


Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Pathogen Isolation
2.2. DNA Extraction from Phlyctema Vagabunda Isolates
2.3. Fungal DNA Amplification
2.4. Nucleotide Sequencing and Phylogenetic Analyses
2.5. Effect of Temperature on Mycelial Growth and Conidial Production of Phlyctema Vagabunda
2.6. Pathogenicity of Phlyctema Vagabunda Isolates In Vivo Condition
2.7. Propolis Extract Source
2.8. Effect of Methanolic Propolis Extracts on the Mycelial Growth of Phlyctema Vagabunda Isolates
2.9. Effect of Methanolic Propolis Extract on the Conidial Germination in In Vitro Condition
2.10. Effect of Propolis Extract on the Control of Phlyctema Vagabunda in Apple
2.11. Data Analysis
3. Results
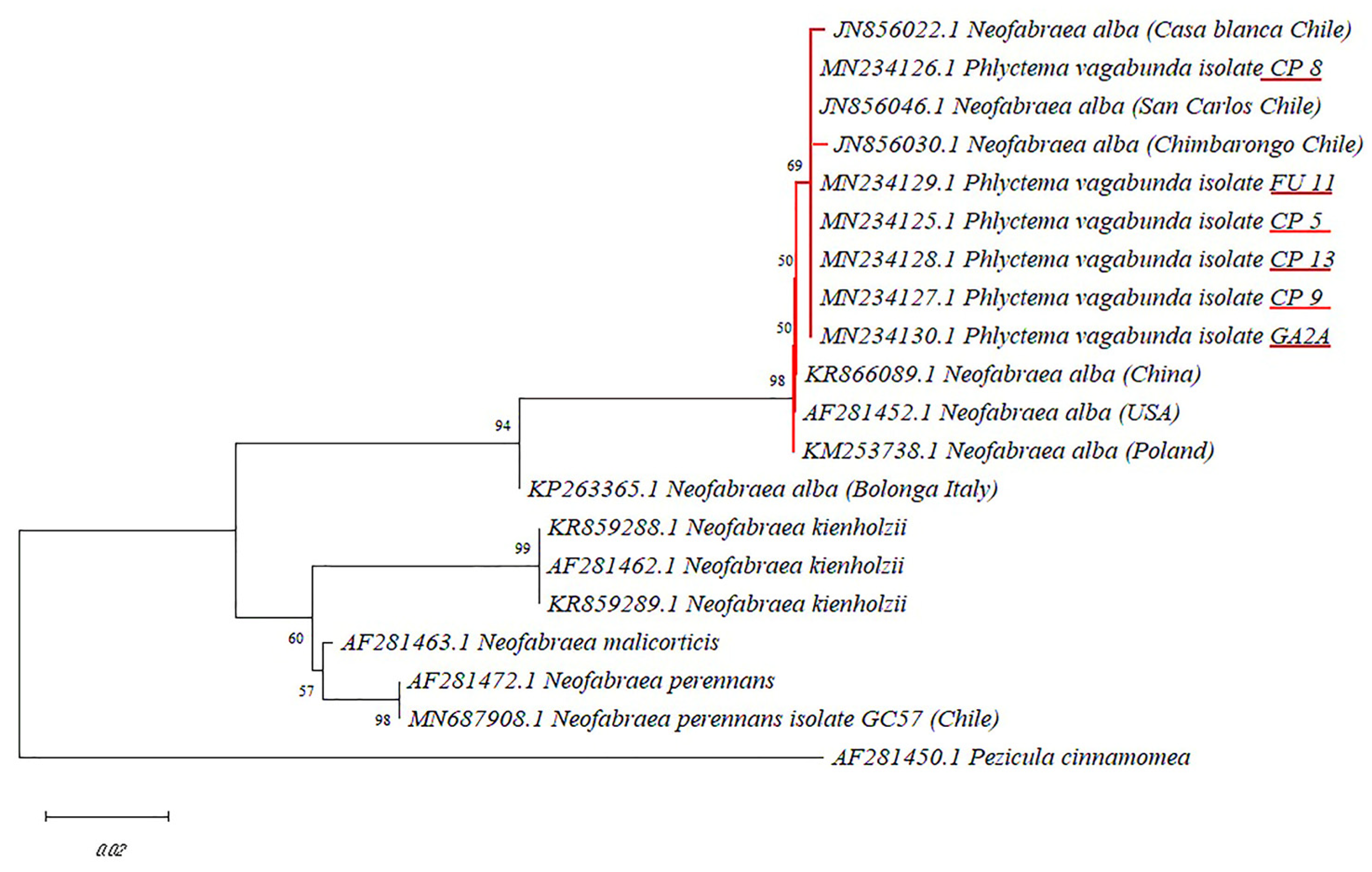
3.1. Isolation, Characterization and Molecular Identification of the Chilean Phlyctema Vagabunda Isolates
3.2. Effect of Temperature on Mycelial Growth and Conidial Production of Phlyctema Vagabunda
3.3. Pathogenicity of Chilean Phlyctema Vagabunda Isolates on Apple Fruit
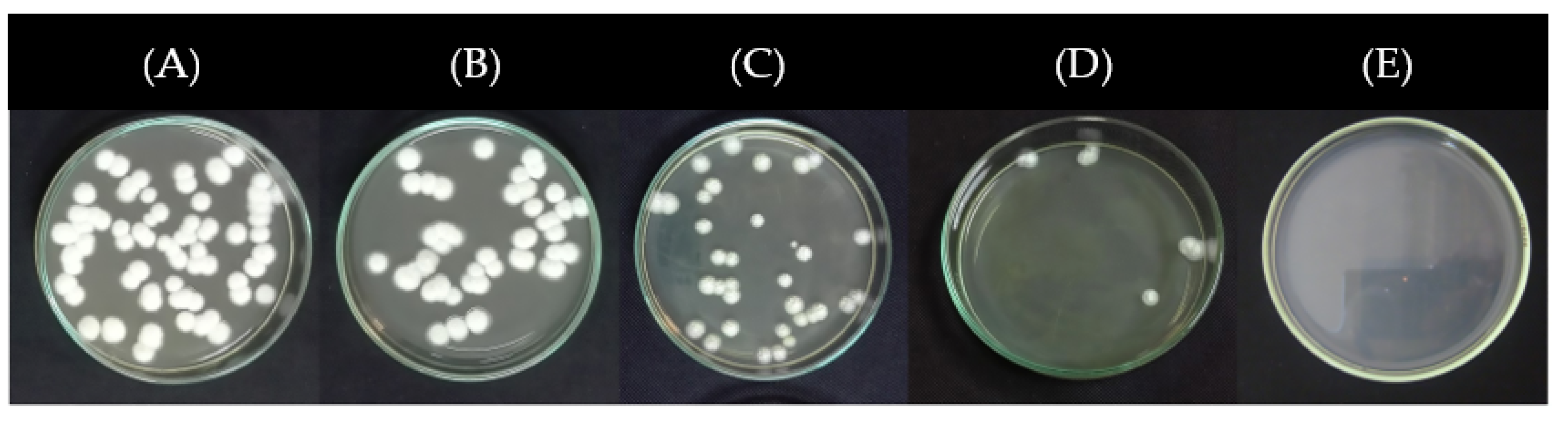
3.4. Effect of Propolis Extracts on the Mycelial Growth of Phlyctema Vagabunda
3.5. Effect of Propolis Extract on Conidial Germination of Phlyctema Vagabunda in In Vitro Condition
3.6. Effect of Propolis Extract LR16 on the Control of Phlyctema Vagabunda CP13 Isolate in In Vivo Condition
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cameldi, I.; Neri, F.; Menghini, M.; Pirondi, A.; Nanni, I.M.; Collina, M.; Mari, M. Characterization of Neofabraea vagabunda isolates causing apple bull’s eye rot in Italy (Emilia-Romagna region). Plant Pathol. 2017, 66, 1432–1444. [Google Scholar] [CrossRef]
- Den Breeyen, A.; Rochefort, J.; Russouw, A.; Meitz-Hopkins, J.; Lennox, C.L. Preharvest detection and postharvest incidence of Phlyctema vagabunda on ‘Cripps Pink’ apples in South Africa. Plant Dis. 2020, 104, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Giraud, M.; Coureau, C.; Perrin, J.; Westercamp, P. Apple lenticel rots: State of knowledge on the epidemiology of Neofabraea vagabunda. Acta Hortic. 2021, 1325, 59–66. [Google Scholar] [CrossRef]
- Soto-Alvear, S.; Lolas, M.; Rosales, I.M.; Chávez, E.R.; Latorre, B.A. Characterization of the bull’s eye rot of apple in Chile. Plant Dis. 2013, 97, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriquez, J.L. First report of apple rot caused by Neofabraea alba in Chile. Plant Dis. 2005, 89, 1360. [Google Scholar] [CrossRef] [PubMed]
- González, F.; Salinas, C.; Pinilla, B.; Castillo, A. First report of apple bull’s-eye rot caused by Neofabraea perennans in Chile. Plant Dis. 2020, 104, 1537. [Google Scholar] [CrossRef]
- Aguilar, C.G.; Mazzola, M.; Xiao, C.L. Timing of apple fruit infection by Neofabraea perennans and Neofabraea kienholzii in relation to bull’s-eye rot development in stored apple fruit. Plant Dis. 2017, 101, 800–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spotts, R.A.; Seifert, K.A.; Wallis, K.M.; Sugar, D.; Xiao, C.L.; Serdani, M.; Henriquez, J.L. Description of Cryptosporiopsis kienholzii and species profiles of Neofabraea in major pome fruit growing districts in the Pacific Northwest USA. Mycol. Res. 2009, 113, 1301–1311. [Google Scholar] [CrossRef]
- Vukotić, J.; Stojšin, V.; Nagl, N.; Petreš, M.; Hrustić, J.; Grahovac, M.; Tanović, B. Morphological, molecular, and pathogenic characterization of Neofabraea alba, a postharvest pathogen of apple in Serbia. Agronomy 2022, 12, 2015. [Google Scholar] [CrossRef]
- Lin, H.; Jiang, X.; Yi, J.; Wang, X.; Zuo, R.; Jiang, Z.; Wang, W.; Zhou, E. Molecular identification of Neofabraea species associated with bull’s-eye rot on apple using rolling-circle amplification of partial EF-1α sequence. Can. J. Microbiol. 2018, 64, 57–68. [Google Scholar] [CrossRef]
- Wood, P.N.; Fisher, B.M. The effect of fungicides on spore germination, mycelial growth and lesion development of Phlyctema vagabunda (syn: Neofabraea alba) (bull’s eye rot of apples). N. Z. Plant Prot. 2017, 70, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Wenneker, M.; Thomma, B.P.H.J. Latent postharvest pathogens of pome fruit and their management: From single measures to a systems intervention approach. Eur. J. Plant Pathol. 2020, 156, 663–681. [Google Scholar] [CrossRef] [Green Version]
- Lolas, M.; Díaz, G.; Mendez, R.; Cáceres, M.; Neubauer, L. Evaluation of the efficacy of fungicide fludioxonil in the postharvest control of bull’s eye rot (Neofabraea alba) in Chile. Acta Hortic. 2016, 1144, 461–464. [Google Scholar] [CrossRef]
- Mesías, F.J.; Martín, A.; Hernández, A. Consumers’ growing appetite for natural foods: Perceptions towards the use of natural preservatives in fresh fruit. Food Res. Int. 2021, 150, 110749. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fernández-Ortuño, D.; Grabke, A.; Schnabel, G. Resistance to fludioxonil in Botrytis cinerea isolates from blackberry and strawberry. Phytopathology 2014, 104, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.A.; Vallejo, A.M.; Ballester, A.-R.; Zampini, C.; Isla, M.I.; López-Rubio, A.; Fabra, M.J. Antifungal edible coatings containing Argentinian propolis extract and their application in raspberries. Food Hydrocoll. 2020, 107, 105973. [Google Scholar] [CrossRef]
- Pobiega, K.; Kraśniewska, K.; Derewiaka, D.; Gniewosz, M. Comparison of the antimicrobial activity of propolis extracts obtained by means of various extraction methods. J. Food Sci. Technol. 2019, 56, 5386–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Zhou, Y.; Ye, J.; Fan, G.; Peng, L.; Pan, S. Effects of poplar buds as an alternative to propolis on postharvest diseases control of strawberry fruits: Poplar buds to control postharvest disease of strawberry. J. Sci. Food Agric. 2016, 96, 2136–2141. [Google Scholar] [CrossRef]
- Soylu, E.M.; Ozdemir, A.E.; Erturk, E.; Sahinler, N.; Soylu, S. Antifungal activity of propolis against postharvest disease agent Penicillium digitatum. Asian J. Chem. 2008, 20, 4823–4830. [Google Scholar]
- Akbay, E.; Özenirler, Ç.; Çelemli, Ö.G.; Durukan, A.B.; Onur, M.A.; Sorkun, K. Effects of propolis on warfarin efficacy. Pol. J. Cardio-Thorac. Surg. 2017, 1, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Kasiotis, K.M.; Anastasiadou, P.; Papadopoulos, A.; Machera, K. Revisiting Greek propolis: Chromatographic analysis and antioxidant activity study. PLoS ONE 2017, 12, e0170077. [Google Scholar] [CrossRef] [PubMed]
- Mutlu Sariguzel, F.; Berk, E.; Koc, A.N.; Sav, H.; Demir, G. Antifungal activity of propolis against yeasts isolated from blood culture: In vitro evaluation: Antifungal activity of propolis. J. Clin. Lab. Anal. 2016, 30, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, R.; Igoli, J.O.; Zhang, T.; Gray, A.I.; Ebiloma, G.U.; Clements, C.J.; Fearnley, J.; Edrada Ebel, R.; Paget, T.; de Koning, H.P.; et al. The chemical characterization of nigerian propolis samples and their activity against Trypanosoma brucei. Sci. Rep. 2017, 7, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šturm, L.; Ulrih, N.P. Advances in the propolis chemical composition between 2013 and 2018: A review. eFood 2020, 1, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Montalva, C.; Arismendi, N.; Barta, M.; Rojas, E. Molecular differentiation of recently described Neozygites osornensis (Neozygitales: Neozygitaceae) from two morphologically similar species. J. Invertebr. Pathol. 2014, 115, 92–94. [Google Scholar] [CrossRef]
- Cao, D.; Li, X.; Cao, J.; Wang, W. PCR detection of the three neofabraea pathogenic species responsible for apple bull’s eye rot. Adv. Microbiol. 2013, 3, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Vero, S.; Mondino, P.; Burgueño, J.; Soubes, M.; Wisniewski, M. Characterization of biocontrol activity of two yeast strains from Uruguay against blue mold of apple. Postharvest Biol. Technol. 2002, 26, 91–98. [Google Scholar] [CrossRef]
- Arismendi, N.; Vargas, M.; López, M.D.; Barría, Y.; Zapata, N. Promising antimicrobial activity against the honey bee parasite Nosema ceranae by methanolic extracts from Chilean native plants and propolis. J. Apic. Res. 2018, 57, 522–535. [Google Scholar] [CrossRef]
- Mohammedi, Z.; Atik, F. Fungitoxic effect of natural extracts on mycelial growth, spore germination and aflatoxin B1 production of Aspergillus flavus. Aust. J. Crop Sci. 2013, 7, 293–298. [Google Scholar]
- Daniel, C.K.; Lennox, C.L.; Vries, F.A. In vivo application of garlic extracts in combination with clove oil to prevent postharvest decay caused by Botrytis cinerea, Penicillium expansum and Neofabraea alba on apples. Postharvest Biol. Technol. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Neri, F.; Crucitti, D.; Negrini, F.; Pacifico, D.; Ceredi, G.; Carimi, F.; Lolas, M.A.; Collina, M.; Baraldi, E. New insight into morphological and genetic diversity of Phlyctema vagabunda and Neofabraea kienholzii causing bull’s eye rot on apple and pear. Plant Pathol. 2022, 1–22. [Google Scholar] [CrossRef]
- Hortova, B.; Novotny, D.; Erban, T. Physiological characteristics and pathogenicity of eight Neofabraea isolates from apples in Czechia. Eur. J. Hortic. Sci. 2014, 79, 327–334. [Google Scholar]
- Amaral Carneiro, G.; Walcher, M.; Storti, A.; Baric, S. Phylogenetic diversity and phenotypic characterization of Phlyctema vagabunda (syn. Neofabraea alba ) and Neofabraea kienholzii causing postharvest bull’s eye rot of apple in Northern Italy. Plant Dis. 2022, 106, 451–463. [Google Scholar] [CrossRef] [PubMed]
- de Jong, S.N.; Lévesque, C.A.; Verkley, G.J.M.; Abeln, E.C.A.; Rahe, J.E.; Braun, P.G. Phylogenetic relationships among Neofabraea species causing tree cankers and bull’s-eye rot of apple based on DNA sequencing of ITS nuclear rDNA, mitochondrial rDNA, and the β-tubulin gene. Mycol. Res. 2001, 105, 658–669. [Google Scholar] [CrossRef]
- Ali, A.; Cheong, C.; Noosheen, Z. Composite effect of propolis and gum arabic to control postharvest anthracnose and maintain quality of papaya during storage. Int. J. Agric. Biol. 2014, 1, 1117–1122. [Google Scholar]
- Yang, S.Z.; Peng, L.T.; Su, X.J.; Chen, F.; Cheng, Y.J.; Fan, G.; Pan, S.Y. Bioassay-guided isolation and identification of antifungal components from propolis against Penicillium italicum. Food Chem. 2011, 127, 210–215. [Google Scholar] [CrossRef]
- Petruzzi, L.; Rosaria Corbo, M.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Bevilacqua, A. Antifungal and antibacterial effect of propolis: A comparative hit for food-borne pseudomonas, enterobacteriaceae and fungi. Foods 2020, 9, 559. [Google Scholar] [CrossRef]
- Paula, L.A.L.; Cândido, A.C.B.B.; Santos, M.F.C.; Caffrey, C.R.; Bastos, J.K.; Ambrósio, S.R.; Magalhães, L.G. Antiparasitic properties of propolis extracts and their compounds. Chem. Biodivers. 2021, 18, e2100310. [Google Scholar] [CrossRef]
- Barrientos, L.; Herrera, C.L.; Montenegro, G.; Ortega, X.; Veloz, J.; Alvear, M.; Cuevas, A.; Saavedra, N.; Salazar, L.A. Chemical and botanical characterization of Chilean propolis and biological activity on cariogenic bacteria Streptococcus mutans and Streptococcus sobrinus. Braz. J. Microbiol. 2013, 44, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Wagh, V.D. Propolis: A wonder bees product and its pharmacological potentials. Adv. Pharmacol. Sci. 2013, 2013, 308249. [Google Scholar] [CrossRef] [Green Version]
- Bueno-Silva, B.; Marsola, A.; Ikegaki, M.; Alencar, S.M.; Rosalen, P.L. The effect of seasons on Brazilian red propolis and its botanical source: Chemical composition and antibacterial activity. Nat. Prod. Res. 2017, 31, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, E.N.; Sampietro, D.A.; Soberon, J.R.; Sgariglia, M.A.; Vattuone, M.A. Propolis from the northwest of Argentina as a source of antifungal principles. J. Appl. Microbiol. 2006, 101, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Probst, I.; Sforcin, J.; Vlm, R.; Fernandes, A.; Fernandes Júnior, A. Antimicrobial activity of propolis and essential oils and synergism between these natural products. J. Venom. Anim. Toxins Trop. Dis. 2011, 17, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Peng, L.; Cheng, Y.; Chen, F.; Pan, S. Control of citrus green and blue molds by Chinese propolis. Food Sci. Biotechnol. 2010, 19, 1303–1308. [Google Scholar] [CrossRef]
- Peng, L.; Yang, S.; Cheng, Y.J.; Chen, F.; Pan, S.; Fan, G. Antifungal activity and action mode of pinocembrin from propolis against Penicillium italicum. Food Sci. Biotechnol. 2012, 21, 1533–1539. [Google Scholar] [CrossRef]
- Pereira, L.; Cunha, A.; Almeida-Aguiar, C. Portuguese propolis from Caramulo as a biocontrol agent of the apple blue mold. Food Control 2022, 139, 109071. [Google Scholar] [CrossRef]
- Loebler, M.; Sánchez, C.; Muchagato Maurício, E.; Diogo, E.; Santos, M.; Vasilenko, P.; Cruz, A.S.; Mendes, B.; Gonçalves, M.; Duarte, M.P. Potential application of propolis extracts to control the growth of Stemphylium vesicarium in “Rocha” pear. Appl. Sci. 2020, 10, 1990. [Google Scholar] [CrossRef] [Green Version]
- Marino, A.K.; Junior, J.S.P.; Magalhães, K.M.; Mattiuz, B.-H. Chitosan-propolis combination inhibits anthracnose in “Hass” avocados. Emir. J. Food Agric. 2018, 681. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.-L. The effects of temperature and nutritional conditions on mycelium growth of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Isolate | Mean (SE) | Conidial Production |
|---|---|---|---|
| 20 | CP 13 | 20.50 (0.5) a | − |
| 15 | CP 5 | 19.92 (0.36) ab | − |
| 20 | CP 8 | 19.67 (0.66) ab | − |
| 15 | CP 9 | 19.50 (1.00) ab | − |
| 15 | CP 13 | 19.50 (0.00) ab | − |
| 15 | CP 8 | 19.17 (0.44) ab | − |
| 20 | CP 9 | 19.17 (0.22) ab | − |
| 20 | CP 5 | 17.75 (0.8) b | − |
| 10 | CP 9 | 15.00 (0.00) c | − |
| 25 | CP 5 | 14.50 (1.00) c | − |
| 10 | CP 8 | 14.33 (0.08) cd | − |
| 10 | CP 5 | 13.58 (0.22) cd | − |
| 10 | CP 13 | 12.92 (0.30) cd | − |
| 25 | CP 9 | 11.67 (0.16) de | − |
| 25 | CP 13 | 9.83 (0.88) ef | − |
| 25 | CP 8 | 8.17 (0.22) fg | − |
| 5 | CP 5 | 6.33 (0.66) gh | + |
| 5 | CP 8 | 5.50 (0.00) ghi | + |
| 5 | CP 9 | 5.50 (0.28) ghi | + |
| 5 | CP 13 | 5.17 (0.92) hi | + |
| 0 | CP 13 | 3.83 (0.16) hi | + |
| 0 | CP 8 | 3.50 (0.50) i | + |
| 0 | CP 9 | 3.25 (0.38) i | + |
| 0 | CP 5 | 3.00 (0.00) i | + |
| 30 | CP 9 | 0.00 (0.00) j | − |
| 30 | CP 13 | 0.00 (0.00) j | − |
| 30 | CP 8 | 0.00 (0.00) j | − |
| 30 | CP 5 | 0.00 (0.00) j | − |
| Isolate | Growth Temperature (°C) | Conidia N° Colonies−1 | Length (µm) | Width (µm) |
|---|---|---|---|---|
| CP 5 | 0 | 1.44 × 105 ± 2.6 × 104 ab | 10.42 ± 1.9 a | 2.74 ± 0.2 a |
| CP 8 | 0 | 1.78 × 105 ± 1.0 × 104 a | 10.42 ± 1.4 a | 2.97 ± 0.1 a |
| CP 9 | 0 | 9.68 × 104 ± 2.2 × 103 b | 9.99 ± 1.9 a | 2.79 ± 0.1 a |
| CP 13 | 0 | 1.03 × 105 ± 1.4 × 103 b | 10.78 ± 2.5 a | 3.06 ± 0.1 a |
| CP 5 | 5 | 1.05 × 105 ± 1.3 × 104 b | 10,94 ± 1.6 a | 2.87 ± 0.2 a |
| CP 8 | 5 | 1.34 × 105 ± 2.5 × 104 ab | 10,10 ± 2.4 a | 2.83 ± 0.2 a |
| CP 9 | 5 | 9.91 × 104 ± 3.3 × 103 b | 10.72 ± 0.9 a | 2.87 ± 0.2 a |
| CP 13 | 5 | 2.02 × 105 ± 8.6 × 102 a | 11.11 ± 2.5 a | 3.05 ± 0.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urrea, I.; Arismendi, N.; Sepúlveda, X.; Gerding, M.; Vero, S.; Vargas, M. Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda. Agronomy 2023, 13, 104. https://doi.org/10.3390/agronomy13010104
Urrea I, Arismendi N, Sepúlveda X, Gerding M, Vero S, Vargas M. Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda. Agronomy. 2023; 13(1):104. https://doi.org/10.3390/agronomy13010104
Chicago/Turabian StyleUrrea, Iván, Nolberto Arismendi, Ximena Sepúlveda, Macarena Gerding, Silvana Vero, and Marisol Vargas. 2023. "Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda" Agronomy 13, no. 1: 104. https://doi.org/10.3390/agronomy13010104
APA StyleUrrea, I., Arismendi, N., Sepúlveda, X., Gerding, M., Vero, S., & Vargas, M. (2023). Antifungal Activity of Propolis Extracts against Postharvest Pathogen Phlyctema vagabunda. Agronomy, 13(1), 104. https://doi.org/10.3390/agronomy13010104