
How Much Phosphorus Uptake Is Required for Achieving Maximum Maize Grain Yield? Part 2: Impact of Phosphorus Uptake on Grain Quality and Partitioning of Nutrients



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
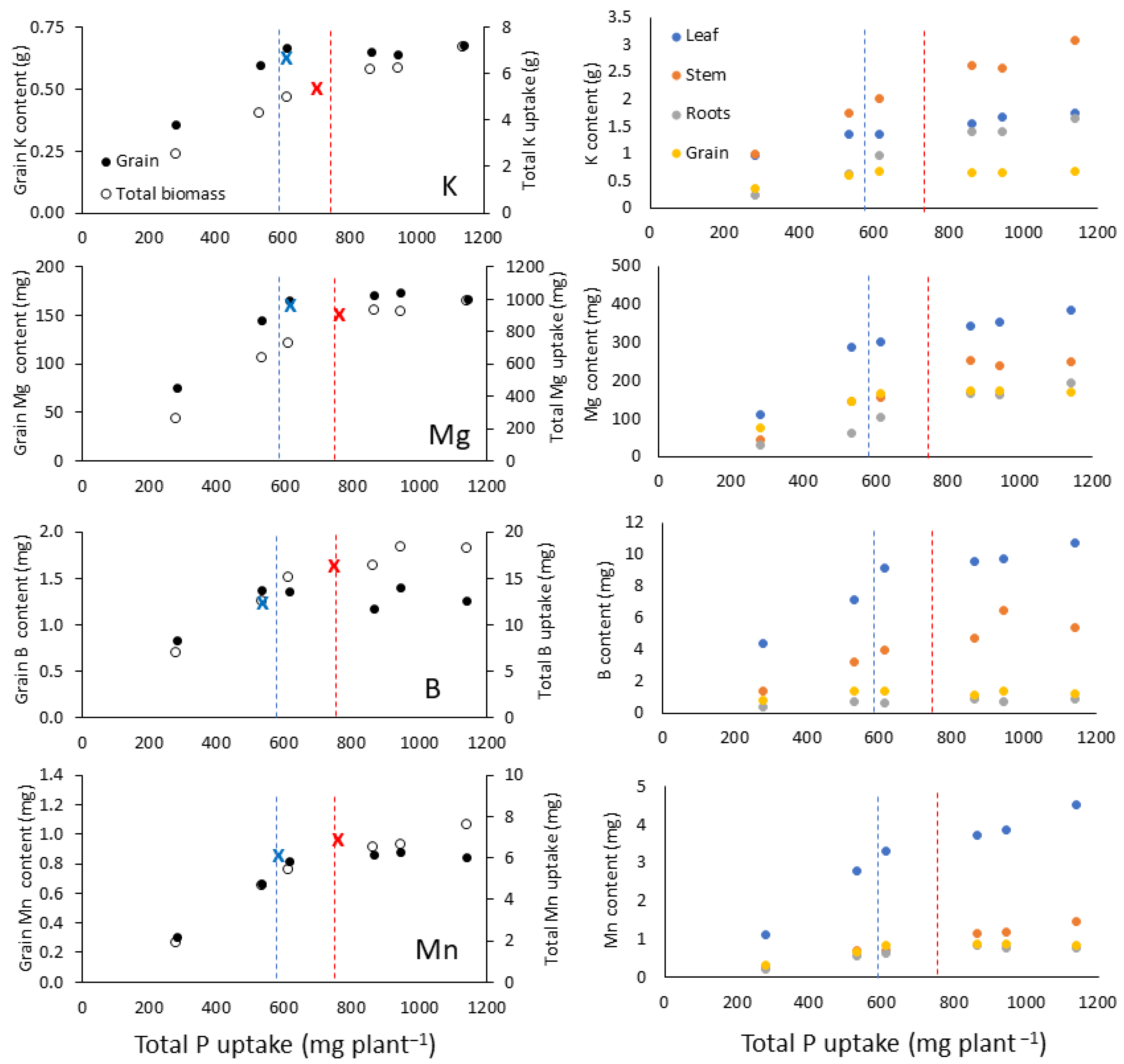
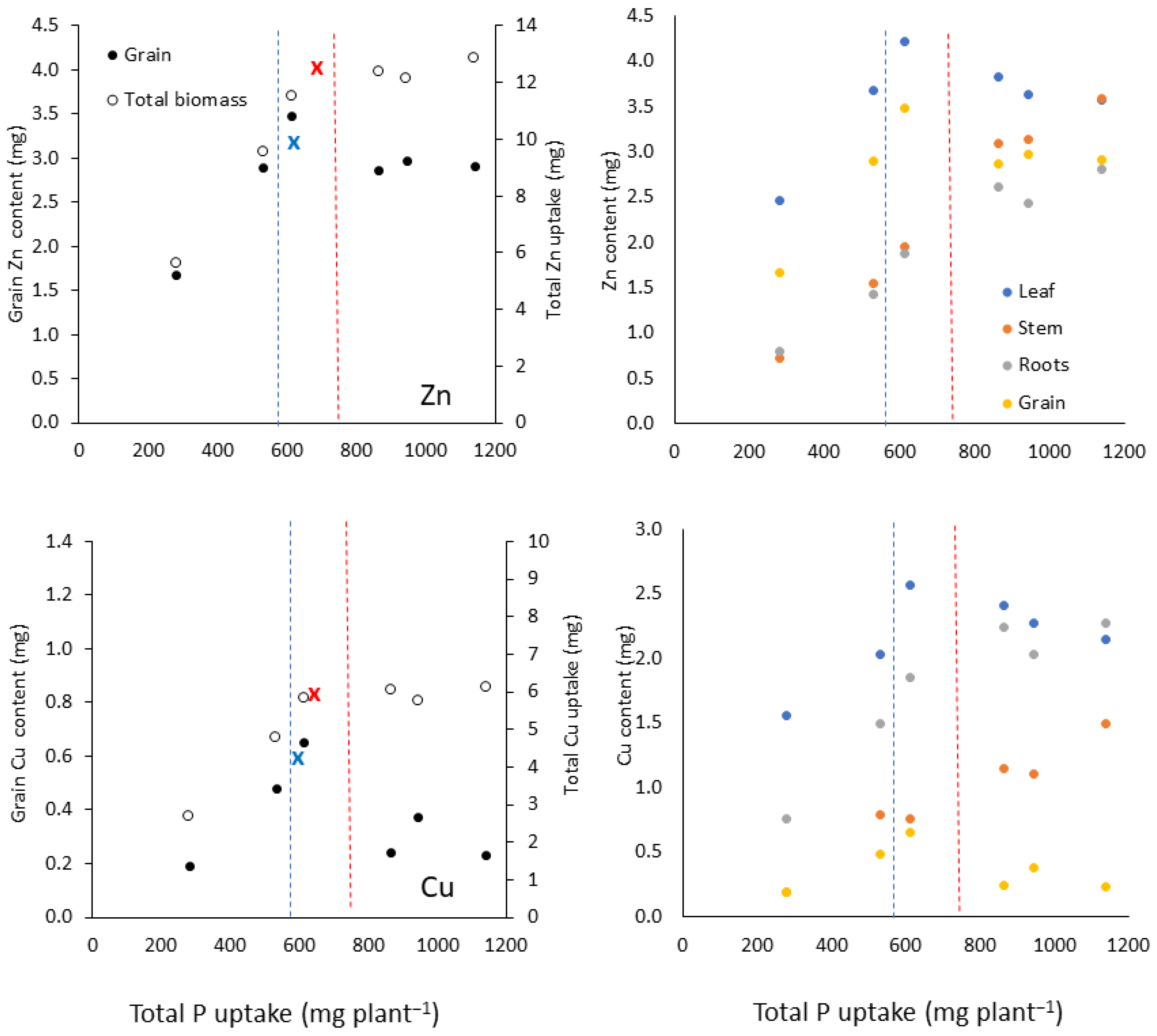
3.1. Nutrient Partitioning in Response to Phosphorus Uptake


{kind=link}
{kind=link}
{kind=link}
| Nutrient | Plant Part | Model | P Uptake Breakpoint | Intercept | Slope | Slope2 | Joint Level | R2 |
|---|---|---|---|---|---|---|---|---|
| (mg P plant−1) | mg or g plant−1 | mg or g mg P−1 | mg or g mg P−1 | mg or g plant−1 | ||||
| K (g) | Leaf | Linear-linear | 628 | 0.63 | 0.001 | 0.0007 | 1.41 | 0.98 *** |
| Stem | Linear-linear | 664 | 0.12 | 0.003 | 0.0018 | 2.16 | 0.99 *** | |
| Roots | Linear-linear | 832 | −0.36 | 0.002 | 0.0009 | 1.33 | 0.99 *** | |
| Grain | Linear-plateau | 595 | 0.094 | 0.001 | NA | 0.66 | 0.99 *** | |
| Total | Linear-linear | 679 | 0.50 | 0.007 | 0.0036 | 5.44 | 0.99 ** | |
| Mg (g) | Leaf | Linear-linear | 533 | −89 | 0.71 | 0.16 | 288 | 0.99 *** |
| Stem | Linear-plateau | 840 | −54 | 0.36 | NA | 245 | 0.99 *** | |
| Roots | Linear-plateau | 1041 | −37 | 0.22 | NA | 192 | 0.97 ** | |
| Grain | Linear-plateau | 630 | −0.81 | 0.27 | NA | 170 | 0.99 *** | |
| Total | Linear-plateau | 760 | −139 | 1.4 | NA | 950 | 0.99 *** | |
| B (mg) | Leaf | Linear-linear | 612 | 0.50 | 0.014 | 0.0035 | 8.78 | 0.98 *** |
| Stem | Linear-plateau | 823 | −0.77 | 0.008 | NA | 5.53 | 0.90 ** | |
| Roots | Linear-plateau | 800 | 0.16 | 0.0008 | NA | 0.85 | 0.82 * | |
| Grain | Linear-plateau | 527 | 0.31 | 0.002 | NA | 1.31 | 0.84 * | |
| Total | Linear-plateau | 734 | 0.33 | 0.024 | NA | 17.7 | 0.97 ** | |
| Mn (mg) | Leaf | Linear-linear | 583 | −0.79 | 0.007 | 0.002 | 3.14 | 0.99 *** |
| Stem | Linear-plateau | 890 | 0.16 | 0.004 | NA | 4.15 | 0.90 ** | |
| Roots | Linear-plateau | 742 | −0.13 | 0.001 | NA | 0.79 | 0.98 *** | |
| Grain | Linear-plateau | 647 | −0.120 | 0.002 | NA | NA | 0.99 *** | |
| Total | Linear-plateau | 747 | −1.10 | 0.011 | NA | NA | 0.97 ** |

| Nutrient | Plant Part | Model | P Uptake Breakpoint | Intercept | Slope | Slope2 | Joint Level | R2 |
|---|---|---|---|---|---|---|---|---|
| (mg P plant−1) | mg or g plant−1 | mg or g mg P−1 | mg or g mg P−1 | mg or g plant−1 | ||||
| N (g) | Leaf | Linear-plateau | 926 | 0.38 | 0.001 | NA | 1.32 | 0.93 ** |
| Stem | Linear-linear | 864 | −0.34 | 0.003 | 0.002 | 2.33 | 0.99 ** | |
| Roots | Linear-plateau | 758 | −0.27 | 0.002 | NA | 1.27 | 0.97 *** | |
| Grain | Linear-linear | 523 | 0.503 | 0.006 | −0.001 | 3.64 | 0.95 * | |
| Total | Linear-plateau | 718 | 0.82 | 0.010 | NA | 7.71 | 0.98 ** | |
| S (g) | Leaf | Linear-plateau | 526 | −45.6 | 0.77 | NA | 390 | 0.96 *** |
| Stem | Linear-linear | 757 | −48.9 | 0.37 | 0.15 | 229 | 0.99 ** | |
| Roots | Linear-plateau | 828 | −126 | 0.72 | NA | 470 | 0.99 *** | |
| Grain | Linear-linear | 570 | 32.2 | 0.41 | −0.075 | 264 | 0.98 * | |
| Total | Linear-plateau | 703 | −145 | 2.1 | NA | 1355 | 0.99 *** | |
| Fe (mg) | Leaf | Linear-plateau | 855 | 0.47 | 0.006 | NA | 5.57 | 0.99 *** |
| Stem | Linear-linear | 885 | −1.39 | 0.008 | 0.006 | 5.83 | 0.99 *** | |
| Roots | Linear-plateau | 745 | −5.31 | 0.062 | NA | 2.62 | 0.94 ** | |
| Grain | Linear-linear | 589 | 0.16 | 0.006 | −0.0014 | 3.58 | 0.96 * | |
| Total | Linear-plateau | 758 | −5.7 | 0.08 | NA | 56 | 0.97 ** |

| Nutrient | Grain or Total Content | Model | P Uptake Breakpoint | Intercept | Slope | Slope2 | Joint Level | R2 |
|---|---|---|---|---|---|---|---|---|
| (mg P plant−1) | mg plant−1 | mg mg P−1 | mg mg P−1 | mg plant−1 | ||||
| Zn (mg) | Leaf | Linear-linear | 613 | 1.15 | 0.005 | −0.001 | 4.10 | 0.98 * |
| Stem | Linear-linear | 864 | −0.46 | 0.004 | 0.002 | 2.97 | 0.99 *** | |
| Roots | Linear-linear | 846 | −0.08 | 0.003 | 0.0009 | 2.49 | 0.98 * | |
| Grain | Linear-linear | 612 | 0.29 | 0.005 | −0.00095 | 3.31 | 0.96 * | |
| Total | Linear-plateau | 683 | 0.78 | 0.017 | NA | 12.5 | 0.99 *** | |
| Cu (mg) | Leaf | Linear-linear | 674 | 0.8 | 0.003 | −0.001 | 2.59 | 0.88 * |
| Stem | Linear-linear | 532 | −0.39 | 0.002 | 0.001 | 0.71 | 0.97 * | |
| Roots | Linear-plateau | 731 | −0.14 | 0.003 | NA | 2.18 | 0.97 *** | |
| Grain | Linear-linear | 612 | −0.13 | 0.0012 | −0.0007 | 0.58 | 0.80 * | |
| Total | Linear-plateau | 643 | 0.094 | 0.0092 | NA | 6.0 | 0.97 ** |
3.1.1. Group 1 Nutrients: K, Mg, B, and Mn
3.1.2. Group 2 Nutrients: N, S, and Fe
3.1.3. Group 3 Nutrients: Zn and Cu
3.2. Nutrient Partitioning and the Decrease in Grain Yield with Excess P Uptake
4. Implications and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Penn, C.J.; Camberato, M.W. How Much Phosphorus Uptake is Required for Achieving Maximum Maize Grain Yield? Part 1: Luxury Consumption and Implications for Yield. Agronomy 2023, 13, 95. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Nutrient sufficiency concepts for modern corn hybrids: Impacts of management practices and yield levels. Crop Manag. 2014, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mathan, K.K.; Amberger, A. Influence of iron on the uptake of phosphorus by maize. Plant Soil 1977, 46, 413–422. [Google Scholar] [CrossRef]
- Awan, Z.I.; Abbasi, M.K. Interactive effect of phosphorus and copper on maize growth. Pakistan J. Agric. Res. 2000, 16, 105–108. [Google Scholar]
- Zhang, W.; Zou, C.; Chen, X.; Liu, Y.; Liu, D.; Yang, H.; Deng, Y.; Chen, X. Phosphorus application decreased copper concentration but not iron in maize grain. Agronomy 2020, 10, 1716. [Google Scholar] [CrossRef]
- Safaya, N.M. Phosphorus-zinc interaction in relation to absorption rates of phosphorus, zinc, copper, manganese, and iron in corn. Soil Sci. Soc. Am. J. 1976, 40, 719–722. [Google Scholar] [CrossRef]
- Izsáki, Z. Effects of phosphorus supplies on the nutritional status of maize (Zea mays L.). Commun. Soil Sci. Plant Anal. 2014, 45, 516–529. [Google Scholar] [CrossRef]
- Wiethorn, M.; Penn, C.; Camberato, J. A Research Method for Semi-Automated Large-Scale Cultivation of Maize to Full Maturity in an Artificial Environment. Agronomy 2021, 11, 1898. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, X.-X.; Liu, Y.-M.; Liu, D.-Y.; Chen, X.-P.; Zou, C.-Q. Zinc uptake by roots and accumulation in maize plants as affected by phosphorus application and arbuscular mycorrhizal colonization. Plant Soil 2017, 413, 59–71. [Google Scholar] [CrossRef]
- Loneragan, J.F. The effect of applied phosphate on the uptake of zinc by flax. Aust. J. Biol. Sci. 1951, 4, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bingham, F.T. Relation between phosphorus and micronutrients in plants. Soil Sci. Soc. Am. J. 1963, 27, 389–391. [Google Scholar] [CrossRef]
- Olsen, S.R. Micronutrient interactions. In Micronutrients in Agriculture; Dinauer, R.C., Ed.; Soil Science Society of America: Madison, WI, USA, 1972; pp. 243–264. [Google Scholar]
- Tagwira, F.; Piha, M.; Mugwira, L. Effect of pH, and phosphorus and organic matter contents on zinc availability and distribution in two Zimbabwean soils. Commun. Soil Sci. Plant Anal. 1992, 23, 1485–1500. [Google Scholar] [CrossRef]
- Tagwira, F.; Piha, M.; Mugwira, L. Zinc studies in Zimbabwean soils: Effect of lime and phosphorus on growth, yield, and zinc status of maize. Commun. Soil Sci. Plant Anal. 1993, 24, 717–736. [Google Scholar] [CrossRef]
- Hamilton, M.A.; Westermann, D.T.; James, D.W. Factors affecting zinc uptake in cropping systems. Soil Sci. Soc. Am. J. 1993, 57, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Burleson, C.A.; Dacus, A.D.; Gerard, C.J. Effect of Phosphorus Fertilization on the Zinc Nutrition of Several Irrigated Crops. Soil Sci. Soc. Am. J. 1961, 5, 365–368. [Google Scholar] [CrossRef]
- Langin, E.J.; Ward, R.C.; Olson, R.A.; Rhoades, H.F. Factors responsible for poor response of corn and grain sorghum to phosphorus fertilization: II. Lime and P placement effects on P-Zn relations. Soil Sci. Soc. Am. J. 1962, 26, 574–578. [Google Scholar] [CrossRef]
- Stukenholtz, D.D.; Olsen, R.J.; Gogan, G.; Olson, R.A. On the mechanism of phosphorus-zinc interaction in corn nutrition. Soil Sci. Soc. Am. J. 1966, 30, 759–763. [Google Scholar] [CrossRef]
- Sharma, K.C.; Krantz, B.A.; Brown, A.L.; Quick, J. Interaction of Zn and P in Top and Root of Corn and Tomato 1. Agron. J. 1968, 60, 453–456. [Google Scholar] [CrossRef]
- Warnock, R.E. Micronutrient uptake and mobility within corn plants (Zea mays L.) in relation to phosphorus-induced zinc deficiency. Soil Sci. Soc. Am. J. 1970, 34, 765–769. [Google Scholar] [CrossRef]
- Adriano, D.C.; Paulsen, G.M.; Murphy, L.S. Phosphorus-Iron and Phosphorus-Zinc Relationships in Corn (Zea mays L.) Seedlings as Affected by Mineral Nutrition 1. Agron. J. 1971, 63, 36–39. [Google Scholar] [CrossRef]
- Orabi, A.A.; Abdallah, A.; Mashadi, H.; Barakat, A.H. Zinc-phosphorus relationship in the nutrition of corn plants (Zea mays L.) grown on some calcareous soils. Plant Soil 1981, 59, 51–59. [Google Scholar] [CrossRef]
- Loneragan, J.F.; Grove, T.S.; Robson, A.D.; Snowball, K. Phosphorus toxicity as a factor in zinc-phosphorus interactions in plants. Soil Sci. Soc. Am. J. 1979, 43, 966–972. [Google Scholar] [CrossRef]
- Moraghan, J.T.; Mascagni, H.J., Jr. Environmental and soil factors affecting micronutrient deficiencies and toxicities. Micronutr. Agric. 1991, 4, 371–425. [Google Scholar]
- Shulka, U.C.; Morris, H.D. Relative efficiency of several zinc sources for corn. Agron. J 1967, 59, 200–202. [Google Scholar]
- Allison, J.D.; Brown, D.S.; Novo-Gradac, K.J. MINTEQA2/PRODEFA2, A Geochemical Assessment Model for Environmental Systems: Version 3.0 User’s Manual; Environmental Research Laboratory; Office of Research and Development; US Environmental Protection Agency: Washington, DC, USA, 1991.
- Campbell, C. Reference Sufficiency Ranges for Plant Analysis in the Southern Region of the United States; Southern Region Agricultural Experiment Station: Fayetteville, AR, USA, 2000. [Google Scholar]
- Karimian, N. Effect of nitrogen and phosphorus on zinc nutrition of corn in a calcareous soil. J. Plant Nutr. 1995, 18, 2261–2271. [Google Scholar] [CrossRef]
- Drissi, S.; Aït Houssa, A.; Bamouh, A.; Coquant, J.-M.; Benbella, M. Effect of zinc-phosphorus interaction on corn silage grown on sandy soil. Agriculture 2015, 5, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Bogdanovic, D.; Ubavic, M.; Cuvardic, M. Effect of phosphorus fertilization on Zn and Cd contents in soil and corn plants. Nutr. Cycl. Agroecosystems 1999, 54, 49–56. [Google Scholar] [CrossRef]
- Lönnerdal, B. Phytic acid–trace element (Zn, Cu, Mn) interactions. Int. J. Food Sci. Technol. 2002, 37, 749–758. [Google Scholar] [CrossRef]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef]
- Shukla, R.; Cheryan, M. Zein: The industrial protein from corn. Ind. Crops Prod. 2001, 13, 171–192. [Google Scholar] [CrossRef]
- Sharma, A.; Patni, B.; Shankhdhar, D.; Shankhdhar, S.C. Zinc–an indispensable micronutrient. Physiol. Mol. Biol. Plants 2013, 19, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Metal ligands in micronutrient acquisition and homeostasis. Plant. Cell Environ. 2019, 42, 2902–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talib, E.A.; Outten, C.E. Iron-sulfur cluster biogenesis, trafficking, and signaling: Roles for CGFS glutaredoxins and BolA proteins. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2021, 1868, 118847. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, S.; Verbruggen, N.; Cuypers, A.; Meyer, A.J. Essential trace metals in plant responses to heat stress. J. Exp. Bot. 2022, 73, 1775–1788. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Pfeiffer, W.H.; McClafferty, B. Biofortification of durum wheat with zinc and iron. Cereal Chem. 2010, 87, 10–20. [Google Scholar] [CrossRef]
- Peterson, C.J.; Johnson, V.A.; Mattern, P.J. Inluence of Cultivar and Environment on Mineral and Protein Concentrations of Wheat Flour, Bran, and Grain. Cereal Chem. 1986, 63, 183–186. [Google Scholar]
| P Treatment (mg L−1) | Leaf (mg) | Stem (mg) | Roots (mg) | Grain (mg) | Total Biomass (mg) | Root:Shoot | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | |
| 4 | 664 | 165 | 556 | 357 | 299 | 192 | 2182 | 560 | 3700 | 774 | 0.09 | 0.05 |
| 8 | 943 | 164 | 1366 | 584 | 801 | 240 | 3467 | 668 | 6576 | 696 | 0.14 | 0.04 |
| 12 | 960 | 142 | 1410 | 403 | 976 | 306 | 3732 | 541 | 7078 | 688 | 0.16 | 0.06 |
| 15 | 1262 | 456 | 2456 | 1536 | 1271 | 335 | 3336 | 1436 | 8325 | 982 | 0.18 | 0.05 |
| 20 | 1205 | 398 | 2495 | 1486 | 1166 | 471 | 3070 | 1274 | 7937 | 1402 | 0.17 | 0.05 |
| 22 | 1426 | 440 | 2970 | 1012 | 1368 | 517 | 2956 | 951 | 8721 | 1089 | 0.18 | 0.07 |
| K | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | |
| 4 | 970 | 312 | 985 | 147 | 237 | 229 | 360 | 66 | 2552 | 593 | 0.09 | 0.08 |
| 8 | 1345 | 514 | 1734 | 289 | 623 | 321 | 599 | 118 | 4301 | 893 | 0.17 | 0.08 |
| 12 | 1357 | 374 | 2015 | 597 | 966 | 341 | 670 | 64 | 5008 | 726 | 0.24 | 0.08 |
| 15 | 1545 | 470 | 2606 | 924 | 1399 | 463 | 651 | 279 | 6202 | 961 | 0.30 | 0.12 |
| 20 | 1657 | 694 | 2565 | 850 | 1398 | 651 | 642 | 249 | 6263 | 1511 | 0.29 | 0.12 |
| 22 | 1743 | 550 | 3069 | 614 | 1652 | 921 | 676 | 230 | 7140 | 1242 | 0.30 | 0.16 |
| P | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg * | Std | |
| 4 | 30 | 6 | 19 | 6 | 13 | 5 | 218 | 25 | 280 | 28 | 0.05 | 0.02 |
| 8 | 50 | 4 | 47 | 23 | 34 | 14 | 401 | 54 | 532 | 55 | 0.07 | 0.03 |
| 12 | 55 | 12 | 56 | 22 | 44 | 16 | 458 | 147 | 613 | 146 | 0.10 | 0.10 |
| 15 | 90 | 52 | 164 | 151 | 79 | 28 | 532 | 213 | 864 | 65 | 0.10 | 0.04 |
| 20 | 94 | 39 | 216 | 173 | 87 | 41 | 547 | 246 | 943 | 108 | 0.10 | 0.05 |
| 22 | 158 | 83 | 315 | 111 | 119 | 59 | 548 | 263 | 1140 | 182 | 0.13 | 0.08 |
| S | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg * | Std | Avg ** | Std | Avg ** | Std | |
| 4 | 171 | 52 | 52 | 19 | 80 | 50 | 146 | 29 | 448 | 117 | 0.21 | 0.12 |
| 8 | 366 | 101 | 155 | 45 | 237 | 89 | 248 | 50 | 1006 | 178 | 0.32 | 0.12 |
| 12 | 395 | 65 | 169 | 52 | 330 | 133 | 255 | 87 | 1150 | 166 | 0.41 | 0.18 |
| 15 | 363 | 94 | 254 | 107 | 460 | 142 | 254 | 121 | 1331 | 195 | 0.54 | 0.20 |
| 20 | 384 | 91 | 244 | 137 | 467 | 363 | 233 | 116 | 1327 | 447 | 0.52 | 0.34 |
| 22 | 417 | 79 | 290 | 99 | 483 | 283 | 218 | 108 | 1408 | 362 | 0.51 | 0.25 |
| Mg | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg * | Std | |
| 4 | 109 | 31 | 43 | 17 | 30 | 26 | 75 | 13 | 257 | 66 | 0.13 | 0.10 |
| 8 | 287 | 65 | 146 | 46 | 63 | 34 | 145 | 19 | 640 | 101 | 0.11 | 0.06 |
| 12 | 300 | 53 | 156 | 50 | 104 | 45 | 165 | 15 | 726 | 114 | 0.17 | 0.07 |
| 15 | 344 | 111 | 252 | 101 | 164 | 103 | 171 | 70 | 931 | 192 | 0.22 | 0.16 |
| 20 | 353 | 93 | 237 | 137 | 163 | 107 | 173 | 67 | 927 | 256 | 0.20 | 0.10 |
| 22 | 385 | 121 | 247 | 78 | 192 | 91 | 167 | 48 | 991 | 224 | 0.24 | 0.08 |
| Fe | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg | Std | |
| 4 | 2.1 | 0.6 | 1.0 | 0.8 | 11 | 10 | 1.8 | 0.5 | 16 | 11 | 2.1 | 1.23 |
| 8 | 3.8 | 1.2 | 2.8 | 1.3 | 31 | 13 | 3.3 | 0.8 | 41 | 14 | 3.1 | 0.97 |
| 12 | 4.0 | 0.6 | 3.5 | 1.4 | 30 | 15 | 3.6 | 1.0 | 41 | 16 | 2.6 | 1.20 |
| 15 | 5.4 | 1.8 | 5.7 | 3.2 | 43 | 11 | 3.2 | 1.4 | 57 | 40 | 3.2 | 3.09 |
| 20 | 5.5 | 1.4 | 6.1 | 3.3 | 42 | 22 | 2.9 | 1.6 | 57 | 23 | 2.9 | 1.28 |
| 22 | 5.8 | 1.4 | 7.2 | 2.2 | 37 | 18 | 2.9 | 1.1 | 53 | 19 | 2.3 | 0.98 |
| Zn | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | |
| 4 | 2.5 | 0.5 | 0.7 | 0.4 | 0.8 | 0.4 | 1.7 | 0.3 | 6 | 1.2 | 0.16 | 0.06 |
| 8 | 3.7 | 1.0 | 1.6 | 0.9 | 1.4 | 0.5 | 2.9 | 0.5 | 10 | 2.0 | 0.18 | 0.06 |
| 12 | 4.2 | 0.9 | 1.9 | 1.0 | 1.9 | 0.9 | 3.5 | 0.3 | 12 | 2.5 | 0.19 | 0.08 |
| 15 | 3.8 | 1.6 | 3.1 | 2.5 | 2.6 | 0.8 | 2.9 | 1.2 | 12 | 3.1 | 0.29 | 0.11 |
| 20 | 3.6 | 1.3 | 3.1 | 2.1 | 2.4 | 1.0 | 3.0 | 1.1 | 12 | 2.8 | 0.25 | 0.08 |
| 22 | 3.6 | 1.2 | 3.6 | 1.4 | 2.8 | 1.5 | 2.9 | 1.1 | 13 | 2.8 | 0.28 | 0.14 |
| B | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg | Std | Avg | Std | Avg ** | Std | Avg | Std | |
| 4 | 4.4 | 1.2 | 1.4 | 0.8 | 0.38 | 0.27 | 0.8 | 0.3 | 7 | 1.5 | 0.06 | 0.04 |
| 8 | 7.2 | 1.4 | 3.3 | 2.5 | 0.72 | 0.62 | 1.4 | 0.6 | 13 | 4.0 | 0.06 | 0.03 |
| 12 | 9.2 | 1.7 | 3.9 | 2.1 | 0.61 | 0.29 | 1.4 | 0.3 | 15 | 3.2 | 0.04 | 0.02 |
| 15 | 9.6 | 2.2 | 4.7 | 2.8 | 0.91 | 0.30 | 1.2 | 0.6 | 16 | 3.5 | 0.06 | 0.02 |
| 20 | 9.8 | 2.0 | 6.5 | 5.8 | 0.74 | 0.22 | 1.4 | 0.7 | 18 | 6.2 | 0.05 | 0.02 |
| 22 | 10.7 | 3.3 | 5.4 | 3.1 | 0.90 | 0.95 | 1.3 | 0.6 | 18 | 5.2 | 0.05 | 0.04 |
| Cu | ||||||||||||
| Avg | Std | Avg ** | Std | Avg ** | Std | Avg * | Std | Avg ** | Std | Avg | Std | |
| 4 | 1.6 | 0.64 | 0.2 | 0.07 | 0.8 | 0.36 | 0.19 | 0.09 | 2.7 | 0.7 | 0.44 | 0.25 |
| 8 | 2.0 | 0.63 | 0.8 | 0.73 | 1.5 | 0.73 | 0.48 | 0.48 | 4.8 | 1.5 | 0.50 | 0.24 |
| 12 | 2.6 | 0.80 | 0.8 | 0.36 | 1.9 | 1.02 | 0.65 | 0.63 | 5.8 | 2.2 | 0.49 | 0.26 |
| 15 | 2.4 | 1.13 | 1.1 | 0.48 | 2.2 | 0.84 | 0.24 | 0.15 | 6.0 | 1.4 | 0.62 | 0.24 |
| 20 | 2.3 | 0.89 | 1.1 | 0.51 | 2.0 | 1.04 | 0.38 | 0.43 | 5.8 | 1.3 | 0.56 | 0.31 |
| 22 | 2.1 | 0.90 | 1.5 | 0.51 | 2.3 | 1.38 | 0.23 | 0.13 | 6.1 | 1.9 | 0.60 | 0.37 |
| Mn | ||||||||||||
| Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg ** | Std | Avg | Std | |
| 4 | 1.1 | 0.31 | 0.24 | 0.12 | 0.21 | 0.11 | 0.31 | 0.09 | 1.9 | 0.5 | 0.12 | 0.05 |
| 8 | 2.8 | 0.77 | 0.69 | 0.45 | 0.55 | 0.26 | 0.67 | 0.19 | 4.7 | 1.2 | 0.13 | 0.05 |
| 12 | 3.3 | 0.77 | 0.70 | 0.21 | 0.61 | 0.24 | 0.82 | 0.14 | 5.4 | 1.0 | 0.13 | 0.05 |
| 15 | 3.7 | 1.07 | 1.14 | 0.57 | 0.84 | 0.53 | 0.86 | 0.43 | 6.5 | 1.5 | 0.15 | 0.08 |
| 20 | 3.8 | 0.87 | 1.20 | 0.66 | 0.75 | 0.30 | 0.88 | 0.36 | 6.7 | 1.3 | 0.13 | 0.04 |
| 22 | 4.5 | 1.02 | 1.46 | 0.55 | 0.77 | 0.44 | 0.84 | 0.33 | 7.6 | 1.3 | 0.11 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penn, C.J.; Camberato, J.J.; Wiethorn, M.A. How Much Phosphorus Uptake Is Required for Achieving Maximum Maize Grain Yield? Part 2: Impact of Phosphorus Uptake on Grain Quality and Partitioning of Nutrients. Agronomy 2023, 13, 258. https://doi.org/10.3390/agronomy13010258
Penn CJ, Camberato JJ, Wiethorn MA. How Much Phosphorus Uptake Is Required for Achieving Maximum Maize Grain Yield? Part 2: Impact of Phosphorus Uptake on Grain Quality and Partitioning of Nutrients. Agronomy. 2023; 13(1):258. https://doi.org/10.3390/agronomy13010258
Chicago/Turabian StylePenn, Chad J., James J. Camberato, and Matthew A. Wiethorn. 2023. "How Much Phosphorus Uptake Is Required for Achieving Maximum Maize Grain Yield? Part 2: Impact of Phosphorus Uptake on Grain Quality and Partitioning of Nutrients" Agronomy 13, no. 1: 258. https://doi.org/10.3390/agronomy13010258
APA StylePenn, C. J., Camberato, J. J., & Wiethorn, M. A. (2023). How Much Phosphorus Uptake Is Required for Achieving Maximum Maize Grain Yield? Part 2: Impact of Phosphorus Uptake on Grain Quality and Partitioning of Nutrients. Agronomy, 13(1), 258. https://doi.org/10.3390/agronomy13010258