
Soil Organic Nitrogen Indirectly Enhances Pepper-Residue-Mediated Soil Disease Suppression through Manipulation of Soil Microbiome
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crop Residue Preparation and Design of Greenhouse Pot Trials
2.2. Disease Incidence Determination
2.3. Banana Biomass Determination
2.4. Collection of Soil Samples
2.5. Determination of Soil Chemical Properties
2.6. Determination of Soil Culturable Microbes
2.7. Preparation of Plant Residual Extracts
2.8. Determination of the Inhibitory Effect of Plant Residue Extracts on the Growth of Pathogens
2.9. Statistical Analyses
3. Results
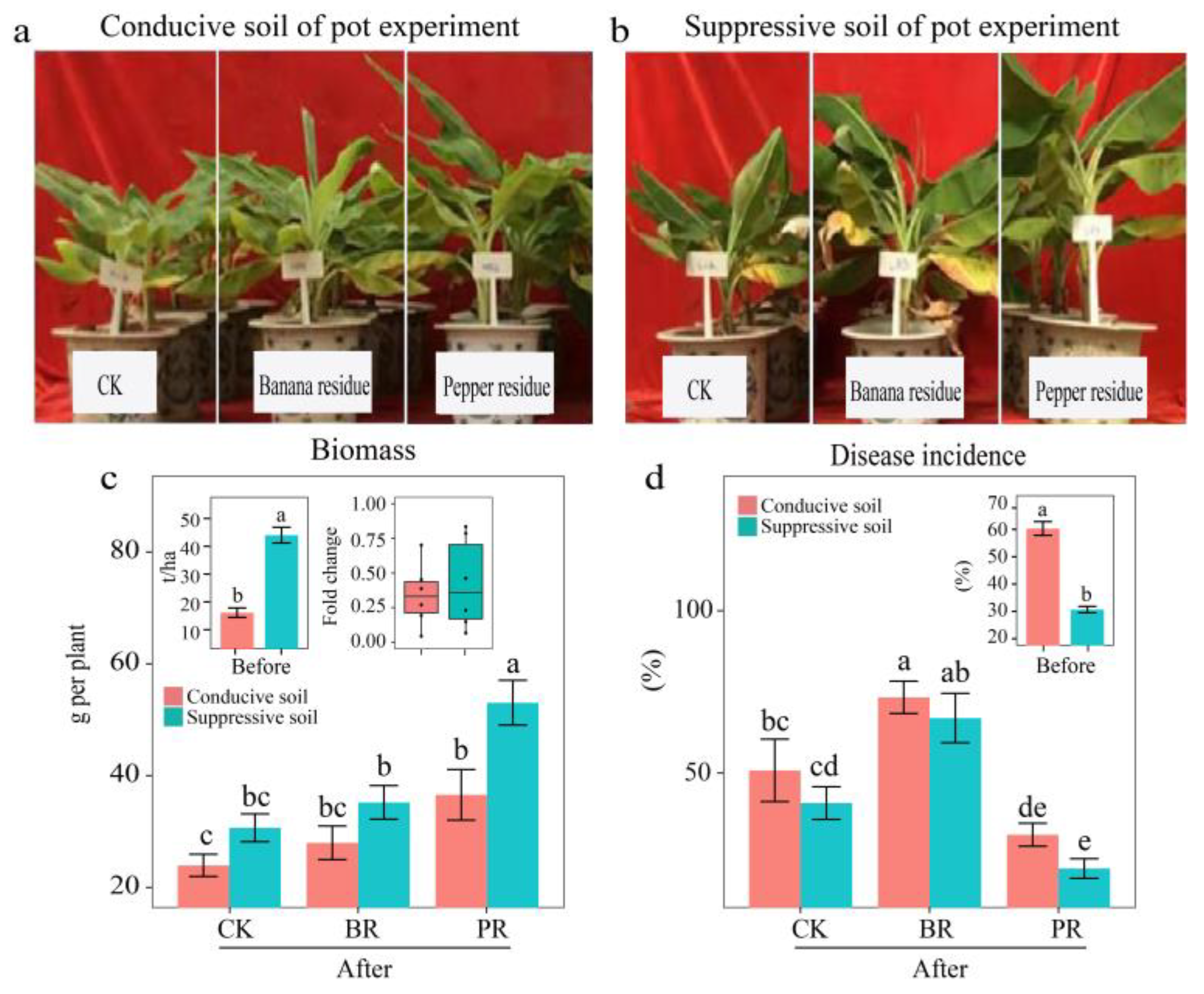
3.1. Changes in the Biomass and Disease Incidence of Banana after Adding Residues to Conducive and Suppressive Soil
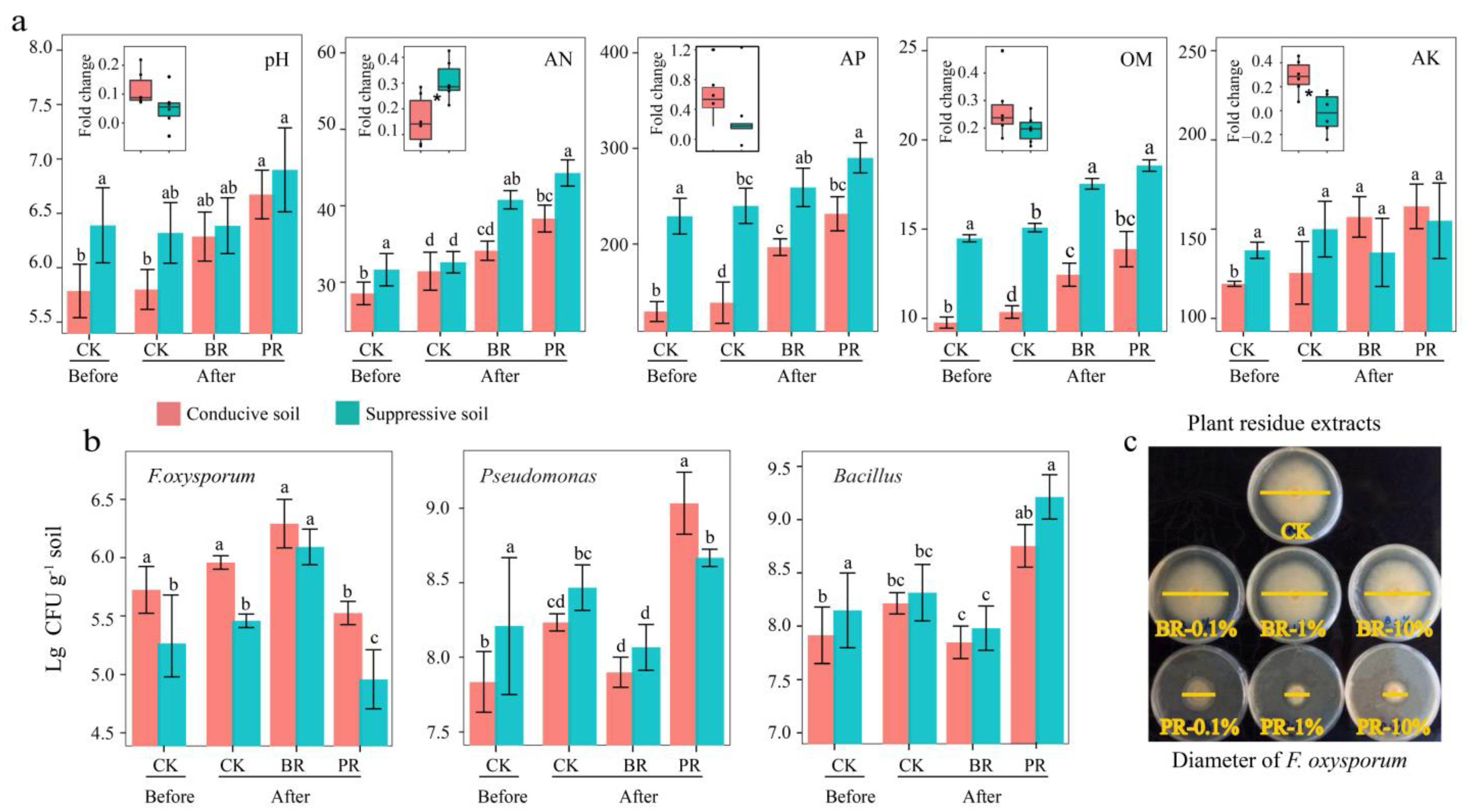
3.2. Soil Physicochemical, Culturable Microbes and Disease Suppression Effect of Residue Extract
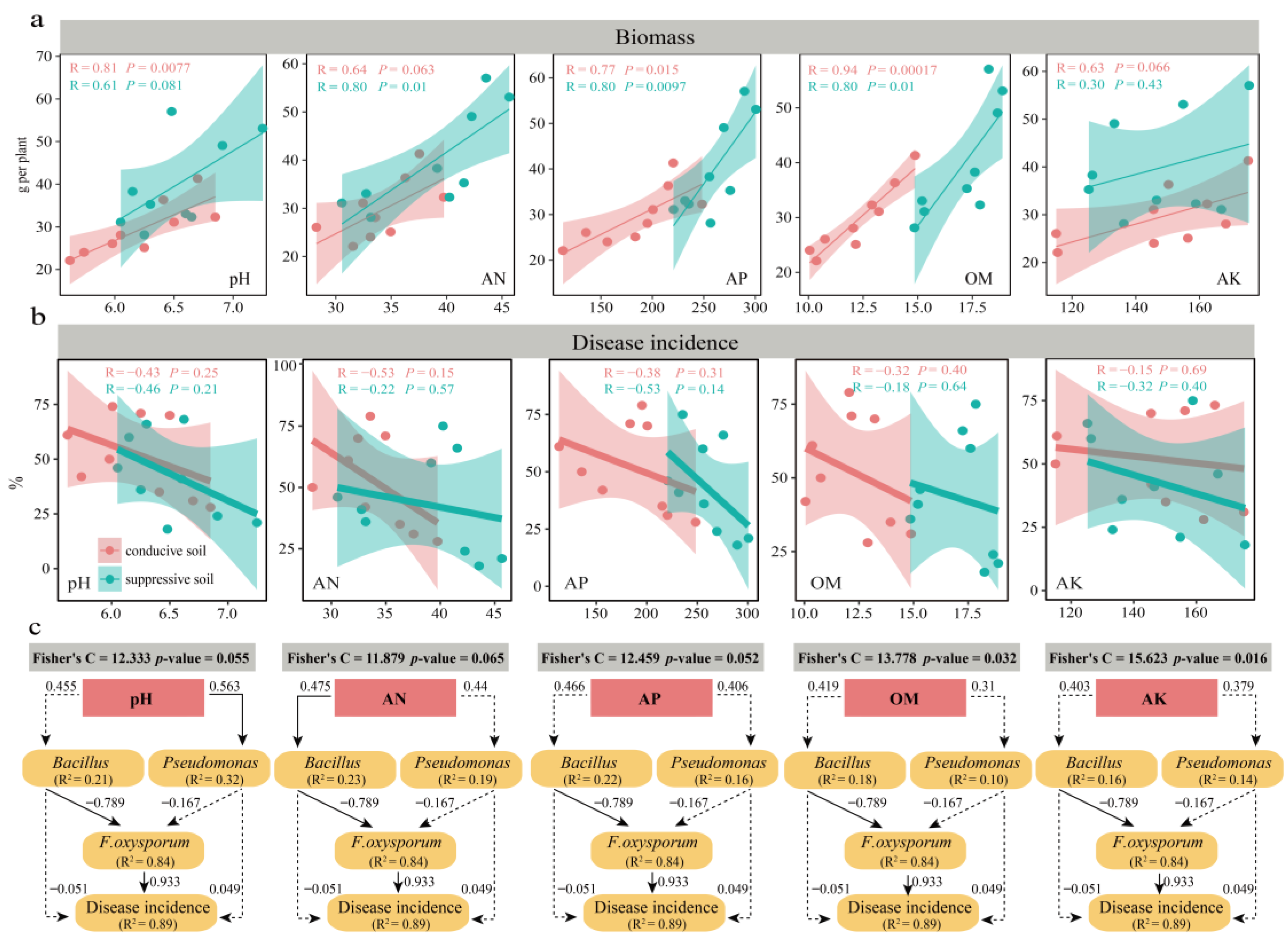
3.3. Effect of the Soil Physicochemical Association of Culturable Microorganisms on Banana Disease Incidence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.R. Fusarium oxysporum and the Fusarium Wilt Syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.; Cook, R.J. Biological Control of Plant Pathogens; WH Freeman and Company: New York, NY, USA, 1974. [Google Scholar]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Ruan, Y.; Xue, C.; Zhong, S.; Li, R.; Shen, Q. Soils naturally suppressive to banana Fusarium wilt disease harbor unique bacterial communities. Plant Soil 2015, 393, 21–33. [Google Scholar] [CrossRef]
- Deltour, P.; Franca, S.C.; Pereira, O.L.; Cardoso, I.; De Neve, S.; Debode, J.; Hofte, M. Disease suppressiveness to Fusarium wilt of banana in an agroforestry system: Influence of soil characteristics and plant community. Agric. Ecosyst. Environ. 2017, 239, 173–181. [Google Scholar] [CrossRef]
- Kyselkova, M.; Kopecky, J.; Frapolli, M.; Defago, G.; Sagova-Mareckova, M.; Grundmann, G.L.; Moenne-Loccoz, Y. Comparison of rhizobacterial community composition in soil suppressive or conducive to tobacco black root rot disease. ISME J. 2009, 3, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Shen, Z.; Xue, C.; Penton, C.R.; Thomashow, L.S.; Zhang, N.; Wang, B.; Ruan, Y.; Li, R.; Shen, Q. Suppression of banana Panama disease induced by soil microbiome reconstruction through an integrated agricultural strategy. Soil Biol. Biochem. 2019, 128, 164–174. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, B.; Hong, S.; Xiong, W.; Shen, Z.; Ruan, Y.; Li, R.; Shen, Q.; Dini-Andreote, F. Promoting soil microbial-mediated suppressiveness against Fusarium wilt disease by the enrichment of specific fungal taxa via crop rotation. Biol. Fertil. Soils 2021, 57, 1137–1153. [Google Scholar] [CrossRef]
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of fungal-mediated soil suppressiveness against Fusarium wilt disease via plant residue manipulation. Microbiome 2021, 9, 200. [Google Scholar] [CrossRef]
- Toussoun, T.; Patrick, Z.; Snyder, W.C. Influence of crop residue decomposition products on the germination of Fusarium solani f. phaseoli chlamydospores in soil. Nature 1963, 197, 1314–1316. [Google Scholar] [CrossRef]
- Bonanomi, G.; Lorito, M.; Vinale, F.; Woo, S.L. Organic Amendments, Beneficial Microbes, and Soil Microbiota: Toward a Unified Framework for Disease Suppression. Annu. Rev. Phytopathol. 2018, 56, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Honjo, H.; Becker, J.O.; Fukui, R. Temperature-dependent effects of soil amendment with crop residues on suppression of Rhizoctonia damping-off of sugar beet. Plant Soil 2013, 366, 467–477. [Google Scholar] [CrossRef]
- Williams, M.A.; Myrold, D.D.; Bottomley, P.J. Carbon flow from 13C-labeled clover and ryegrass residues into a residue-associated microbial community under field conditions. Soil Biol. Biochem. 2007, 39, 819–822. [Google Scholar] [CrossRef]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Soil immune responses Soil microbiomes may be harnessed for plant health. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef]
- Carrion, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; de Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.J.; Gomez-Exposito, R.; Elsayed, S.S.; et al. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef]
- Cha, J.-Y.; Han, S.; Hong, H.-J.; Cho, H.; Kim, D.; Kwon, Y.; Kwon, S.-K.; Crüsemann, M.; Lee, Y.B.; Kim, J.F.; et al. Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 2016, 10, 119–129. [Google Scholar] [CrossRef]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.Y.; Wu, H.S.; Jousset, A.; Shen, Q.R. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Svenningsen, N.B.; Watts-Williams, S.J.; Joner, E.J.; Battini, F.; Efthymiou, A.; Cruz-Paredes, C.; Nybroe, O.; Jakobsen, I. Suppression of the activity of arbuscular mycorrhizal fungi by the soil microbiota. ISME J. 2018, 12, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, M.; Li, G.; Liu, K.; Liu, T.; Wu, M.; Saleem, M.; Li, Z. Phosphorus availability increases pathobiome abundance and invasion of rhizosphere microbial networks by Ralstonia. Environ. Microbiol. 2021, 23, 5992–6003. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Jv, H.L.; Lu, M.; Wang, B.B.; Zhao, Y.; Ruan, Y.Z. Significant decline in banana Fusarium wilt disease is associated with soil microbiome reconstruction under chilli pepper-banana rotation. Eur. J. Soil Biol. 2020, 97, 103154. [Google Scholar] [CrossRef]
- Bao, S.D.; Su, H.Y.; An, Z.S. Soil Agrochemical Analysis, 2nd ed.; China Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Gong, B.; Bloszies, S.; Li, X.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Efficacy of garlic straw application against root-knot nematodes on tomato. Sci. Hortic. 2013, 161, 49–57. [Google Scholar] [CrossRef]
- Jeger, M.; Eden-Green, S.; Thresh, J.; Johanson, A.; Waller, J.; Brown, A. Banana Diseases. In Bananas and Plantains; Springer: Cham, Switzerland, 1995; pp. 317–381. [Google Scholar]
- Ciavatta, C.; Govi, M.; Antisari, L.V.; Sequi, P. Determination of organic carbon in aqueous extracts of soils and fertilizers. Commun. Soil Sci. Plant Anal. 1991, 22, 795–807. [Google Scholar] [CrossRef]
- Komada, H. Development of a selective medium for quantitative isolation of Fusarium oxysporum from natural soil. Rev. Plant Prot. Res. 1975, 8, 114–125. [Google Scholar]
- Li, X.; Jousset, A.; de Boer, W.; Carrion, V.J.; Zhang, T.; Wang, X.; Kuramae, E.E. Legacy of land use history determines reprogramming of plant physiology by soil microbiome. ISME J. 2019, 13, 738–751. [Google Scholar] [CrossRef]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease Suppressive Soils: New Insights from the Soil Microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef]
- Larkin, R.P.; Halloran, J.M. Management Effects of Disease-Suppressive Rotation Crops on Potato Yield and Soilborne Disease and Their Economic Implications in Potato Production. Am. J. Potato Res. 2014, 91, 429–439. [Google Scholar] [CrossRef]
- Tyler, H.L. Single-versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields. Agronomy 2021, 11, 2334. [Google Scholar] [CrossRef]
- Xun, W.; Huang, T.; Zhao, J.; Ran, W.; Wang, B.; Shen, Q.; Zhang, R. Environmental conditions rather than microbial inoculum composition determine the bacterial composition, microbial biomass and enzymatic activity of reconstructed soil microbial communities. Soil Biol. Biochem. 2015, 90, 10–18. [Google Scholar] [CrossRef]
- Lobmann, M.T.; Vetukuri, R.R.; de Zinger, L.; Alsanius, B.W.; Grenville-Briggs, L.J.; Walter, A.J. The occurrence of pathogen suppressive soils in Sweden in relation soil biota, soil properties, and farming practices. Appl. Soil Ecol. 2016, 107, 57–65. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, J.; Wu, H.; Yan, S.; Guo, D.; Wang, G.; Ma, Y. Soil chemical and microbial responses to biogas slurry amendment and its effect on Fusarium wilt suppression. Appl. Soil Ecol. 2016, 107, 116–123. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmolle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef]
- Tao, C.; Li, R.; Xiong, W.; Shen, Z.; Liu, S.; Wang, B.; Ruan, Y.; Geisen, S.; Shen, Q.; Kowalchuk, G.A. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome 2020, 8, 137. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The Soil-Borne Legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef]
- Careaga, M.; Fernandez, E.; Dorantes, L.; Mota, L.; Jaramillo, M.E.; Hernandez-Sanchez, H. Antibacterial activity of Capsicum extract against Salmonella typhimurium and Pseudomonas aeruginosa inoculated in raw beef meat. Int. J. Food Microbiol. 2003, 83, 331–335. [Google Scholar] [CrossRef]
- Bottos, E.M.; Laughlin, D.C.; Herbold, C.W.; Lee, C.K.; McDonald, I.R.; Cary, S.C. Abiotic factors influence patterns of bacterial diversity and community composition in the Dry Valleys of Antarctica. FEMS Microbiol. Ecol. 2020, 96, fiaa042. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, A.H.C.; Finckh, M.R. Plant Diseases and Management Approaches in Organic Farming Systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, D.; Watson, R. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Residue | Nutrient Content | Nutrient Content Corrections | ||||
|---|---|---|---|---|---|---|
| N (g/kg) | P2O5 (g/kg) | K2O (g/kg) | CO(NH2)2 (g/pot) | Ca(H2PO4)2 (g/pot) | K2SO4 (g/pot) | |
| CK a | 0 | 0 | 0 | 3.81 | 2.52 | 13.61 |
| Banana | 10.50 | 3.35 | 70.75 | 1.52 | 1.23 | 0 |
| Pepper | 17.51 | 6.55 | 35.35 | 0 | 0 | 6.81 |
| Target | Conducive Soil | Suppressive Soil |
|---|---|---|
| Site | Jianfeng Town | Jianfeng Town |
| Latitude | 18.753° N | 18.723° N |
| Longitude | 108.648° E | 108.688° E |
| Temperature (°C) a | 24 | 24 |
| Precipitation (mm) b | 1150 | 1150 |
| Monocultured year | 15 | 15 |
| Soil type | Dry red | Dry red |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.; Jv, H.; Yuan, X.; Geng, J.; Wang, B.; Zhao, Y.; Wang, Q.; Li, R.; Jia, Z.; Ruan, Y. Soil Organic Nitrogen Indirectly Enhances Pepper-Residue-Mediated Soil Disease Suppression through Manipulation of Soil Microbiome. Agronomy 2022, 12, 2077. https://doi.org/10.3390/agronomy12092077
Hong S, Jv H, Yuan X, Geng J, Wang B, Zhao Y, Wang Q, Li R, Jia Z, Ruan Y. Soil Organic Nitrogen Indirectly Enhances Pepper-Residue-Mediated Soil Disease Suppression through Manipulation of Soil Microbiome. Agronomy. 2022; 12(9):2077. https://doi.org/10.3390/agronomy12092077
Chicago/Turabian StyleHong, Shan, Hongling Jv, Xianfu Yuan, Jianjian Geng, Beibei Wang, Yan Zhao, Qing Wang, Rong Li, Zhongjun Jia, and Yunze Ruan. 2022. "Soil Organic Nitrogen Indirectly Enhances Pepper-Residue-Mediated Soil Disease Suppression through Manipulation of Soil Microbiome" Agronomy 12, no. 9: 2077. https://doi.org/10.3390/agronomy12092077
APA StyleHong, S., Jv, H., Yuan, X., Geng, J., Wang, B., Zhao, Y., Wang, Q., Li, R., Jia, Z., & Ruan, Y. (2022). Soil Organic Nitrogen Indirectly Enhances Pepper-Residue-Mediated Soil Disease Suppression through Manipulation of Soil Microbiome. Agronomy, 12(9), 2077. https://doi.org/10.3390/agronomy12092077