoverpa Armigera (Hübner). J. Integr. Agric. 2013, 12, 853–861. [Google Scholar] [CrossRef]
- Li; Gu, T.; Chen, C.; Huang, K.; Tian, S.; Zhao, X.; Hao, D. CDNA Cloning, Sequence Analysis and Expression Profie of a Chemosensory Protoin from the Clostera Restitura (Lepidoptera:Notodontidae). Sci. Silvae Sin. 2018, 54, 67–75. [Google Scholar] [CrossRef]
- Nomura Kitabayashi, A.; Arai, T.; Kubo, T.; Natori, S. Molecular Cloning of CDNA for P10, a Novel Protein That Increases in the Regenerating Legs of Periplaneta Americana (American Cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- He, P.; Li, Z.-Q.; Zhang, Y.-F.; Chen, L.; Wang, J.; Xu, L.; Zhang, Y.-N.; He, M. Identification of Odorant-Binding and Chemosensory Protein Genes and the Ligand Affinity of Two of the Encoded Proteins Suggest a Complex Olfactory Perception System in Periplaneta Americana: Cockroach Odorant Binding and Chemosensory Protein. Insect Mol. Biol. 2017, 26, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-N.; Ye, Z.-F.; Yang, K.; Dong, S.-L. Antenna-Predominant and Male-Biased CSP19 of Sesamia Inferens Is Able to Bind the Female Sex Pheromones and Host Plant Volatiles. Gene 2014, 536, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.-H.; Wang, S.-Y.; Zhang, X.-Y.; Ji, P.; Liu, J.-T.; Wang, G.-R.; Wu, K.-M.; Guo, Y.-Y.; Zhou, J.-J.; Zhang, Y.-J. Functional Characterizations of Chemosensory Proteins of the Alfalfa Plant Bug Adelphocoris Lineolatus Indicate Their Involvement in Host Recognition. PLoS ONE 2012, 7, e42871. [Google Scholar] [CrossRef]
- Iovinella, I.; Bozza, F.; Caputo, B.; Della Torre, A.; Pelosi, P. Ligand-Binding Study of Anopheles Gambiae Chemosensory Proteins. Chem. Senses 2013, 38, 409–419. [Google Scholar] [CrossRef]
- Dyanov, H.M.; Dzitoeva, S.G. Method for Attachment of Microscopic Preparations on Glass for in Situ Hybridization, PRINS and in Situ PCR Studies. Biotechniques 1995, 18, 822–824, 826. [Google Scholar]
- Singh, S.; Tyagi, C.; Rather, I.A.; Sabir, J.S.M.; Hassan, M.I.; Singh, A.; Singh, I.K. Molecular Modeling of Chemosensory Protein 3 from Spodoptera Litura and Its Binding Property with Plant Defensive Metabolites. IJMS 2020, 21, 4073. [Google Scholar] [CrossRef]
- Waris, M.I.; Younas, A.; Ameen, A.; Rasool, F.; Wang, M.-Q. Expression Profiles and Biochemical Analysis of Chemosensory Protein 3 from Nilaparvata Lugens (Hemiptera: Delphacidae). J. Chem. Ecol. 2020, 46, 363–377. [Google Scholar] [CrossRef]
- Zhou, X.-H.; Ban, L.-P.; Iovinella, I.; Zhao, L.-J.; Gao, Q.; Felicioli, A.; Sagona, S.; Pieraccini, G.; Pelosi, P.; Zhang, L.; et al. Diversity, Abundance, and Sex-Specific Expression of Chemosensory Proteins in the Reproductive Organs of the Locust Locusta Migratoria Manilensis. Biol. Chem. 2013, 394, 43–54. [Google Scholar] [CrossRef]
- Maleszka, J.; Forêt, S.; Saint, R.B.; Maleszka, R. RNAi-Induced Phenotypes Suggest a Novel Role for a Chemosensory Protein CSP5 in the Development of Embryonic Integument in the Honeybee (Apis Mellifera). Dev. Genes Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef]
- Nomura, A.; Kawasaki, K.; Kubo, T.; Natori, S. Purification and Localization of P10, a Novel Protein That Increases in Nymphal Regenerating Legs of Periplaneta Americana (American Cockroach). Int. J. Dev. Biol. 1992, 36, 391–398. [Google Scholar] [PubMed]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory Proteins in the Honey Bee: Insights from the Annotated Genome, Comparative Analyses and Expressional Profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Iovinella, I.; Dani, F.R.; Liu, Y.-L.; Huang, L.-Q.; Liu, Y.; Wang, C.-Z.; Pelosi, P.; Wang, G. Conserved Chemosensory Proteins in the Proboscis and Eyes of Lepidoptera. Int. J. Biol. Sci. 2016, 12, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Dani, F.R.; Michelucci, E.; Francese, S.; Mastrobuoni, G.; Cappellozza, S.; La Marca, G.; Niccolini, A.; Felicioli, A.; Moneti, G.; Pelosi, P. Odorant-Binding Proteins and Chemosensory Proteins in Pheromone Detection and Release in the Silkmoth Bombyx Mori. Chem. Senses 2011, 36, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Iovinella, I.; Dani, F.R.; Niccolini, A.; Sagona, S.; Michelucci, E.; Gazzano, A.; Turillazzi, S.; Felicioli, A.; Pelosi, P. Differential Expression of Odorant-Binding Proteins in the Mandibular Glands of the Honey Bee According to Caste and Age. J. Proteome Res. 2011, 10, 3439–3449. [Google Scholar] [CrossRef]
- Jacquin-Joly, E. Functional and Expression Pattern Analysis of Chemosensory Proteins Expressed in Antennae and Pheromonal Gland of Mamestra Brassicae. Chem. Senses 2001, 26, 833–844. [Google Scholar] [CrossRef]
- Schiff, N.; Goulet, H.; Smith, D.; Boudreault, C.; Wilson, A.; Scheffler, B.E. Siricidae (Hymenoptera: Symphyta: Siricoidea) of the Western Hemisphere. Can. J. Arthropod Identif. 2012, 21, 1–305. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Fu, N.; Gao, C.; Ao, T.; Ren, L.; Luo, Y. Comparison of Wing, Ovipositor, and Cornus Morphologies between Sirex Noctilio and Sirex Nitobei Using Geometric Morphometrics. Insects 2020, 11, 84. [Google Scholar] [CrossRef]
- Fukuda, H.; Hijii, N. Different Parasitism Patterns of Two Hymenopterous Parasitoids (Ichineumonidae and Ibaliidae) Depending on the Development of Sirex Nitobei (Hym., Siricidae). J. Appl. Entomol. 1996, 120, 301–305. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sasaki, K.; Enda, N. Correlation between Sirex Nitobei and Amylostereum Areolatum, Associated with the Death of Japanese Pine Trees during Winter Season. J. Jpn. For. Soc. 1978, 60, 405–411. [Google Scholar]
- Fitza, K.N.E.; Tabata, M.; Kanzaki, N.; Kimura, K.; Garnas, J.; Slippers, B. Host Specificity and Diversity of Amylostereum Associated with Japanese Siricids. Fungal Ecol. 2016, 24, 76–81. [Google Scholar] [CrossRef]
- Xiao, G.R.; Huang, X.Y.; Zhou, S.Z.; Wu, J.; Zhang, P. Economic Sawfly Fauna of China; Tianze Eldonejo: Beijing, China, 1991; pp. 37–42. [Google Scholar]
- Guo, B.; Hao, E.; Qiao, H.; Wang, J.; Wu, W.; Zhou, J.; Lu, P. Antennal Transcriptome Analysis of Olfactory Genes and Characterizations of Odorant Binding Proteins in Two Woodwasps, Sirex Noctilio and Sirex Nitobei (Hymenoptera: Siricidae). BMC Genom. 2021, 22, 172. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein Structure Homology Modeling Using SWISS-MODEL Workspace. Nat. Protoc. 2009, 4, 1–13. [Google Scholar] [CrossRef]
- Luthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of Protein Models with Three-Dimensional Profiles. Nature 1992, 356, 83–85. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of Protein Structures: Patterns of Nonbonded Atomic Interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Moss, D.S.; Thornton, J.M. Main-Chain Bond Lengths and Bond Angles in Protein Structures. J. Mol. Biol. 1993, 231, 1049–1067. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Ren, J.; Yuan, X.; Li, J.; Lin, S.; Yang, B.; Chen, C.; Zhao, J.; Zheng, W.; Liao, H.; Yang, Z.; et al. Assessing the Performance of the G_mmpbsa Tools to Simulate the Inhibition of Oseltamivir to Influenza Virus Neuraminidase by Molecular Mechanics Poisson–Boltzmann Surface Area Methods. J. Chin. Chem. Soc. 2020, 67, 46–53. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Sousa da Silva, A.W.; Vranken, W.F. ACPYPE-AnteChamber PYthon Parser InterfacE. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.; Jaiswal, N.; Singh, S.; Pandey, V.P.; Dwivedi, U.N. Molecular Docking and Dynamics Simulation Analyses Unraveling the Differential Enzymatic Catalysis by Plant and Fungal Laccases with Respect to Lignin Biosynthesis and Degradation. J. Biomol. Struct. Dyn. 2015, 33, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Calvello, M.; Guerra, N.; Brandazza, A.; D’Ambrosio, C.; Scaloni, A.; Dani, F.R.; Turillazzi, S.; Pelosi, P. Soluble Proteins of Chemical Communication in the Social Wasp Polistes Dominulus. Cell. Mol. Life Sci. (CMLS) 2003, 60, 1933–1943. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Chiang, V.; Leal, W.S. Protein That Makes Sense in the Argentine Ant. Naturwissenschaften 2002, 89, 505–507. [Google Scholar] [CrossRef]
- González, D.; Zhao, Q.; McMahan, C.; Velasquez, D.; Haskins, W.E.; Sponsel, V.; Cassill, A.; Renthal, R. The Major Antennal Chemosensory Protein of Red Imported Fire Ant Workers. Insect Mol. Biol. 2009, 18, 395–404. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, F.; Zhang, X.; Zhang, S.; Guo, S.; Zhu, G.; Liu, Q.; Li, M. Transcriptome and Expression Patterns of Chemosensory Genes in Antennae of the Parasitoid Wasp Chouioia Cunea. PLoS ONE 2016, 11, e0148159. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, S.-N.; Li, K.-M.; Liu, J.-T.; Zheng, Y.; Shan, S.; Yang, Y.-Q.; Li, R.-J.; Zhang, Y.-J.; Guo, Y.-Y. Identification of Odorant Binding Proteins and Chemosensory Proteins in Microplitis Mediator as Well as Functional Characterization of Chemosensory Protein 3. PLoS ONE 2017, 12, e0180775. [Google Scholar] [CrossRef]
- Tomaselli, S.; Crescenzi, O.; Sanfelice, D.; Ab, E.; Wechselberger, R.; Angeli, S.; Scaloni, A.; Boelens, R.; Tancredi, T.; Pelosi, P.; et al. Solution Structure of a Chemosensory Protein from the Desert Locust Schistocerca Gregaria. Biochemistry 2006, 45, 10606–10613. [Google Scholar] [CrossRef]
- Campanacci, V.; Lartigue, A.; Hällberg, B.M.; Jones, T.A.; Giudici-Orticoni, M.-T.; Tegoni, M.; Cambillau, C. Moth Chemosensory Protein Exhibits Drastic Conformational Changes and Cooperativity on Ligand Binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5069–5074. [Google Scholar] [CrossRef]
- Li, Y.-N.; Hao, E.-H.; Li, H.; Yuan, X.-H.; Lu, P.-F.; Qiao, H.-L. Computational Interaction Analysis of Sirex Noctilio Odorant-Binding Protein (SnocOBP7) Combined with Female Sex Pheromones and Symbiotic Fungal Volatiles. Agronomy 2021, 11, 2461. [Google Scholar] [CrossRef]
- Fan, J. cDNA Isolation and Protein Expression of OBP3 in Six Insects and Identification of Proteins Associated with Olfaction in Sitobion Avenae; Chinese Academy of Agricultural Sciences: Beijing, China, 2011. [Google Scholar]
- Tian, Z.; Liu, J.; Zhang, Y. Structural Insights into Cydia Pomonella Pheromone Binding Protein 2 Mediated Prediction of Potentially Active Semiochemicals. Sci. Rep. 2016, 6, 22336. [Google Scholar] [CrossRef] [PubMed]
- Venthur, H.; Machuca, J.; Godoy, R.; Palma-Millanao, R.; Zhou, J.; Larama, G.; Bardehle, L.; Quiroz, A.; Ceballos, R.; Mutis, A. Structural Investigation of Selective Binding Dynamics for the Pheromone-binding Protein 1 of the Grapevine Moth, Lobesia Botrana. Arch. Insect Biochem. Physiol 2019, 101, e21557. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Jiao, J.; Wang, Q. Brief report on luring and collecting Sirex nitobei by trap log. Heilongjiang Agric. Sci. 2011, 5, 57–59. [Google Scholar]
- Bao, M.; Qiao, H.; Shi, J.; Luo, Y.; Lu, P. Research progress in reproductive behavior and chemical ecological regulation of the european woodwasp (Sirex noctilio), a severe invasive pest. Sci. Silvae Sin. 2020, 56, 127–141. [Google Scholar] [CrossRef]
- Xu, Q.; Sun, X.-T.; Lu, P.-F.; Luo, Y.-Q.; Shi, J. Volatile Profiles of Three Tree Species in the Northeastern China and Associated Effects on Sirex Noctilio Activity. J. Plant Interact. 2019, 14, 334–339. [Google Scholar] [CrossRef]
- Wang, M.; Bao, M.; Ao, T.; Ren, L.; Luo, Y. Population Distribution Patterns and Ecological Niches of Two Sirex Species Damaging Pinus Sylvestris Var. Mongolica. Chin. J. Appl. Entomol. 2017, 54, 924–932. [Google Scholar]
- Boroczky, K.; Crook, D.J.; Francese, J.A.; Mastro, V.C.; Tumlinson, J.H. Chemical Ecology of Sirex Noctilio. In Proceedings 19th U.S. Department of Agriculture Interagency Research Forum on Invasive Species 2008; McManus, K.A., Gottschalk, K.W., Eds.; 2009. [Google Scholar]
- Bashford, R. The Development of Static Trapping Systems to Monitor for Wood-Boring Insects in Forestry Plantations. Aust. For. 2008, 71, 236–241. [Google Scholar] [CrossRef]
- Li, D.; Shi, J.; Lu, M.; Ren, L.; Zhen, C.; Luo, Y. Detection and Identification of the Invasive Sirex Noctilio (Hymenoptera: Siricidae) Fungal Symbiont, Amylostereum Areolatum (Russulales: Amylostereacea), in China and the Stimulating Effect of Insect Venom on Laccase Production by A. Areolatum YQL03. J. Econ. Entomol. 2015, 108, 1136–1147. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.; Zong, S.; Tao, J. Molecular Characterization and Expression Profiling of Chemosensory Proteins in Male Eogystia Hippophaecolus (Lepidoptera: Cossidae). J. Entomol. Sci. 2021, 56, 217–234. [Google Scholar] [CrossRef]
- Nagnan-Le Meillour, P. Chemosensory Proteins from the Proboscis of Mamestra Brassicae. Chem. Senses 2000, 25, 541–553. [Google Scholar] [CrossRef]
- Thompson, B.M.; Grebenok, R.J.; Behmer, S.T.; Gruner, D.S. Microbial Symbionts Shape the Sterol Profile of the Xylem-Feeding Woodwasp, Sirex Noctilio. J. Chem. Ecol. 2013, 39, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.M.; Bodart, J.; McEwen, C.; Gruner, D.S. Adaptations for Symbiont-Mediated External Digestion in Sirex Noctilio (Hymenoptera: Siricidae). Ann. Entomol. Soc. Am. 2014, 107, 453–460. [Google Scholar] [CrossRef]
- Buchanan, P.K. Aphyllophorales in Australasia. Aust. Systematic Bot. 2001, 14, 417. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, T.-F.; Qi, L.; Liu, F.; Zhu, Y.-J.; Hou, C.; Yu, L.-S. Spatiotemporal expression levels of three chemosensory proteins in adult Apis mellifera ligustica workers. Chin. J. Appl. Entomol. 2018, 55, 71–78. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primers Sequence (5′→3′) | Annealing Temperature, °C | Segment Length, bp |
|---|---|---|---|
| SnitCSP4 | F: ATTTCGAACAGCGCAGTTGG R: TCATCAAAGTCGGGCTTTCG | 59.0 | 164 |
| β-Tubulin | F: CGTCGGTTCCGTTGATAAGTTG R: AGAATATCCCGACCGAGTGTTG | 59.0 | 122 |
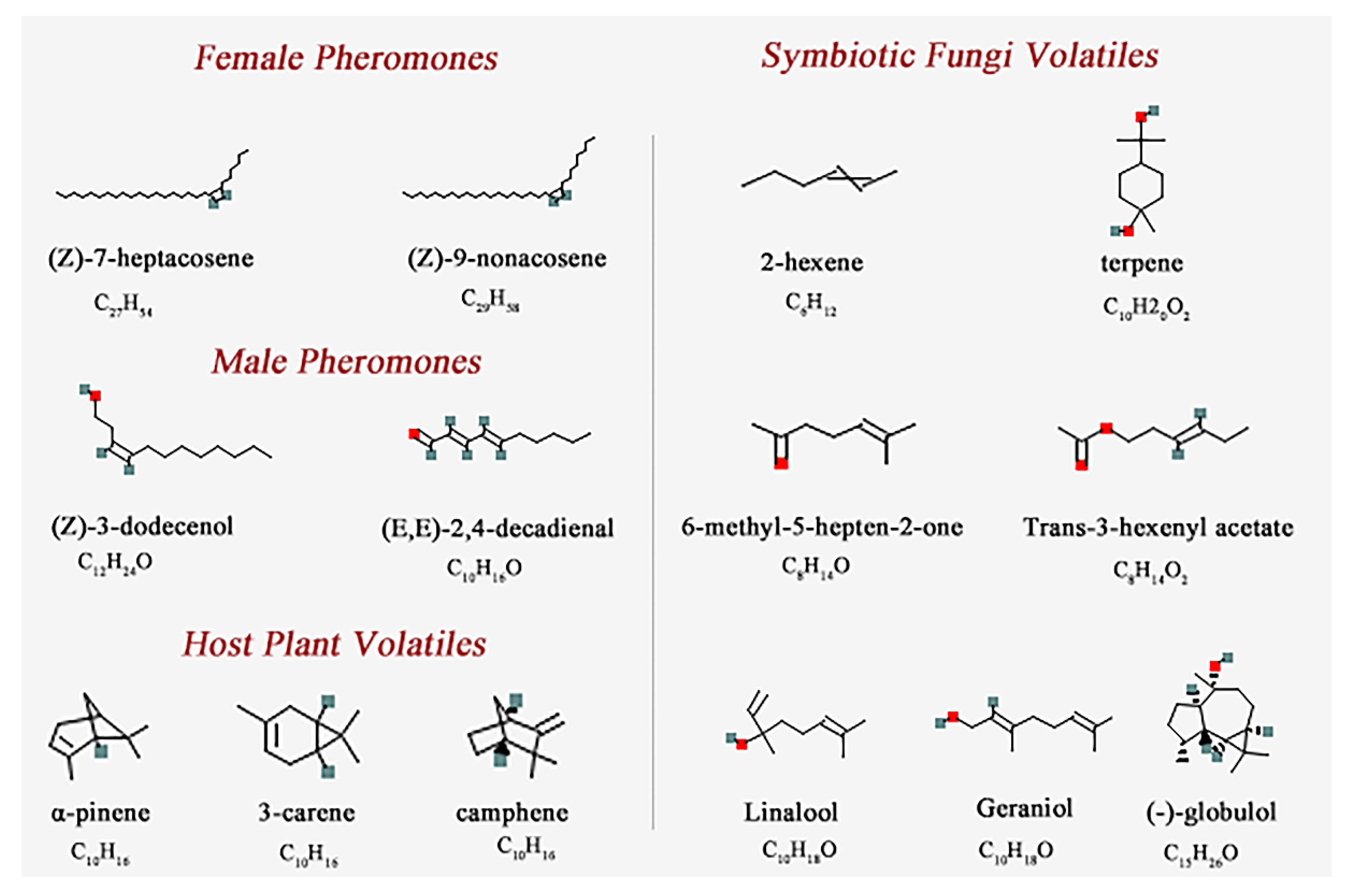
| Chemical Name | PubChem CID No. | Molecular Formula | Complex Code | Chemical Name | PubChem CID No. | Molecular Formula | Complex Code |
|---|---|---|---|---|---|---|---|
| Female Pheromones | Male Pheromones | ||||||
| (Z)-7-heptacosene | 56936088 | C27H54 | CP-1 | (Z)-3-dodecenol | 5364626 | C12H24O | CP-3 |
| (Z)-9-nonacosene | 14367299 | C29H58 | CP-2 | (E,E)-2,4-decadienal | 5283349 | C10H16O | CP-4 |
| Host Plant Volatiles | Symbiotic Fungi Volatiles | ||||||
| α-pinene | 6654 | C10H16 | CH-5 | 2-hexene | 19966 | C6H12 | CF-8 |
| 3-carene | 26049 | C10H16 | CH-6 | terpene | 6651 | C10H20O2 | CF-9 |
| camphene | 92221 | C10H16 | CH-7 | 6-methyl-5-hepten-2-one | 9862 | C8H14O | CF-10 |
| Trans-3-hexenyl acetate | 5352557 | C8H14O2 | CF-11 | ||||
| Linalool | 6549 | C10H18O | CF-12 | ||||
| Geraniol | 637566 | C10H18O | CF-13 | ||||
| (-)-globulol | 12304985 | C15H26O | CF-14 |
| Name | Molecular Formula | MW (ku) | pI | Aliphatic Index | Instability Index | GRAVY |
|---|---|---|---|---|---|---|
| SnitCSP4 | C675H1079N179O193S9 | 15.08 | 8.73 | 82.60 | 38.08 | −0.330 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Hao, E.; Li, H.; Yang, X.; Lu, P.; Qiao, H. Expression Pattern, Molecular Docking and Dynamics Simulation Analysis of CSP4 from Sirex nitobei Provides Molecular Basis of CSP Bound to Scent Molecules. Agronomy 2022, 12, 1994. https://doi.org/10.3390/agronomy12091994
Guo P, Hao E, Li H, Yang X, Lu P, Qiao H. Expression Pattern, Molecular Docking and Dynamics Simulation Analysis of CSP4 from Sirex nitobei Provides Molecular Basis of CSP Bound to Scent Molecules. Agronomy. 2022; 12(9):1994. https://doi.org/10.3390/agronomy12091994
Chicago/Turabian StyleGuo, Pingping, Enhua Hao, Han Li, Xi Yang, Pengfei Lu, and Haili Qiao. 2022. "Expression Pattern, Molecular Docking and Dynamics Simulation Analysis of CSP4 from Sirex nitobei Provides Molecular Basis of CSP Bound to Scent Molecules" Agronomy 12, no. 9: 1994. https://doi.org/10.3390/agronomy12091994
APA StyleGuo, P., Hao, E., Li, H., Yang, X., Lu, P., & Qiao, H. (2022). Expression Pattern, Molecular Docking and Dynamics Simulation Analysis of CSP4 from Sirex nitobei Provides Molecular Basis of CSP Bound to Scent Molecules. Agronomy, 12(9), 1994. https://doi.org/10.3390/agronomy12091994